
Genome-Wide Association Study of Maternal Genetic Effects on Intramuscular Fat and Fatty Acid Composition in Rabbits

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals and Phenotypes
2.3. Quantifying the Importance of Maternal Genetic Effects
2.4. Genotype Data
2.5. Maternal Genome-Wide Association and Gene Annotation
3. Results and Discussion
3.1. Quantifying the Importance of Maternal Genetic Effects
3.1.1. Descriptive Statistics
3.1.2. Estimate of Genetic, Maternal, and Common Environment Parameters for IMF
3.1.3. Estimates of Genetic, Maternal, and Common Environment Parameters for Fatty Acid Composition
3.1.4. Estimates of Direct Additive and Maternal Genetic Correlations between IMF and Fatty Acid Composition, and the Direct-Maternal Genetic Correlations of Fatty Acid Composition
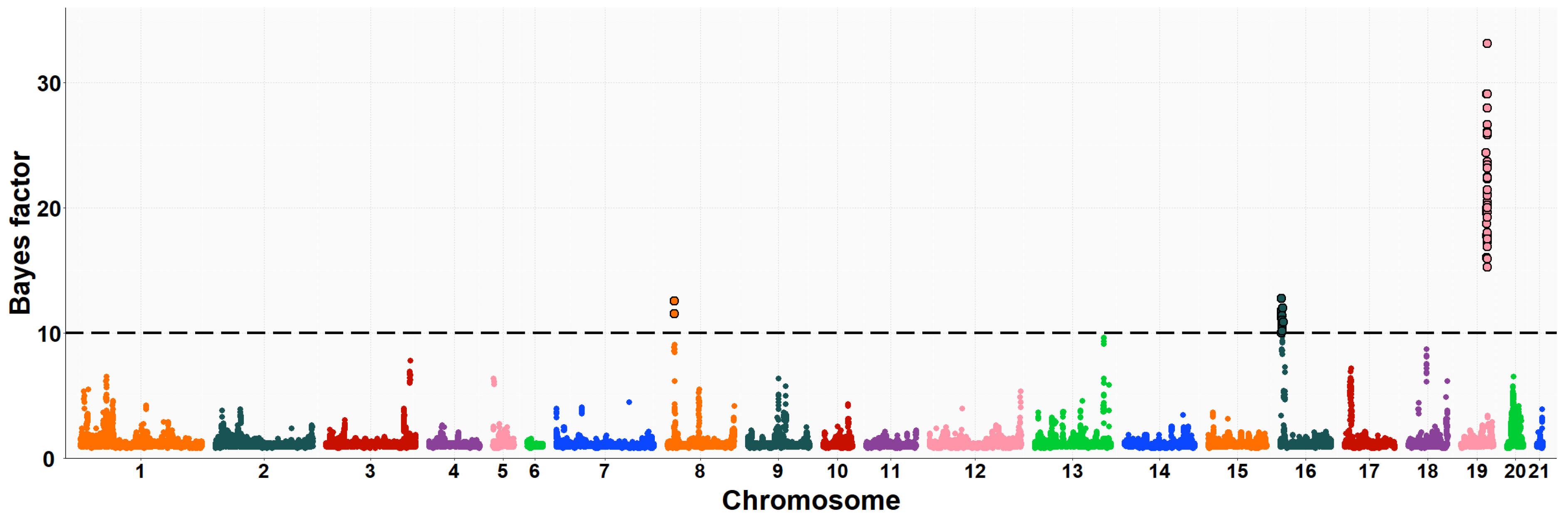
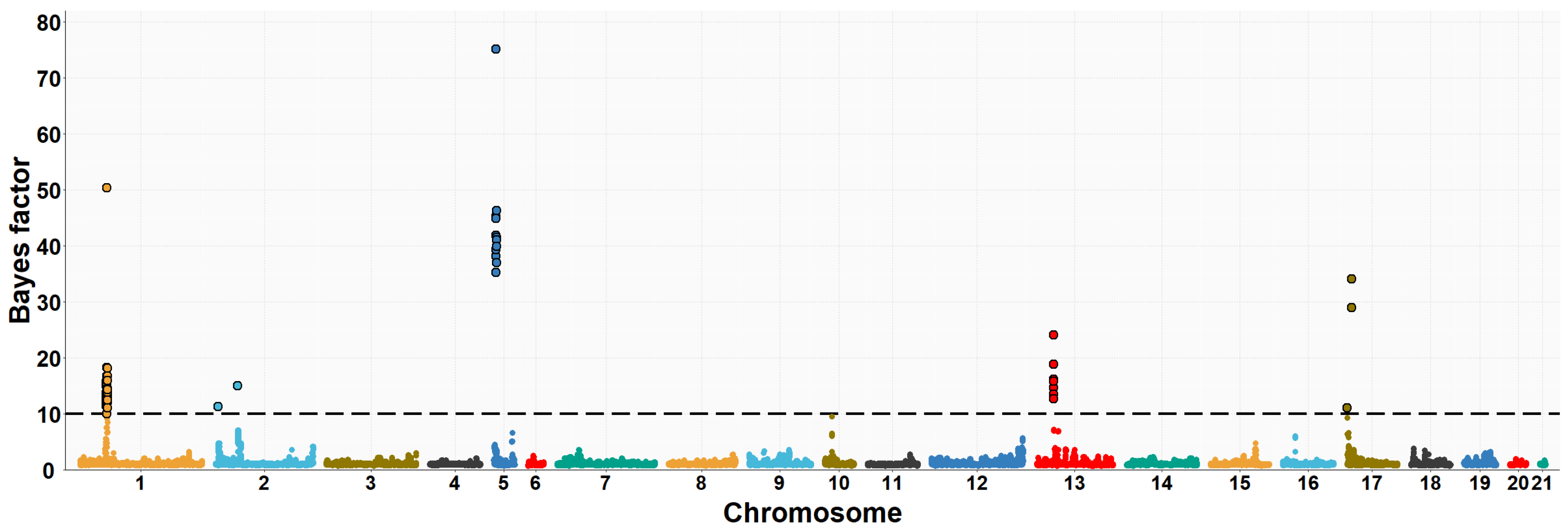
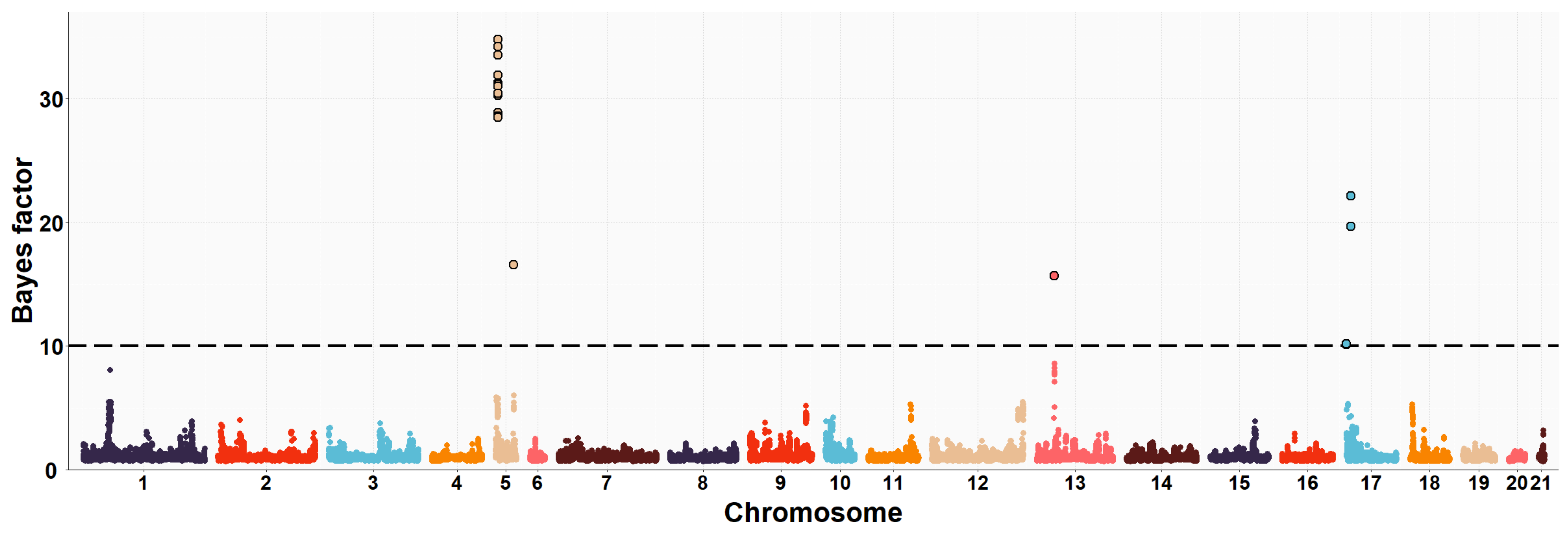
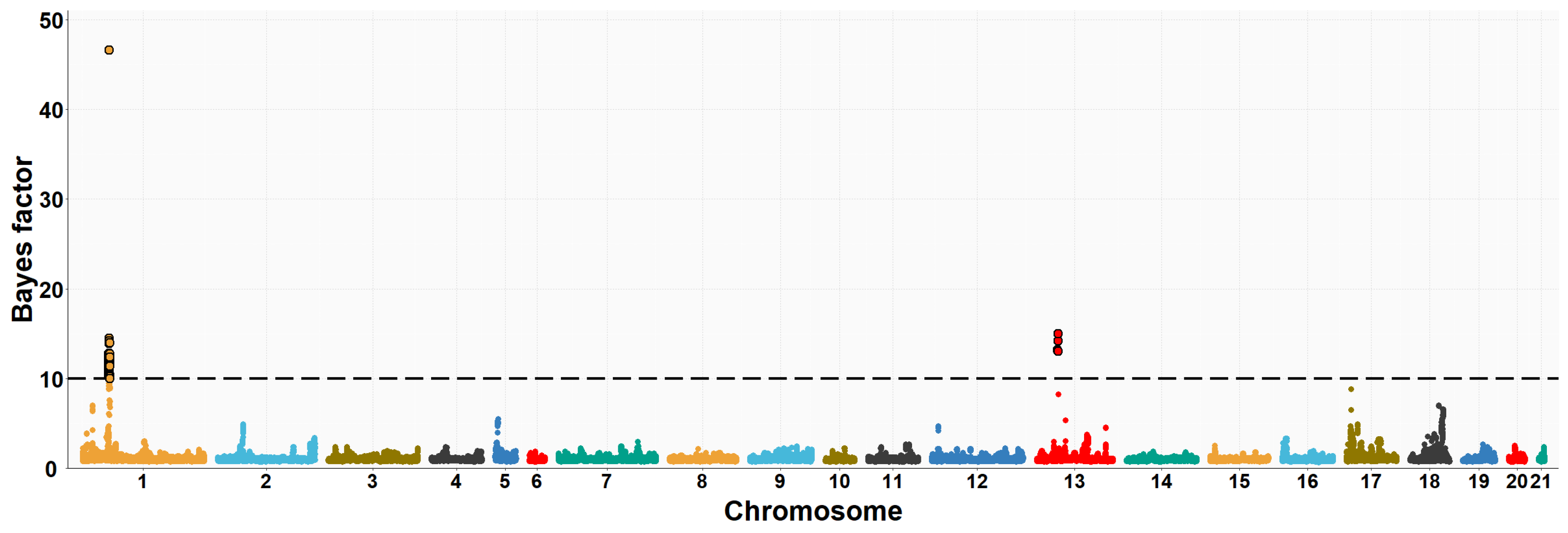
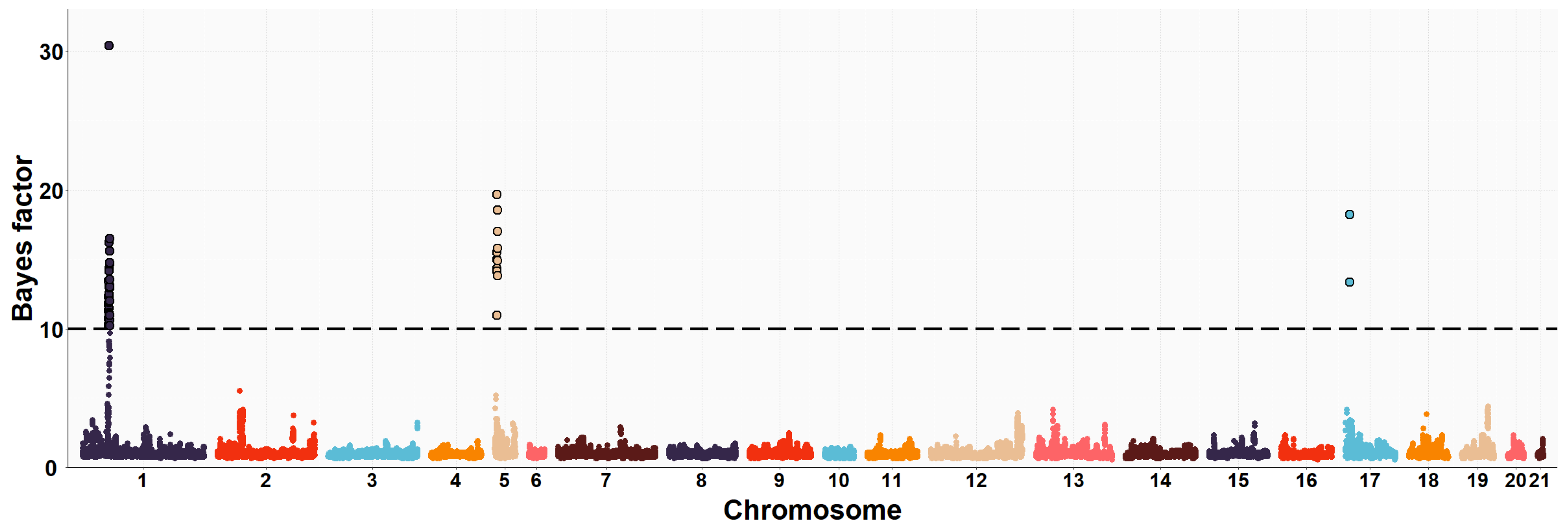
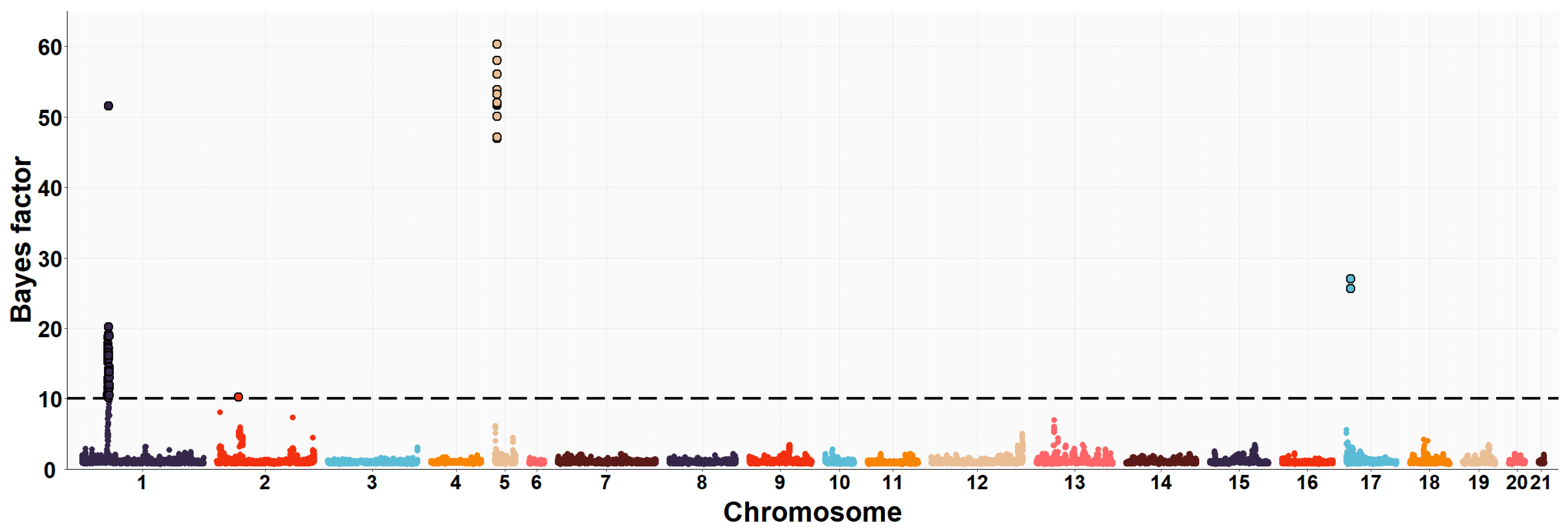
3.2. Maternal GWAS for IMF and Fatty Acid Composition
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bijma, P. Estimating Maternal Genetic Effects in Livestock. J. Anim. Sci. 2006, 84, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.B.; Wade, M.J. What Are Maternal Effects (and What Are They Not)? Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Maniatis, N.; Pollott, G.E. Maternal Effects on Weight and Ultrasonically Measured Traits of Lambs in a Small Closed Suffolk Flock. Small Rumin. Res. 2002, 45, 235–246. [Google Scholar] [CrossRef]
- Meyer, K.; Carrick, M.J.; Donnelly, B.J.P. Genetic Parameters for Growth Traits of Australian Beef Cattle from a Multibreed Selection Experiment. J. Anim. Sci. 1993, 71, 2614–2622. [Google Scholar] [CrossRef]
- Ibáñez-Escriche, N.; Magallón, E.; Gonzalez, E.; Tejeda, J.F.; Noguera, J.L. Genetic Parameters and Crossbreeding Effects of Fat Deposition and Fatty Acid Profiles in Iberian Pig Lines. J. Anim. Sci. 2016, 94, 28–37. [Google Scholar] [CrossRef]
- Gol, S.; González-Prendes, R.; Bosch, L.; Tor, M.; Reixach, J.; Pena, R.N.; Estany, J. Linoleic Acid Metabolic Pathway Allows for an Efficient Increase of Intramuscular Fat Content in Pigs. J. Anim. Sci. Biotechnol. 2019, 10, 33. [Google Scholar] [CrossRef]
- Wood, J.D.; Nute, G.R.; Richardson, R.I.; Whittington, F.M.; Southwood, O.; Plastow, G.; Mansbridge, R.; Da Costa, N.; Chang, K.C. Effects of Breed, Diet and Muscle on Fat Deposition and Eating Quality in Pigs. Meat Sci. 2004, 67, 651–667. [Google Scholar] [CrossRef]
- Hocquette, J.F.; Gondret, F.; Baza, E.; Mdale, F.; Jurie, C.; Pethick, D.W. Intramuscular Fat Content in Meat-Producing Animals: Development, Genetic and Nutritional Control, and Identification of Putative Markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef]
- Aboagye, G.; Zappaterra, M.; Laghi, L.; Dall’Olio, S.; Petracci, M.; Nanni Costa, L. Water Status in Meat from Pig Breeds Strongly Differing in Growth Performances. Food Chem. 2020, 305, 125445. [Google Scholar] [CrossRef]
- Martinez-Alvaro, M.; Blasco, A.; Hernandez, P. Effect of Selection for Intramuscular Fat on the Fatty Acid Composition of Rabbit Meat. Animal 2018, 12, 2002–2008. [Google Scholar] [CrossRef]
- Sosa-Madrid, B.S.; Hernández, P.; Blasco, A.; Haley, C.S.; Fontanesi, L.; Santacreu, M.A.; Pena, R.N.; Navarro, P.; Ibáñez-Escriche, N. Genomic Regions Influencing Intramuscular Fat in Divergently Selected Rabbit Lines. Anim. Genet. 2020, 51, 58–69. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, R.; Hu, X.; Li, N. Application of Genomic Technologies to the Improvement of Meat Quality of Farm Animals. Meat Sci. 2007, 77, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Ros-Freixedes, R.; Gol, S.; Pena, R.N.; Tor, M.; Ibáñez-Escriche, N.; Dekkers, J.C.M.; Estany, J. Genome-Wide Association Study Singles out SCD and LEPR as the Two Main Loci Influencing Intramuscular Fat Content and Fatty Acid Composition in Duroc Pigs. PLoS ONE 2016, 11, e0152496. [Google Scholar] [CrossRef] [PubMed]
- Pena, R.N.; Noguera, J.L.; García-Santana, M.J.; González, E.; Tejeda, J.F.; Ros-Freixedes, R.; Ibáñez-Escriche, N. Five Genomic Regions Have a Major Impact on Fat Composition in Iberian Pigs. Sci. Rep. 2019, 9, 2031. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Álvaro, M.; Hernández, P.; Blasco, A. Divergent Selection on Intramuscular Fat in Rabbits: Responses to Selection and Genetic Parameters. J. Anim. Sci. 2016, 94, 4993–5003. [Google Scholar] [CrossRef]
- Zubiri-Gaitán, A.; Blasco, A.; Ccalta, R.; Satué, K.; Hernández, P. Intramuscular Fat Selection in Rabbits Modifies the Fatty Acid Composition of Muscle and Liver Tissues. Animals 2022, 12, 893. [Google Scholar] [CrossRef] [PubMed]
- Zomeño, C.; Juste, V.; Hernández, P. Application of NIRS for Predicting Fatty Acids in Intramuscular Fat of Rabbit. Meat Sci. 2012, 91, 155–159. [Google Scholar] [CrossRef]
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A Direct Method for Fatty Acid Methyl Ester Synthesis: Application to Wet Meat Tissues, Oils, and Feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef]
- Laghouaouta, H.; Sosa-Madrid, B.S.; Zubiri-Gaitán, A.; Hernández, P.; Blasco, A. Novel Genomic Regions Associated with Intramuscular Fatty Acid Composition in Rabbits. Animals 2020, 10, 2090. [Google Scholar] [CrossRef]
- Blasco, A. Bayesian Data Analysis for Animal Scientists; Springer: Berlin/Heidelberg, Germany, 2017. [Google Scholar]
- Misztal, I.; Tsuruta, S.; Strabel, T.; Auvray, B.; Druet, T.; Lee, D.H. Blupf90 and Related Programs (Bgf90). In Proceedings of the 7th World Congress on Genetics Applied to Livestock Production, Montpellier, France, 19–23 August 2002; pp. 743–744. [Google Scholar]
- Green, M.R.; Sambrook, J. Quantification of DNA and RNA by Real-Time Polymerase Chain Reaction. In Molecular Cloning: Laboratory Manual, 4th ed.; Cold Spring Harb. Lab. Press: New York, NY, USA, 2012; pp. 631–679. [Google Scholar]
- Browning, B.L.; Browning, S.R. Genotype Imputation with Millions of Reference Samples. Am. J. Hum. Genet. 2016, 98, 116–126. [Google Scholar] [CrossRef]
- Marras, G.; Rossoni, A.; Schwarzenbacher, H.; Biffani, S.; Biscarini, F.; Nicolazzi, E.L. Zanardi: An Open-Source Pipeline for Multiple-Species Genomic Analysis of SNP Array Data. Anim. Genet. 2017, 48, 121. [Google Scholar] [CrossRef] [PubMed]
- Garrick, D.J.; Fernando, R.L. Implementing a QTL Detection Study (GWAS) Using Genomic Prediction Methodology. In Genome-Wide Association Studies and Genomic Prediction; Springer Nature: Berlin/Heidelberg, Germany, 2013; pp. 275–298. [Google Scholar]
- Kass, R.E.; Raftery, A.E. Bayes Factors. J. Am. Stat. Assoc. 1995, 90, 773–795. [Google Scholar] [CrossRef]
- Cunningham, F.; Achuthan, P.; Akanni, W.; Allen, J.; Amode, M.R.; Armean, I.M.; Bennett, R.; Bhai, J.; Billis, K.; Boddu, S.; et al. Ensembl 2019. Nucleic Acids Res. 2019, 47, D745–D751. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics Enrichment Tools: Paths toward the Comprehensive Functional Analysis of Large Gene Lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jiao, X.; Sherman, B.T.; Huang, D.W.; Stephens, R.; Baseler, M.W.; Lane, H.C.; Lempicki, R.A. DAVID-WS: A Stateful Web Service to Facilitate Gene/Protein List Analysis. Bioinformatics 2012, 28, 1805–1806. [Google Scholar] [CrossRef]
- Hernández, P.; Zotte, A.D. Influence of Diet on Rabbit Meat Quality. In Nutrition of the Rabbit; CABI: Wallingford, UK, 2010; pp. 163–178. [Google Scholar]
- Dalle Zotte, A. Perception of Rabbit Meat Quality and Major Factors Influencing the Rabbit Carcass and Meat Quality. Livest. Prod. Sci. 2002, 75, 11–32. [Google Scholar] [CrossRef]
- Hoque, M.A.; Kadowaki, H.; Shibata, T.; Suzuki, K. Maternal and Direct Genetic Parameters for Production Traits and Maternal Correlations among Production and Feed Efficiency Traits in Duroc Pigs. Asian-Australas. J. Anim. Sci. 2008, 21, 961–966. [Google Scholar] [CrossRef]
- Zomeño, C.; Hernández, P.; Blasco, A. Divergent Selection for Intramuscular Fat Content in Rabbits. I. Direct Response to Selection. J. Anim. Sci. 2013, 91, 4526–4531. [Google Scholar] [CrossRef]
- Knapp, P.; Willam, A.; Sölkner, J. Genetic Parameters for Lean Meat Content and Meat Quality Traits in Different Pig Breeds. Livest. Prod. Sci. 1997, 52, 69–73. [Google Scholar] [CrossRef]
- Mcallister, C.M.; Speidel, S.E.; Crews, D.H., Jr.; Enns, R.M. Genetic Parameters for Intramuscular Fat Percentage, Marbling Score, Scrotal Circumference, and Heifer Pregnancy in Red Angus Cattle 1. J. Anim. Sci. 2011, 89, 2068–2072. [Google Scholar] [CrossRef]
- Torres-vazquez, J.A. Genetic Parameters for Docility, Weaning Weight, Yearling Weight and Intramuscular Fat Percentage in Hereford Cattle. J. Anim. Sci. 2016, 94, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Halli, K.; Bohlouli, M.; Schulz, L.; Sundrum, A.; König, S. Estimation of Direct and Maternal Genetic Effects and Annotation of Potential Candidate Genes for Weight and Meat Quality Traits in a Genotyped Outdoor Dual-Purpose Cattle Breed. Transl. Anim. Sci. 2022, 6, txac022. [Google Scholar] [CrossRef]
- Suzuki, K.; Irie, M.; Kadowaki, H.; Shibata, T.; Kumagai, M.; Nishida, A. Genetic Parameter Estimates of Meat Quality Traits in Duroc Pigs Selected for Average Daily Gain, Longissimus Muscle Area, Backfat Thickness, and Intramuscular Fat Content. J. Anim. Sci. 2005, 83, 2058–2065. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.; Carrick, M.J.; Donnelly, B.J.P. Genetic Parameters for Milk Production of Australian Beef Cows and Weaning Weight of Their Calves. J. Anim. Sci. 1994, 72, 1155–1165. [Google Scholar] [CrossRef] [PubMed]
- Sellier, P.; Maignel, L.; Bidanel, J.P. Genetic Parameters for Tissue and Fatty Acid Composition of Backfat, Perirenal Fat and Longissimus Muscle in Large White and Landrace Pigs. Animal 2010, 4, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Ros-Freixedes, R.; Reixach, J.; Bosch, L.; Tor, M.; Estany, J. Genetic Correlations of Intramuscular Fat Content and Fatty Acid Composition among Muscles and with Subcutaneous Fat in Duroc Pigs. J. Anim. Sci. 2014, 92, 5417–5425. [Google Scholar] [CrossRef]
- Nogi, T.; Honda, T.; Mukai, F.; Okagaki, T.; Oyama, K. Heritabilities and Genetic Correlations of Fatty Acid Compositions in Longissimus Muscle Lipid with Carcass Traits in Japanese Black Cattle. J. Anim. Sci. 2011, 89, 615–621. [Google Scholar] [CrossRef]
- Buchanan, J.W.; Reecy, J.M.; Garrick, D.J.; Duan, Q.; Beitz, D.C.; Mateescu, R.G. Genetic Parameters and Genetic Correlations among Triacylglycerol and Phospholipid Fractions in Angus Cattle. J. Anim. Sci. 2015, 93, 522–528. [Google Scholar] [CrossRef]
- Quintanilla, R.; Piedrafita, J. Efectos Maternos En El Peso Al Destete Del Ganado Vacuno de Carne: Revisión. Itea 2000, 96, 7–39. [Google Scholar]
- Lee, C. On the Negative Estimates of Direct and Maternal Genetic Correlation—A Review. Asian-Australas. J. Anim. Sci. 2002, 15, 1222–1226. [Google Scholar] [CrossRef]
- Olsen, H.G.; Hayes, B.J.; Kent, M.P.; Nome, T.; Svendsen, M.; Lien, S. A Genome Wide Association Study for QTL Affecting Direct and Maternal Effects of Stillbirth and Dystocia in Cattle. Anim. Genet. 2010, 41, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Yin, T.; König, S. Genome-Wide Associations and Detection of Potential Candidate Genes for Direct Genetic and Maternal Genetic Effects Influencing Dairy Cattle Body Weight at Different Ages. Genet. Sel. Evol. 2019, 51, 4. [Google Scholar] [CrossRef] [PubMed]
- Huttner, W.B.; Schmidt, A. Lipids, Lipid Modification and Lipid-Protein Interaction in Membrane Budding and Fission—Insights from the Roles of Endophilin A1 and Synaptophysin in Synaptic Vesicle Endocytosis. Curr. Opin. Neurobiol. 2000, 10, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Kogelman, L.J.A.; Pant, S.D.; Fredholm, M.; Kadarmideen, H.N. Systems Genetics of Obesity in an F2 Pig Model by Genome-Wide Association, Genetic Network, and Pathway Analyses. Front. Genet. 2014, 5, 214. [Google Scholar] [CrossRef]
- Hess, M.E.; Brüning, J.C. The Fat Mass and Obesity-Associated (FTO) Gene: Obesity and Beyond? Biochim. Biophys. Acta Mol. Basis Dis. 2014, 1842, 2039–2047. [Google Scholar] [CrossRef]
- Fan, B.; Du, Z.-Q.; Rothschild, M.F. The Fat Mass and Obesity-Associated (FTO) Gene Is Associated with Intramuscular Fat Content and Growth Rate in the Pig. Anim. Biotechnol. 2009, 20, 58–70. [Google Scholar] [CrossRef]
- Zhang, G.W.; Gao, L.; Chen, S.Y.; Zhao, X.B.; Tian, Y.F.; Wang, X.; Deng, X.S.; Lai, S.J. Single Nucleotide Polymorphisms in the FTO Gene and Their Association with Growth and Meat Quality Traits in Rabbits. Gene 2013, 527, 553–557. [Google Scholar] [CrossRef]
- Reyer, H.; Varley, P.F.; Murani, E.; Ponsuksili, S.; Wimmers, K. Genetics of Body Fat Mass and Related Traits in a Pig Population Selected for Leanness. Sci. Rep. 2017, 7, 9118. [Google Scholar] [CrossRef]
- Jin, C.; Fang, Z.; Lin, Y.; Che, L.; Wu, C.; Xu, S.; Feng, B.; Li, J.; Wu, D. Influence of Dietary Fat Source on Sow and Litter Performance, Colostrum and Milk Fatty Acid Profile in Late Gestation and Lactation. Anim. Sci. J. 2017, 88, 1768–1778. [Google Scholar] [CrossRef]
- Velasco, S.; Cañeque, V.; Lauzurica, S.; Pérez, C.; Huidobro, F. Effect of Different Feeds on Meat Quality and Fatty Acid Composition of Lambs Fattened at Pasture. Meat Sci. 2004, 66, 457–465. [Google Scholar] [CrossRef]
- Moreno, T.; Varela, A.; Oliete, B.; Carballo, J.A.; Sánchez, L.; Montserrat, L. Nutritional Characteristics of Veal from Weaned and Unweaned Calves: Discriminatory Ability of the Fat Profile. Meat Sci. 2006, 73, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Alfaia, C.M.M.; Castro, M.L.F.; Martins, S.I.V.; Portugal, A.P.V.; Alves, S.P.A.; Fontes, C.M.G.A.; Bessa, R.J.B.; Prates, J.A.M. Effect of Slaughter Season on Fatty Acid Composition, Conjugated Linoleic Acid Isomers and Nutritional Value of Intramuscular Fat in Barrosã-PDO Veal. Meat Sci. 2007, 75, 44–52. [Google Scholar] [CrossRef]
- Fontanesi, L.; Scotti, E.; Buttazzoni, L.; Davoli, R.; Russo, V. The Porcine Fat Mass and Obesity Associated (FTO) Gene Is Associated with Fat Deposition in Italian Duroc Pigs. Anim. Genet. 2009, 40, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Scotti, E.; Buttazzoni, L.; Dall’Olio, S.; Davoli, R.; Russo, V. A Single Nucleotide Polymorphism in the Porcine Cathepsin K (CTSK) Gene Is Associated with Back Fat Thickness and Production Traits in Italian Duroc Pigs. Mol. Biol. Rep. 2010, 37, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Fontanesi, L.; Schiavo, G.; Galimberti, G.; Calò, D.G.; Russo, V. A Genomewide Association Study for Average Daily Gain in Italian Large White Pigs. J. Anim. Sci. 2014, 92, 1385–1394. [Google Scholar] [CrossRef]
- Rempel, L.A.; Casas, E.; Shackelford, S.D.; Wheeler, T.L. Relationship of Polymorphisms within Metabolic Genes and Carcass Traits in Crossbred Beef Cattle. J. Anim. Sci. 2012, 90, 1311–1316. [Google Scholar] [CrossRef] [PubMed]
- Chung, E.-R. Novel SNP in the Coding Region of the FTO Gene Is Associated with Marbling Score in Hanwoo (Korean Cattle). J. Anim. Sci. Technol. 2014, 56, 27. [Google Scholar] [CrossRef] [PubMed]
- Jevsinek Skok, D.; Kunej, T.; Kovac, M.; Malovrh, S.; Potocnik, K.; Petric, N.; Zgur, S.; Dovc, P.; Horvat, S. FTO Gene Variants Are Associated with Growth and Carcass Traits in Cattle. Anim. Genet. 2016, 47, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Zielke, L.G.; Bortfeldt, R.H.; Reissmann, M.; Tetens, J.; Thaller, G.; Brockmann, G.A. Impact of Variation at the FTO Locus on Milk Fat Yield in Holstein Dairy Cattle. PLoS ONE 2013, 8, e63406. [Google Scholar] [CrossRef]
- Babenko, V.; Babenko, R.; Gamieldien, J.; Markel, A. FTO Haplotyping Underlines High Obesity Risk for European Populations. BMC Med. Genom. 2019, 12, 46. [Google Scholar] [CrossRef]
- Tews, D.; Fischer-Posovszky, P.; Wabitsch, M. Regulation of FTO and FTM Expression during Human Preadipocyte Differentiation. Horm. Metab. Res. 2011, 43, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Vogan, K. RPGRIP1L, FTO and Obesity. Nat. Genet. 2014, 46, 532. [Google Scholar] [CrossRef]
- Zheng, X.; Kuang, Y.; Lv, W.; Cao, D.; Sun, Z.; Sun, X. Genome-Wide Association Study for Muscle Fat Content and Abdominal Fat Traits in Common Carp (Cyprinus Carpio). PLoS ONE 2016, 11, e0169127. [Google Scholar] [CrossRef] [PubMed]
- Ishii, A.; Yamaji, K.; Uemoto, Y.; Sasago, N.; Kobayashi, E.; Kobayashi, N.; Matsuhashi, T.; Maruyama, S.; Matsumoto, H.; Sasazaki, S.; et al. Genome-Wide Association Study for Fatty Acid Composition in Japanese Black Cattle. Anim. Sci. J. 2013, 84, 675–682. [Google Scholar] [CrossRef]
- Liu, J.; Damon, M.; Guitton, N.; Guisle, I.; Ecolan, P.; Vincent, A.; Cherel, P.; Gondret, F. Differentially-Expressed Genes in Pig Longissimus Muscles with Contrasting Levels of Fat, as Identified by Combined Transcriptomic, Reverse Transcription PCR, and Proteomic Analyses. J. Agric. Food Chem. 2009, 57, 3808–3817. [Google Scholar] [CrossRef]
- Leung, P.S. The Peptide Hormone Angiotensin II: Its New Functions in Tissues and Organs. Curr. Protein Pept. Sci. 2004, 5, 267–273. [Google Scholar] [CrossRef]
- Riera-Fortuny, C.; Real, J.T.; Chaves, F.J.; Morales-Suárez-Varela, M.; Martínez-Triguero, M.L.; Morillas-Arino, C.; Hernández-Mijares, A. The Relation between Obesity, Abdominal Fat Deposit and the Angiotensin-Converting Enzyme Gene I/D Polymorphism and Its Association with Coronary Heart Disease. Int. J. Obes. 2005, 29, 78–84. [Google Scholar] [CrossRef]
- Lagou, V.; Manios, Y.; Moran, C.N.; Bailey, M.E.S.; Grammatikaki, E.; Oikonomou, E.; Ioannou, E.; Moschonis, G.; Wilson, R.H.; Pitsiladis, Y.P. Developmental Changes in Adiposity in Toddlers and Preschoolers in the GENESIS Study and Associations with the ACE I/D Polymorphism. Int. J. Obes. 2007, 31, 1052–1060. [Google Scholar] [CrossRef]
- Bienertova-Vasku, J.; Bienert, P.; Sablikova, L.; Slovackova, L.; Forejt, M.; Piskackova, Z.; Kucerova, L.; Heczkova, K.; Brazdova, Z.; Vasku, A. Effect of ID ACE Gene Polymorphism on Dietary Composition and Obesity-Related Anthropometric Parameters in the Czech Adult Population. Genes Nutr. 2009, 4, 207–213. [Google Scholar] [CrossRef]
- Hamill, R.M.; Aslan, O.; Mullen, A.M.; O’Doherty, J.V.; McBryan, J.; Morris, D.G.; Sweeney, T. Transcriptome Analysis of Porcine, M. Semimembranosus Divergent in Intramuscular Fat as a Consequence of Dietary Protein Restriction. BMC Genom. 2013, 14, 453. [Google Scholar] [CrossRef]
- Steinberg, G.R.; Dyck, D.J.; Calles-Escandon, J.; Tandon, N.N.; Luiken, J.J.F.P.; Glatz, J.F.C.; Bonen, A. Chronic Leptin Administration Decreases Fatty Acid Uptake and Fatty Acid Transporters in Rat Skeletal Muscle. J. Biol. Chem. 2002, 277, 8854–8860. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, T.G. Porcine Leptin Alters Isolated Adipocyte Glucose and Fatty Acid Metabolism. Domest. Anim. Endocrinol. 2004, 26, 11–21. [Google Scholar] [CrossRef]
- Steiner, A.A.; Romanovsky, A.A. Leptin: At the Crossroads of Energy Balance and Systemic Inflammation. Prog. Lipid Res. 2007, 46, 89–107. [Google Scholar] [CrossRef] [PubMed]
- Yau, M.H.; Wang, Y.; Lam, K.S.L.; Zhang, J.; Wu, D.; Xu, A. A Highly Conserved Motif within the NH2-Terminal Coiled-Coil Domain of Angiopoietin-like Protein 4 Confers Its Inhibitory Effects on Lipoprotein Lipase by Disrupting the Enzyme Dimerization. J. Biol. Chem. 2009, 284, 11942–11952. [Google Scholar] [CrossRef]
- Cánovas, A.; Quintanilla, R.; Amills, M.; Pena, R.N. Muscle Transcriptomic Profiles in Pigs with Divergent Phenotypes for Fatness Traits. BMC Genom. 2010, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Uemoto, Y.; Abe, T.; Tameoka, N.; Hasebe, H.; Inoue, K.; Nakajima, H.; Shoji, N.; Kobayashi, M.; Kobayashi, E. Whole-Genome Association Study for Fatty Acid Composition of Oleic Acid in Japanese Black Cattle. Anim. Genet. 2011, 42, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Lev, S. The Role of the Nir/RdgB Protein Family in Membrane Trafficking and Cytoskeleton Remodeling. Exp. Cell Res. 2004, 297, 1–10. [Google Scholar] [CrossRef]
- Muñoz, M.; Alves, E.; Corominas, J.; Folch, J.M.; Casellas, J.; Noguera, J.L.; Silió, L.; Fernández, A.I. Survey of SSC12 Regions Affecting Fatty Acid Composition of Intramuscular Fat Using High-Density SNP Data. Front. Genet. 2012, 2, 101. [Google Scholar] [CrossRef]
- Lim, D.; Kim, N.K.; Park, H.S.; Lee, S.H.; Cho, Y.M.; Oh, S.J.; Kim, T.H.; Kim, H. Identification of Candidate Genes Related to Bovine Marbling Using Pro-Tein-Protein Interaction Networks. Int. J. Biol. Sci. 2011, 7, 992–1002. [Google Scholar] [CrossRef]
- Wang, H.; Zhong, J.; Zhang, C.; Chai, Z.; Cao, H.; Wang, J.; Zhu, J.; Wang, J.; Ji, Q. The Whole-Transcriptome Landscape of Muscle and Adipose Tissues Reveals the CeRNA Regulation Network Related to Intramuscular Fat Deposition in Yak. BMC Genom. 2020, 21, 347. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Mean | Median | SD | CV (%) |
|---|---|---|---|---|
| IMF | 1.08 | 1.06 | 0.23 | 21.30 |
| C14:0 | 1.25 | 1.31 | 0.52 | 41.60 |
| C16:0 | 26.70 | 26.98 | 1.57 | 5.88 |
| C18:0 | 9.05 | 9.02 | 1.26 | 13.92 |
| SFA | 37.07 | 37.01 | 1.67 | 4.50 |
| C16:1n7 | 1.40 | 1.35 | 0.78 | 55.71 |
| C18:1n9 | 20.45 | 20.92 | 3.26 | 15.94 |
| MUFA | 23.77 | 24.52 | 4.55 | 19.14 |
| C18:2n6 | 26.95 | 27.06 | 2.97 | 11.02 |
| C18:3n3 | 1.64 | 1.69 | 0.43 | 26.22 |
| C20:4n6 | 6.57 | 6.12 | 1.99 | 30.29 |
| PUFA | 39.29 | 38.65 | 4.86 | 12.37 |
| MUFA/SFA | 0.64 | 0.66 | 0.03 | 4.69 |
| PUFA/SFA | 1.06 | 1.04 | 0.06 | 5.66 |
| Trait | dm | C2 | Me2 | ||
|---|---|---|---|---|---|
| Model 1 | 0.84 [0.69, 0.99] | ||||
| Model 2 | 0.44 [0.24, 0.64] | 0.22 [0.08, 0.36] | −0.02 [−0.54, 0.51] | ||
| Model 3 | 0.54 [0.39, 0.69] | 0.18 [0.12, 0.24] | |||
| Model 4 | 0.45 [0.28, 0.65] | 0.09 [0.01, 0.19] | −0.02 [−0.63, 0.59] | 0.14 [0.07, 0.21] | |
| Model 5 | 0.46 [0.28, 0.63] | 0.08 [0.01, 0.19] | 0 [−0.64, 0.64] | 0.13 [0.06, 0.21] |
| Trait | C2 | ||
|---|---|---|---|
| C14:0 | 0.50 [0.34, 0.67] | 0.16 [0.07, 0.26] | 0.16 [0.09, 0.23] |
| C16:0 | 0.54 [0.38, 0.71] | 0.15 [0.06, 0.24] | 0.14 [0.09, 0.20] |
| C18:0 | 0.40 [0.24, 0.56] | 0.10 [0.01, 0.19] | 0.14 [0.07, 0.20] |
| SFA | 0.35 [0.22, 0.47] | 0.14 [0.04, 0.24] | 0.21 [0.15, 0.27] |
| C16:1n7 | 0.45 [0.27, 0.62] | 0.46 [0.38, 0.54] | 0.15 [0.08, 0.22] |
| C18:1n9 | 0.33 [0.23, 0.43] | 0.17 [0.07, 0.26] | 0.05 [−0.01, 0.10] |
| MUFA | 0.35 [0.24, 0.46] | 0.46 [0.33, 0.60] | 0.03 [0, 0.07] |
| C18:2n6 | 0.46 [0.35, 0.57] | 0.31 [0.18, 0.44] | 0.12 [0.05, 0.18] |
| C18:3n3 | 0.50 [0.32, 0.67] | 0.33 [0.19, 0.47] | 0.18 [0.10, 0.26] |
| C20:4n6 | 0.38 [0.20, 0.56] | 0.20 [0.05, 0.36] | 0.11 [0.05, 0.18] |
| PUFA | 0.43 [0.30, 0.55] | 0.17 [0.08, 0.27] | 0.09 [0.02, 0.15] |
| Trait | d 1 | m 2 | dm 3 |
|---|---|---|---|
| C14:0 | 0.96 [0.90, 1.00] | 0.68 [0.27, 1.00] | −0.21 [−0.60, 0.18] |
| C16:0 | 0.69 [0.46, 0.91] | −0.02 [−0.56, 0.52] | −0.66 [−0.94, −0.39] |
| C18:0 | −0.81 [−0.98, −0.64] | −0.33 [−0.81, 0.16] | −0.15 [−0.61, 0.32] |
| SFA | 0.97 [0.88, 1.00] | −0.03 [−0.55, 0.49] | −0.89 [−1.00, −0.74] |
| C16:1n7 | 0.92 [0.81, 1.00] | 0.74 [0.50, 0.97] | −0.39 [−0.73, −0.05] |
| C18:1n9 | 0.36 [0.15, 0.57] | 0.68 [0.44, 0.91] | −0.66 [−0.83, −0.49] |
| MUFA | 0.40 [0.19, 0.61] | 0.52 [0.17, 0.88] | −0.53 [−0.69, −0.37] |
| C18:2n6 | −0.27 [−0.51, −0.03] | −0.13 [−0.57, 0.31] | −0.54 [−0.77, −0.31] |
| C18:3n3 | 0.52 [0.20, 0.84] | 0.33 [−0.39, 1.00] | −0.38 [−0.82, 0.06] |
| C20:4n6 | −0.63 [−0.84, −0.42] | −0.81 [−1.00, −0.61] | 0.15 [−0.30, 0.60] |
| PUFA | −0.51 [−0.69, −0.32] | −0.19 [−0.58, 0.20] | −0.56 [−0.72, −0.40] |
| OCU | Map Position (bp) | Trait (% of Explained Variance 2) | No. of SNPs 3 |
|---|---|---|---|
| 1 | 35,902,181–37,321,725 | PUFA/SFA (13.9%) | 50 |
| 1 | 35,902,181–37,337,488 | C16:0 (12.2%) | 48 |
| 1 | 35,902,181–37,337,488 | C16:1n7 (9.9%) | 43 |
| 1 | 35,902,181–37,337,488 | PUFA (3.2%) | 44 |
| 5 | 10,141,256–10,568,012 | C16:0 (3.5%) | 12 |
| 5 | 10,141,256–10,568,012 | SFA (5.1%) | 12 |
| 5 | 10,141,256–10,568,012 | PUFA/SFA (4.3%) | 12 |
| 5 | 10,161,032–10,568,012 | C18:2n6 (3.8%) | 8 |
| 19 | 48,610,554–50,061,214 | IMF (8.0%) | 36 |
| 19 | 49,573,782–49,993,977 | C20:4n6 (1.2%) | 21 |
| 20 | 16,022,958–16,482,079 | C20:4n6 (1.6%) | 19 |
| Trait | OCU | Map Position (bp) | No. of Genes | PCG 1 | |
|---|---|---|---|---|---|
| Start | End | ||||
| IMF | 19 | 48,000,588 | 50,994,791 | 41 | TANC2, CYB561, ACE, ENSOCUG00000034369, KCNH6, DCAF7, MAP3K3, LIMD2, STRADA, CCDC47, DDX42, FTSJ3, ENSOCUG00000004329, SMARCD2, ENSOCUG00000026494, GHB1, CD79B, SCN4A, ENSOCUG00000035912, ICAM2, ENSOCUG00000029995, ENSOCUG00000004362, TEX2, ENSOCUG00000034748, PECAM1, ENSOCUG00000007896, POLG2, DDX5, CEP95, SMURF2, ENSOCUG00000023204, ENSOCUG00000015913, BPTF, NOL11, PITPNC1, PSMD12, HELZ, ENSOCUG00000008850, CACNG4, CACNG5, PRKCA |
| C16:0 | 1 | 35,072,649 | 36,992,566 | 4 | ADAMTSL1, SH3GL2, CNTLN, BNC2 |
| 5 | 10,022,607 | 10,999,540 | 2 | RPGRIP1L, FTO | |
| SFA | 1 | 36,002,870 | 36,992,566 | 2 | SH3GL2, CNTLN |
| 5 | 10,022,607 | 10,999,540 | 2 | RPGRIP1L, FTO | |
| C16:1n7 | 1 | 35,072,649 | 37,977,724 | 4 | ADAMTSL1, SH3GL2, CNTLN, BNC2 |
| PUFA | 1 | 35,072,649 | 37,977,724 | 4 | ADAMTSL1, SH3GL2, CNTLN, BNC2 |
| 5 | 10,022,607 | 10,999,540 | 2 | RPGRIP1L, FTO | |
| PUFA/SFA | 1 | 35,017,581 | 37,620,996 | 4 | ADAMTSL1, SH3GL2, CNTLN, BNC2 |
| 5 | 9,928,946 | 10,444,669 | 2 | RPGRIP1L, FTO | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
EL Nagar, A.G.; Heddi, I.; Sosa-Madrid, B.S.; Blasco, A.; Hernández, P.; Ibáñez-Escriche, N. Genome-Wide Association Study of Maternal Genetic Effects on Intramuscular Fat and Fatty Acid Composition in Rabbits. Animals 2023, 13, 3071. https://doi.org/10.3390/ani13193071
EL Nagar AG, Heddi I, Sosa-Madrid BS, Blasco A, Hernández P, Ibáñez-Escriche N. Genome-Wide Association Study of Maternal Genetic Effects on Intramuscular Fat and Fatty Acid Composition in Rabbits. Animals. 2023; 13(19):3071. https://doi.org/10.3390/ani13193071
Chicago/Turabian StyleEL Nagar, Ayman G., Imen Heddi, Bolívar Samuel Sosa-Madrid, Agustín Blasco, Pilar Hernández, and Noelia Ibáñez-Escriche. 2023. "Genome-Wide Association Study of Maternal Genetic Effects on Intramuscular Fat and Fatty Acid Composition in Rabbits" Animals 13, no. 19: 3071. https://doi.org/10.3390/ani13193071