
Exploring Immunohistochemistry in Fish: Assessment of Antibody Reactivity by Western Immunoblotting

 , , , ,
, , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Histology
2.2. Antibodies and Sequence Alignments
2.3. SDS–PAGE and Western Immunoblotting
2.4. Immunohistochemistry
3. Results
3.1. Antibodies and Sequence Alignments
3.2. Pan-Cytokeratin
3.3. Vimentin
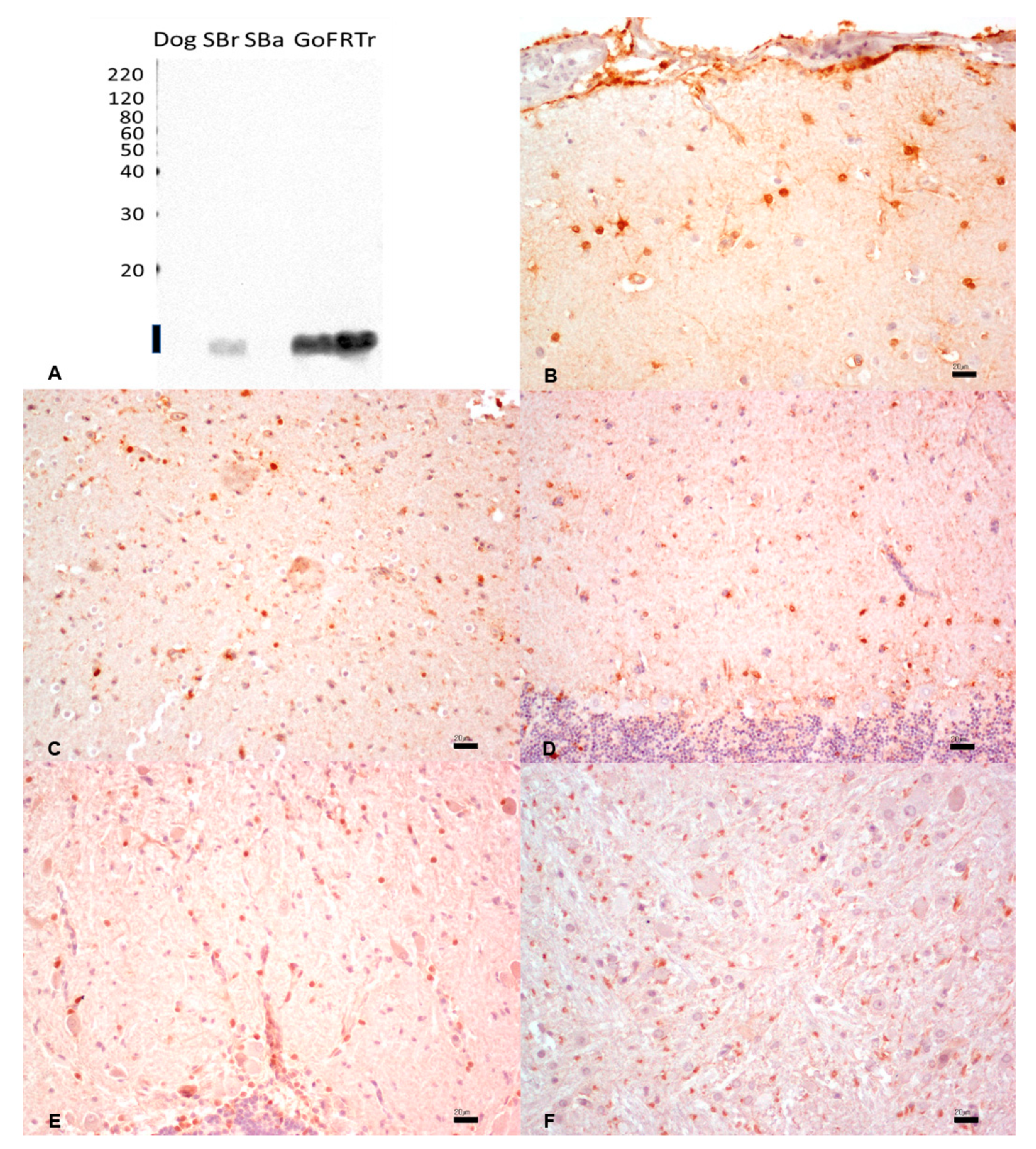
3.4. S100 Protein
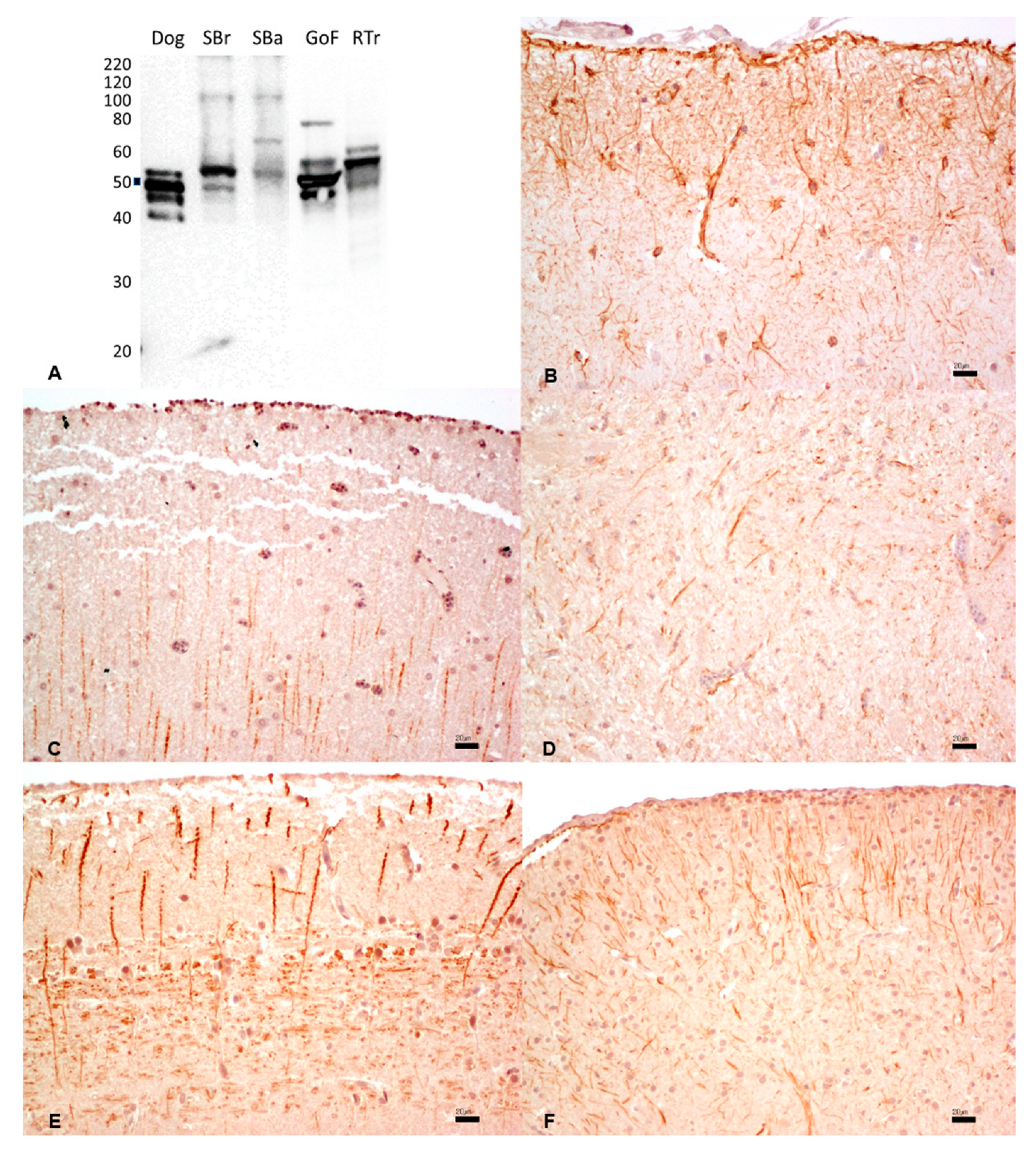
3.5. Glial Fibrillary Acidic Protein
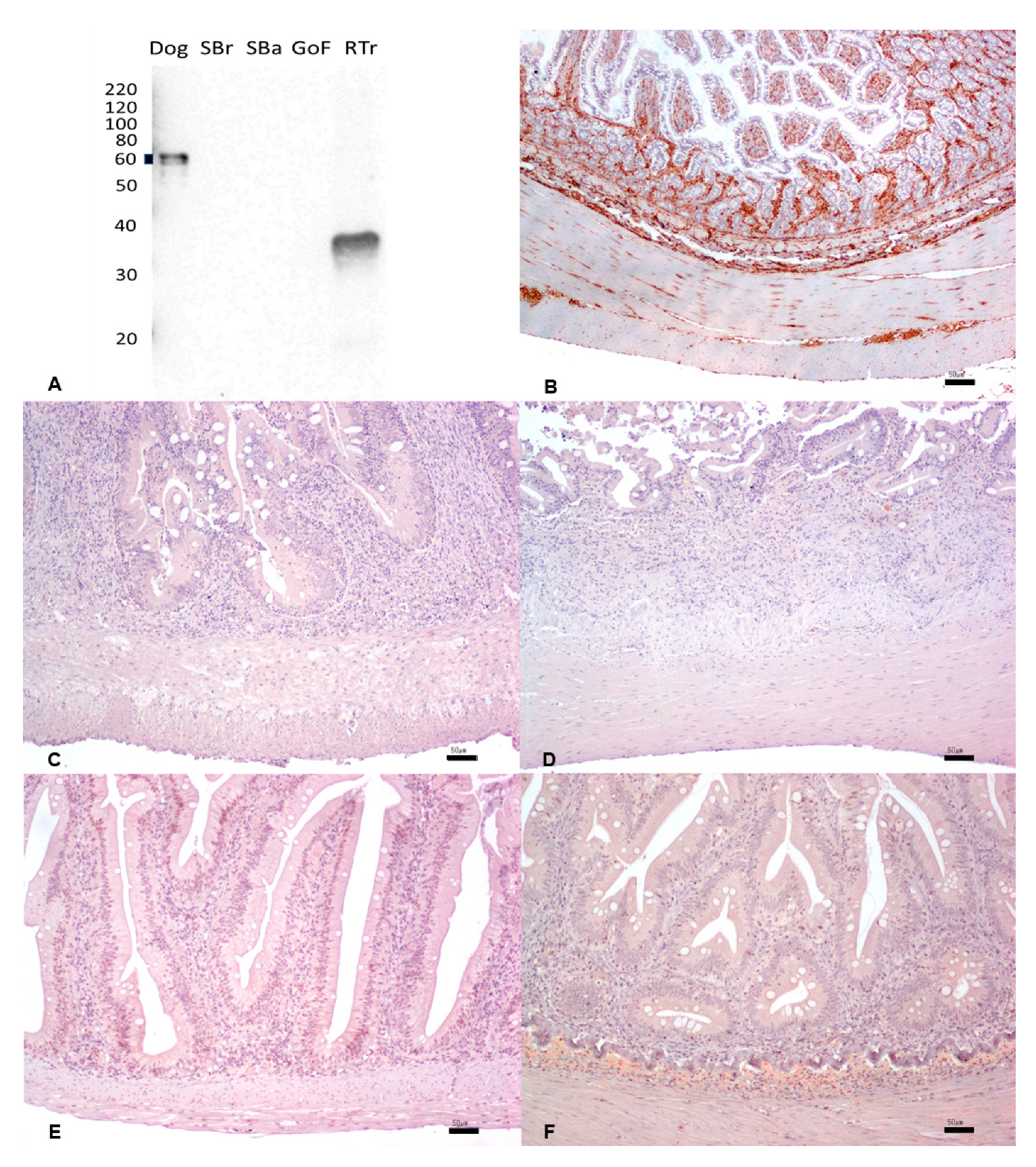
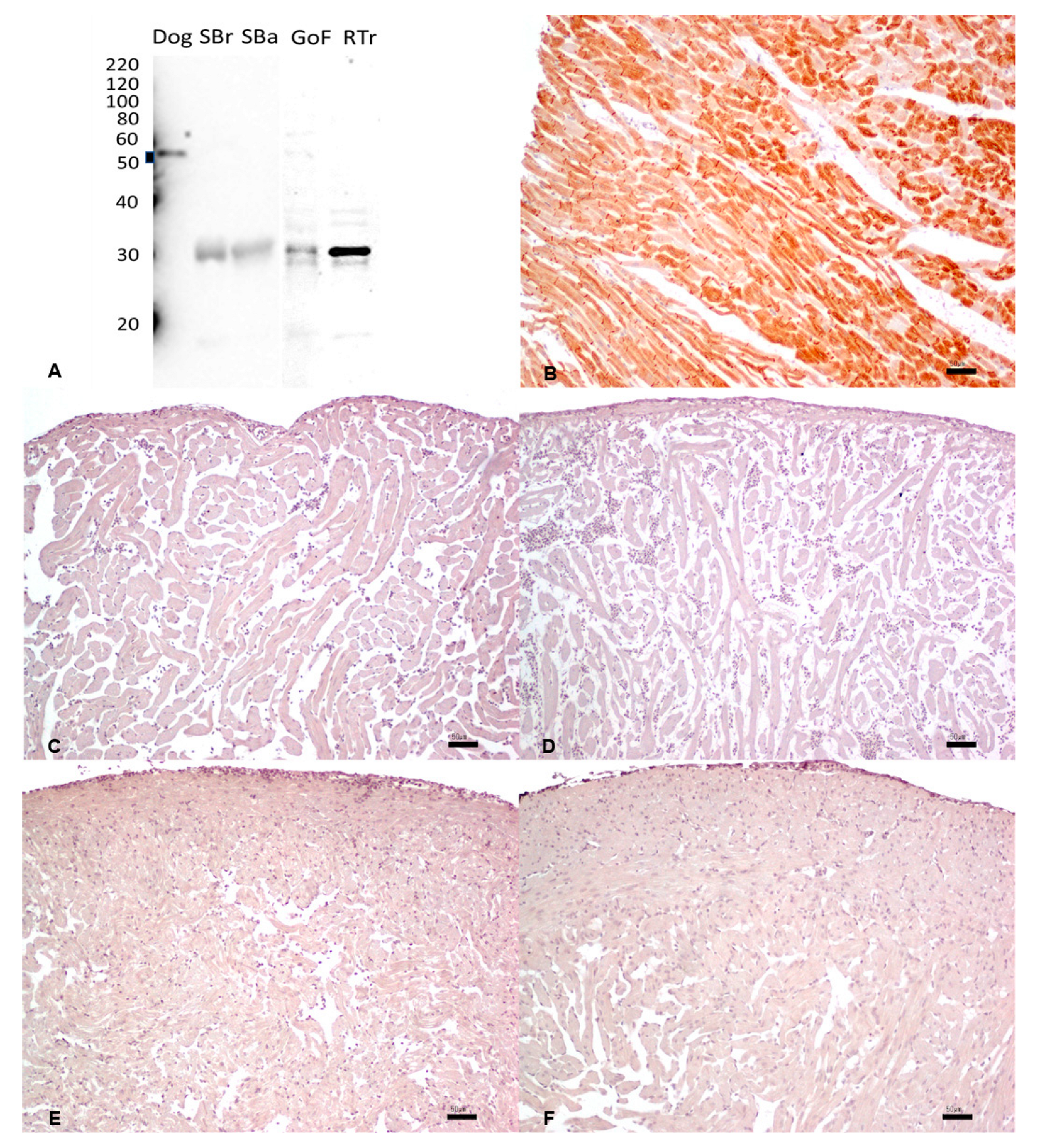
3.6. Desmin


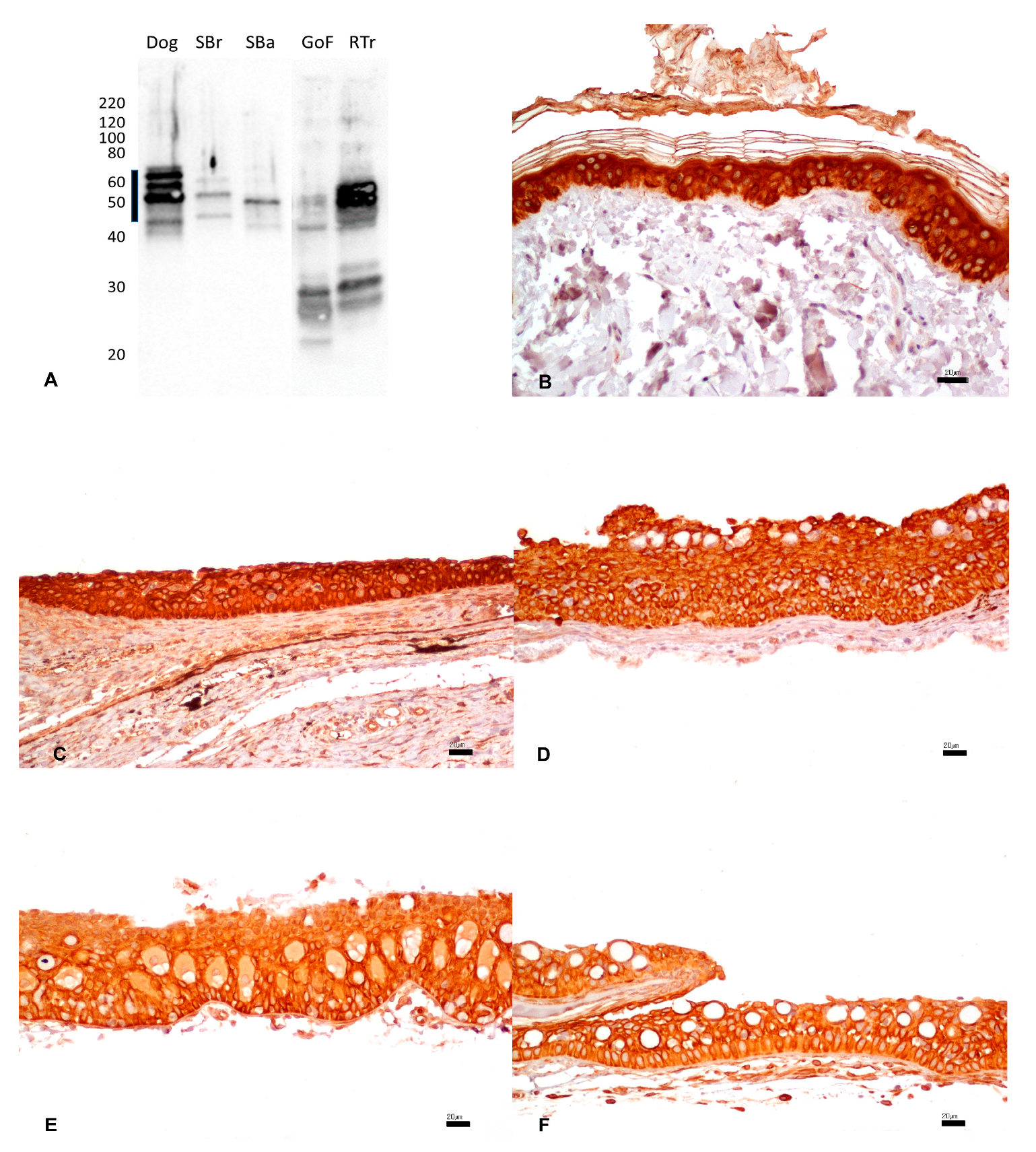{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody and Tissue | Species | WB | IHC |
|---|---|---|---|
| Pan-cytokeratin (skin) Monoclonal mouse anti-human cytokeratin CKAE1/AE3 | Canine | + | + |
| Sparus aurata | + | + | |
| Dicentrarchus labrax | + | + | |
| Carassius auratus | + | + | |
| Oncorhynchus mykiss | + | + | |
| Vimentin (intestine) Monoclonal mouse anti-vimentin, clone V9 | Canine | + | + |
| Sparus aurata | - | - | |
| Dicentrarchus labrax | - | - | |
| Carassius auratus | - | - | |
| Oncorhynchus mykiss | × | - | |
| S-100 (brain) Polyclonal rabbit, code Z0311 | Canine | - | + |
| Sparus aurata | + | + | |
| Dicentrarchus labrax | - | + | |
| Carassius auratus | + | + | |
| Oncorhynchus mykiss | + | + | |
| GFAP (brain) Polyclonal rabbit anti-glial fibrillary acidic protein, code Z0334 | Canine | + | + |
| Sparus aurata | + | + | |
| Dicentrarchus labrax | + | + | |
| Carassius auratus | + | + | |
| Oncorhynchus mykiss | + | + | |
| Desmin (skin) Mouse monoclonal anti-human desmin, clone D33 | Canine | + | + |
| Sparus aurata | × | - | |
| Dicentrarchus labrax | × | - | |
| Carassius auratus | × | - | |
| Oncorhynchus mykiss | × | - |
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramos-Vara, J.A.; Miller, M.A. When Tissue Antigens and Antibodies Get Along. Veter. Pathol. 2013, 51, 42–87. [Google Scholar] [CrossRef] [PubMed]
- Markl, J.; Franke, W.W. Localization of cytokeratins in tissues of the rainbow trout: Fundamental differences in expression pattern between fish and higher vertebrates. Differentiation 1988, 39, 97–122. [Google Scholar] [CrossRef] [PubMed]
- Bunton, T.E. The Immunocytochemistry of Cytokeratin in Fish Tissues. Veter. Pathol. 1993, 30, 418–425. [Google Scholar] [CrossRef]
- Germanà, A.; Marino, F.; Guerrera, M.C.; Campo, S.; de Girolamo, P.; Montalbano, G.; Germanà, G.; Ochoa-Erena, F.J.; Ciriaco, E.; Vega, J.A. Expression and distribution of S100 protein in the nervous system of the adult zebrafish (Danio rerio). Microsc. Res. Tech. 2008, 71, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Iaria, C.; Ieni, A.; Corti, I.; Puleio, R.; Brachelente, C.; Mazzullo, G.; Lanteri, G. Immunohistochemical Study of Four Fish Tumors. J. Aquat. Anim. Health 2019, 31, 97–106. [Google Scholar] [CrossRef]
- Ronza, P.; Villamarín, A.; Méndez, L.; Pardo, B.G.; Bermúdez, R.; Quiroga, M.I. Immunohistochemical expression of E–cadherin in different tissues of the teleost fish Scophthalmus maximus. Aquaculture 2019, 501, 465–472. [Google Scholar] [CrossRef]
- Šálková, E.; Schmidt-Posthaus, H.; Lutz, I.; Kroupová, H.K.; Steinbach, C. Immunohistochemical investigation of epithelial, mesenchymal, neuroectodermal, immune and endocrine markers in sterlet (Acipenser ruthenus), shortnose sturgeon (Acipenser brevirostrum) and common carp (Cyprinus carpio). Fish Physiol. Biochem. 2022, 48, 1737–1749. [Google Scholar] [CrossRef]
- Romano, L.A.; Pedrosa, V.F. Neoplasias in Fish: Review of the Last 20 Years. A Look from the Pathology. Annu. Res. Rev. Biol. 2020, 35, 134–153. [Google Scholar] [CrossRef]
- Paquette, C.E.; Kent, M.L.; Peterson, T.S.; Wang, R.; Dashwood, R.H.; Löhr, C.V. Immunohistochemical characterization of intestinal neoplasia in zebrafish Danio rerio indicates epithelial origin. Dis. Aquat. Org. 2015, 116, 191–197. [Google Scholar] [CrossRef]
- Quaglio, F.; Zappulli, V.; Poppi, L.; Capovilla, P.; Capparucci, F.; Marino, F. Squamous cell carcinoma in a wild European bullhead Cottus gobio. Dis. Aquat. Org. 2016, 122, 73–76. [Google Scholar] [CrossRef]
- Rahmati-Holasoo, H.; Shokrpoor, S.; Masoudifard, M.; Davudypoor, S.; Vaseghi, M. Telangiectatic osteosarcoma and renal adenocarcinoma in an Oscar (Astronotus ocellatus, Agassiz): Diagnostic imaging and immunohistochemical study. J. Fish Dis. 2018, 41, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Stilwell, J.M.; McManamon, R.; Sturgeon, G.L.; Camus, M.S.; Camus, A.C. Microscopic, immunohistochemical and ultrastructural characterization of spontaneous lepidosarcomas in a neon tetra, Paracheirodon innesi, and a black crappie, Pomoxis nigromaculatus. J. Fish Dis. 2019, 42, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Denk, D.; Verin, R.; Ressel, L.; Lewis, E.; Stidworthy, M.F. Spontaneous neoplasia in captive syngnathid species: A retrospective case series (2003–2014) and literature review. J. Fish Dis. 2020, 43, 929–939. [Google Scholar] [CrossRef] [PubMed]
- Galeotti, M.; Sarli, G.; Sirri, R.; Mandrioli, L.; Beraldo, P.; Bronzatti, P.; Giavenni, R.; Orioles, M.; Magi, G.E.; Volpatti, D. Red mark syndrome of trout (Oncorhynchus mykiss; Walbaum, 1792): Histopathological scoring and correlation with gross lesions. J. Fish Dis. 2021, 44, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Sirri, R.; Tura, G.; Budai, J.; Beraldo, P.; Fiorentino, M.; Barbé, T.; Galeotti, M.; Sarli, G.; Mandrioli, L. Histological and immunohistochemical characterization of 17 gonadal tumours in koi carp (Cyprinus carpio koi). J. Fish Dis. 2020, 44, 273–285. [Google Scholar] [CrossRef]
- Polinas, M.; Padrós, F.; Merella, P.; Prearo, M.; Sanna, M.A.; Marino, F.; Burrai, G.P.; Antuofermo, E. Stages of Granulomatous Response Against Histozoic Metazoan Parasites in Mullets (Osteichthyes: Mugilidae). Animals 2021, 11, 1501. [Google Scholar] [CrossRef]
- Stilwell, J.M.; Boylan, S.M.; Vorbach, B.; Camus, A.C. Epizootic neoplasia in a managed population of Atlantic bumper fish, Chloroscombrus chrysurus (Osteichthyes: Carangidae), at a public aquarium. J. Fish Dis. 2020, 44, 415–427. [Google Scholar] [CrossRef]
- Grunow, B.; Böhmert, B.; Fechner, K. Specificity of antibodies established from mammals in rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2013, 29, 1129–1133. [Google Scholar] [CrossRef]
- Hewitt, S.M.; Baskin, D.G.; Frevert, C.W.; Stahl, W.L.; Rosa-Molinar, E. Controls for Immunohistochemistry. J. Histochem. Cytochem. 2014, 62, 693–697. [Google Scholar] [CrossRef]
- Webster, J.D.; Solon, M.; Gibson-Corley, K.N. Validating Immunohistochemistry Assay Specificity in Investigative Studies: Considerations for a Weight of Evidence Approach. Veter. Pathol. 2020, 58, 829–840. [Google Scholar] [CrossRef]
- Faílde, L.D.; Bermúdez, R.; Vigliano, F.; Coscelli, G.A.; Quiroga, M.I. Morphological, immunohistochemical and ultrastructural characterization of the skin of turbot (Psetta maxima L.). Tissue Cell 2014, 46, 334–342. [Google Scholar] [CrossRef]
- Yasumoto, S.; Koga, D.; Tanaka, K.; Kondo, M.; Takahashi, Y. Histopathological and Immunohistochemical Studies of Gonadal Undifferentiated Carcinoma in Common Carp Cyprinus carpio. Fish Pathol. 2015, 50, 53–59. [Google Scholar] [CrossRef]
- Lanteri, G.; Ieni, A.; Toffan, A.; Abbate, J.; Saraò, M.; Barresi, V.; Macrì, B. Immunohistochemical patterns of a non-viral papilloma in goldfish (Carassius auratus, L.). Bull. Eur. Assoc. Fish Pathol. 2016, 36, 208–213. [Google Scholar]
- Herrmann, H.; Münick, M.D.; Brettel, M.; Fouquet, B.; Markl, J. Vimentin in a cold-water fish, the rainbow trout: Highly conserved primary structure but unique assembly properties. J. Cell Sci. 1996, 109, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Moore, B.W. A soluble protein characteristic of the nervous system. Biochem. Biophys. Res. Commun. 1965, 19, 739–744. [Google Scholar] [CrossRef]
- Fonseca, V.G.; Rosa, J.; Laizé, V.; Gavaia, P.J.; Cancela, M.L. Identification of a new cartilage-specific S100-like protein up-regulated during endo/perichondral mineralization in gilthead seabream. Gene Expr. Patterns 2011, 11, 448–455. [Google Scholar] [CrossRef]
- Abbate, F.; Catania, S.; Germanà, A.; González, T.; Diaz-Esnal, B.; Germanà, G.; Vega, J.A. S-100 protein is a selective marker for sensory hair cells of the lateral line system in teleosts. Neurosci. Lett. 2002, 329, 133–136. [Google Scholar] [CrossRef]
- Sirri, R.; Diana, A.; Scarpa, F.; Brachelente, C.; Vitellozzi, G.; Ceredi, L.; Mandrioli, L. Ultrasonographic and pathologic study of schwannoma in a Goldfish (Carassius auratus). Veter. Clin. Pathol. 2015, 44, 586–591. [Google Scholar] [CrossRef]
- Ramos-Vara, J.A.; Borst, L.B. Immunohistochemistry: Fundamentals and Applications in Oncology. In Tumors in Domestic Animals, 5th ed.; Meuten, D.J., Ed.; Wiley Blackwell: Ames, IA, USA, 2017; pp. 44–87. [Google Scholar]
- Kálmán, M. Astroglial architecture of the carp (Cyprinus carpio) brain as revealed by immunohistochemical staining against glial fibrillary acidic protein (GFAP). Anat. Embryol. 1998, 198, 409–433. [Google Scholar] [CrossRef]
- Hagström, C.; Olsson, C. Glial cells revealed by GFAP immunoreactivity in fish gut. Cell Tissue Res. 2010, 341, 73–81. [Google Scholar] [CrossRef]
- Marino, F.; Lanteri, G.; Rapisarda, G.; Perillo, A.; Macrì, B. Spontaneous schwannoma in zebrafish, Danio rerio (Hamilton). J. Fish Dis. 2012, 35, 239–242. [Google Scholar] [CrossRef]
- Rowlerson, A.; Radaelli, G.; Mascarello, F.; Veggetti, A. Regeneration of skeletal muscle in two teleost fish: Sparus aurata and Brachydanio rerio. Cell Tissue Res. 1997, 289, 311–322. [Google Scholar] [CrossRef]
- Clemen, C.S.; Herrmann, H.; Strelkov, S.V.; Schröder, R. Desminopathies: Pathology and mechanisms. Acta Neuropathol. 2012, 125, 47–75. [Google Scholar] [CrossRef]
- Roccabianca, P.; Schulman, Y.; Avallone, G.; Foster, R.; Scruggs, J.; Dittmer, K.; Kiupel, M. Surgical Pathology of Tumors of Domestic Animals. 3: Tumors of Soft Tissue; Davis Thompson Foundation: Gurnee, IL, USA, 2020; ISBN 9781733749121. [Google Scholar]
- Armando, F.; Pigoli, C.; Gambini, M.; Ghidelli, A.; Ghisleni, G.; Corradi, A.; Passeri, B.; Caniatti, M.; Grieco, V.; Baumgärtner, W.; et al. Peripheral Nerve Sheath Tumors Resembling Human Atypical Neurofibroma in Goldfish (Carassius auratus, Linnaeus, 1758). Animals 2021, 11, 2621. [Google Scholar] [CrossRef] [PubMed]
- Alunni, A.; Vaccari, S.; Torcia, S.; Meomartini, M.E.; Nicotra, A.; Alfei, L. Characterization of glial fibrillary acidic protein and astroglial architecture in the brain of a continuously growing fish, the rainbow trout. Eur. J. Histochem. 2005, 49, 157–166. [Google Scholar] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antuofermo, E.; Orioles, M.; Murgia, C.; Burrai, G.P.; Penati, M.; Gottardi, C.; Polinas, M.; Volpatti, D.; Galeotti, M.; Addis, M.F. Exploring Immunohistochemistry in Fish: Assessment of Antibody Reactivity by Western Immunoblotting. Animals 2023, 13, 2934. https://doi.org/10.3390/ani13182934
Antuofermo E, Orioles M, Murgia C, Burrai GP, Penati M, Gottardi C, Polinas M, Volpatti D, Galeotti M, Addis MF. Exploring Immunohistochemistry in Fish: Assessment of Antibody Reactivity by Western Immunoblotting. Animals. 2023; 13(18):2934. https://doi.org/10.3390/ani13182934
Chicago/Turabian StyleAntuofermo, Elisabetta, Massimo Orioles, Claudio Murgia, Giovanni P. Burrai, Martina Penati, Chiara Gottardi, Marta Polinas, Donatella Volpatti, Marco Galeotti, and Maria Filippa Addis. 2023. "Exploring Immunohistochemistry in Fish: Assessment of Antibody Reactivity by Western Immunoblotting" Animals 13, no. 18: 2934. https://doi.org/10.3390/ani13182934