

The Fergana Valley Is an Isolate of Biodiversity: A Discussion of the Endemic Herpetofauna and Description of Two New Species of Alsophylax (Sauria: Gekkonidae) from Eastern Uzbekistan
 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. The Current Status of Conservation in Fergana Valley
1.2. Current Taxonomy of the Gecko Genus Alsophylax
2. Materials and Methods
2.1. Population Density and Field Observations
2.2. Molecular Data and Phylogenetic Analyses
2.3. Morphological Data and Statistical Analyses
3. Results
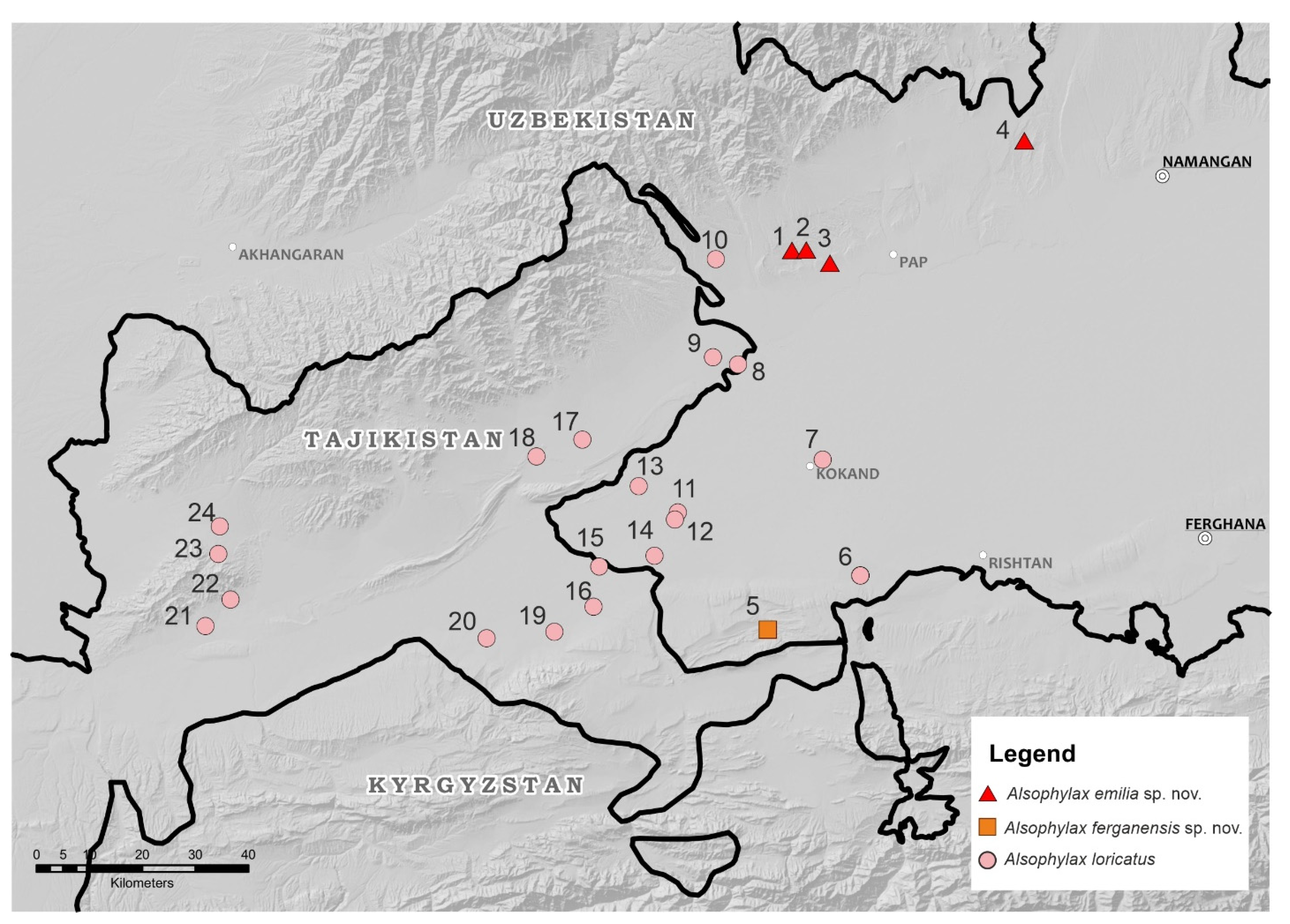
3.1. Population Density and Distribution
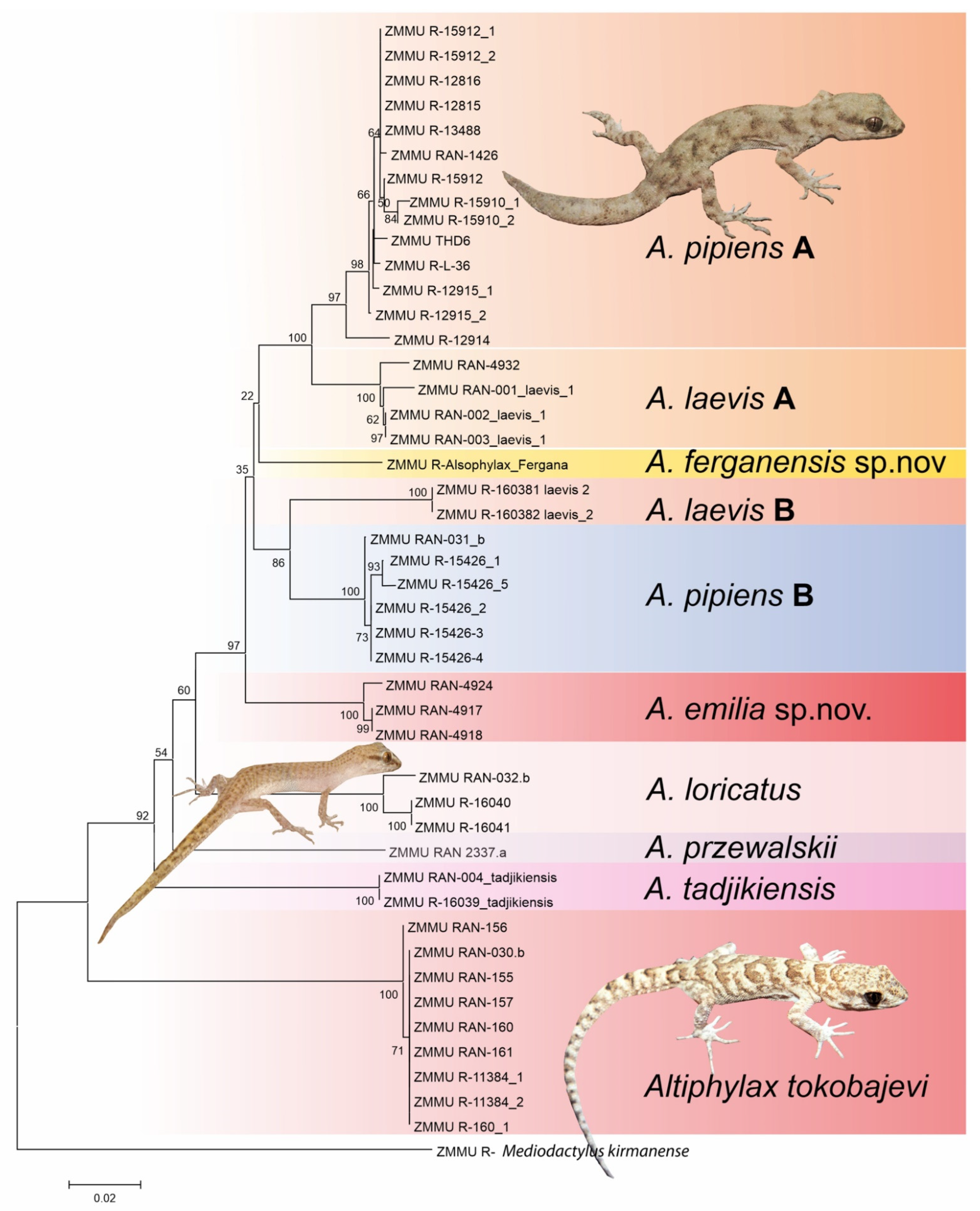
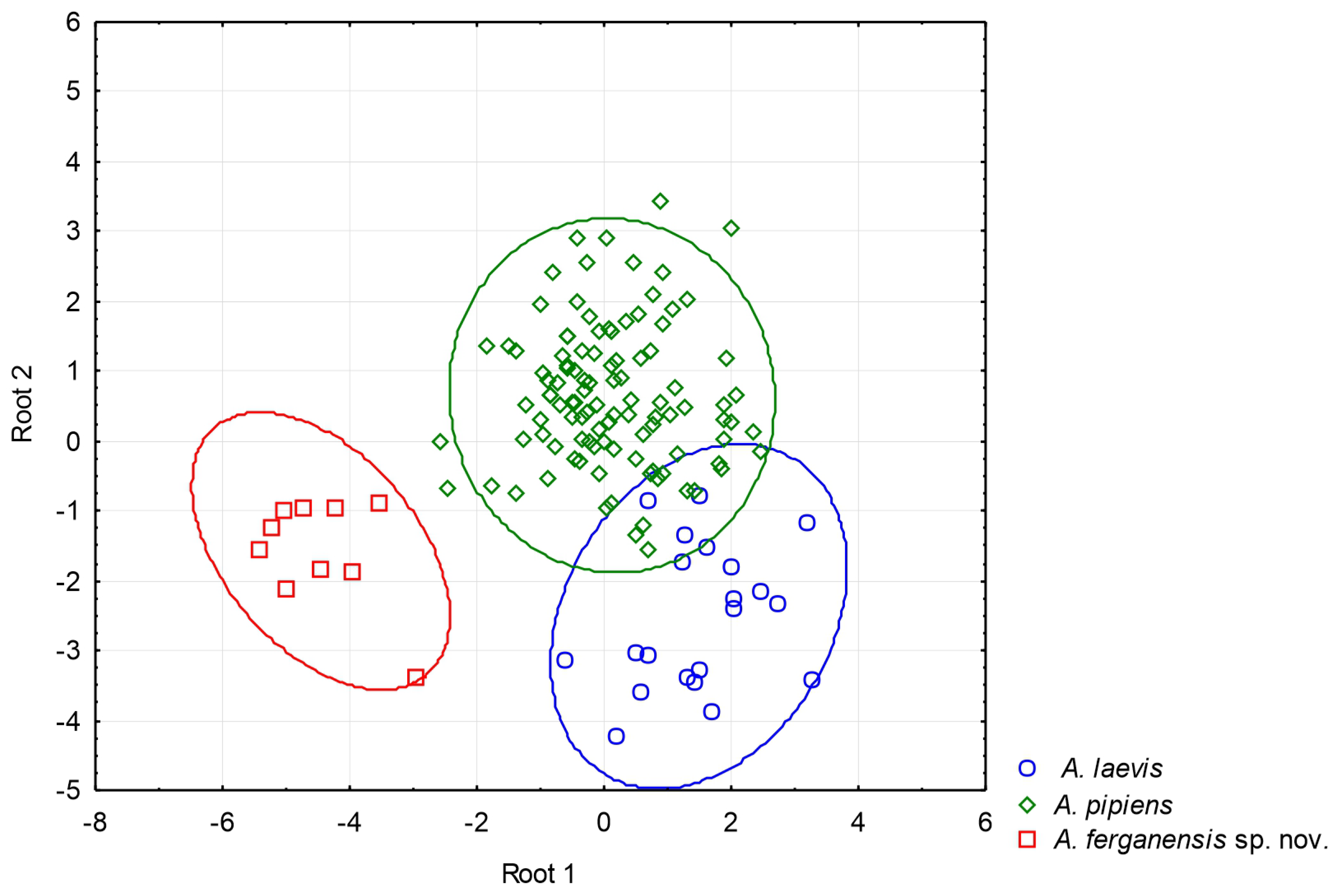
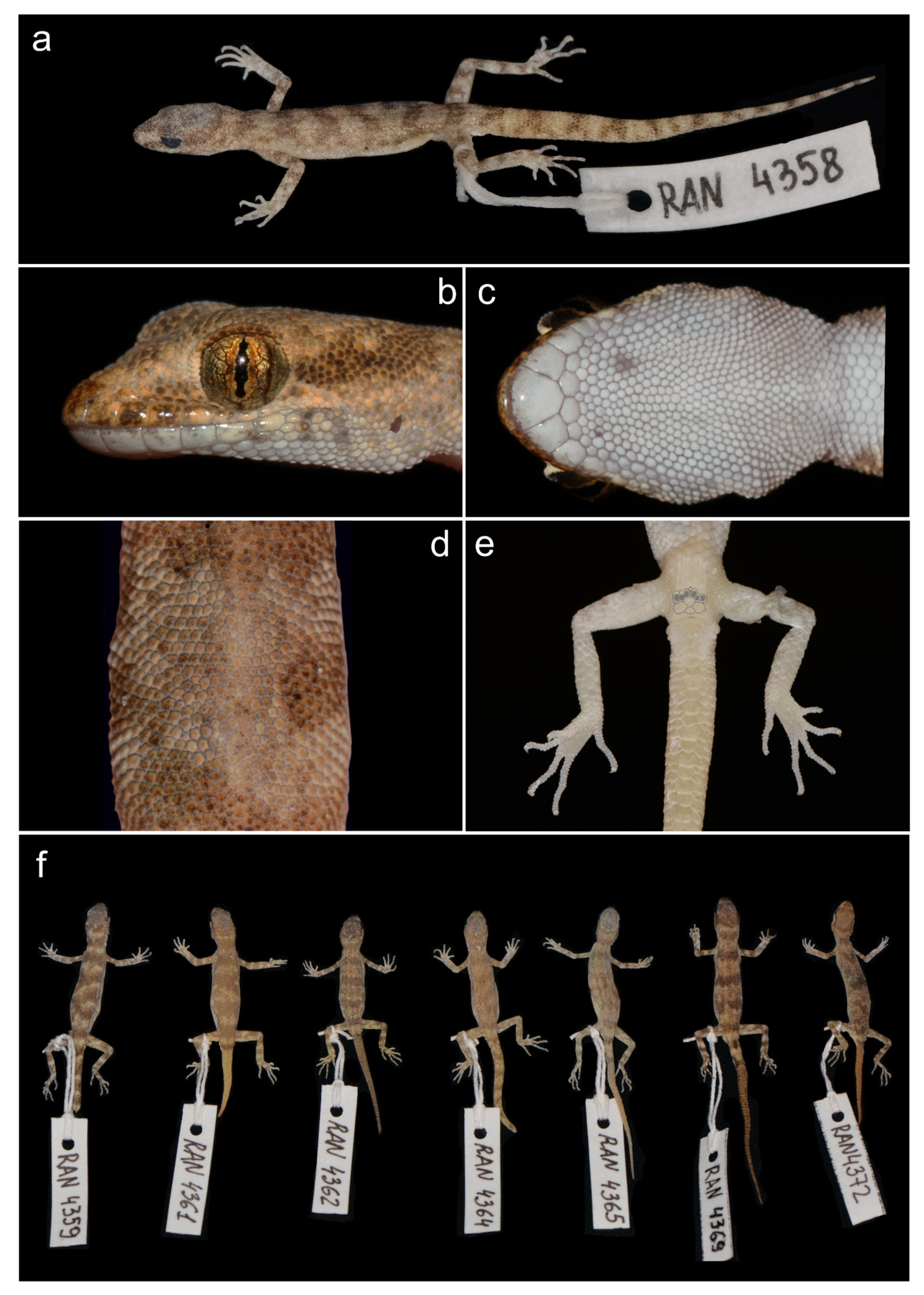
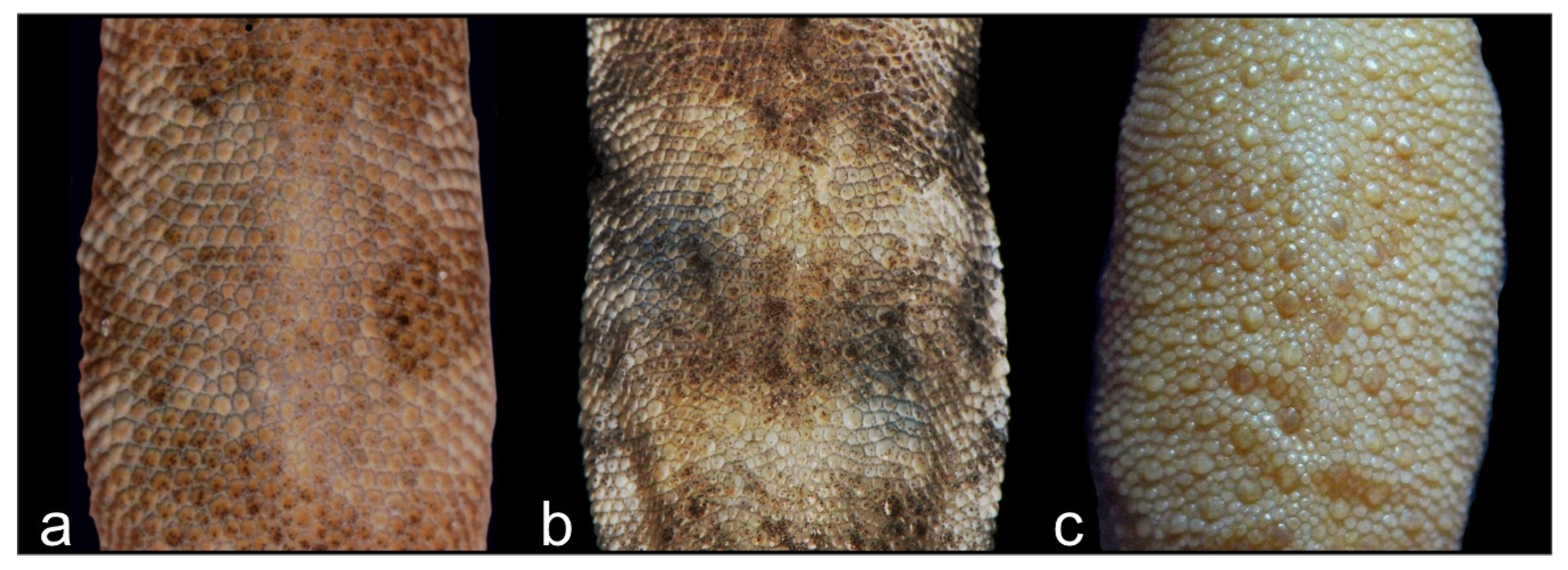
3.2. Molecular Analysis and Morphology
4. Discussion
4.1. Taxonomy of Alsophylax and Conservation in the Fergana Valley
4.2. The History of Herpetological Study in Central Asia and the Fergana Valley
- −
- H. C. Lichtenstein published a list of 18 species of reptiles (including five new ones) and two species of amphibians [31].
- −
- A. Lehmann in July 1841 crossed the Kuvan-Darya and the Kyzylkum desert, and then visited Samarkand and Bukhara. From there, he traveled to the mountains to Iskenderun and went to Orenburg, Kazakhstan [32].
- −
- −
- A. P. Fedchenko in 1871 worked in the Fergana Valley and visited Kokand, Andijon, Namangan, Vuadil, Uch-Kurgan, and Besh-Aryk. The works by Fedchenko [39,40,41] included only part of the materials he collected in Central Asia and the Fergana Valley. The remaining collections were mainly used by A. M. Nikolsky and A. A. Strauch in their works.
- −
- Strauch published a list of geckos from Central Asia, compiled their characteristics, and described a number of new species in Central Asia. Of greatest interest are descriptions of the Turkestan thin-toed gecko, Transcaspian bent-toed gecko, and Strauch’s even-fingered gecko from Uzbekistan and other republics of Central Asia [42].
- −
- Nikolsky published “Reptiles and amphibians of the Turkestan general governorship” and gave a complete literary summary at that time [34]. The author processed the materials of A. P. Fedchenko and others who collected in Central Asia. The paper provides information on the distribution of seven species of amphibians, three species of turtles, 42 species of lizards, and 28 species of snakes. Of these, more than half were collected in Uzbekistan [34].
- −
- Bogdanov published his work “Fauna of the Uzbek SSR. Amphibians and reptiles” [30] and provided all the data that he collected during his time doing field work in the Namangan region during 1949, 1950, 1954, and 1955.
- −
- −
- −
- O.I. Tsaruk paid great attention to the problems and conservation of the unique ecosystems of the Fergana Valley, studying rare and endemic species of Phrynocephalus in the Fergana Valley [48].
- −
- −
- Bondarenko and Peregontsev (2017) described the spatial distribution of the Central Asian Tortoise, Testudo horsfieldii in Uzbekistan, including the Fergana Valley [51].
- −
4.3. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Collection Number | N | E | H (m) | Locality |
|---|---|---|---|---|---|
| A. laevis | ZMMU R-6643 | 37.4139 | 65.0347 | 244 | Turkmenistan, Chardzhou region, eastern part of the Obruchev Steppe, vicinity of the Yusun well |
| ZMMU R-7280 1–2 | 40.697583 | 63.659622 | Uzbekistan, Bukhara region, foothills of Kuldzhuk-tau, 10 km west. Shuruk vill. | ||
| ZMMU R-4125 | 37.271453 | 60.245347 | Turkmenistan, Takhir RW station | ||
| ZMMU R-7281 1–2 | 40.546495 | 63.790836 | Uzbekistan, Bukhara region, 15 km south Shuruk vill. | ||
| ZMMU R-5895 | 38.046054 | 58.035237 | Turkmenistan, Gekdere vill. | ||
| ZMMU R-5940 | 39.213124 | 55.709714 | Turkmenistan, 10 km from Uzun-Su RW station | ||
| ZMMU R-16038 | 39.508916 | 54.362538 | Turkmenistan, 18 km northwest of Kyzyl Arvat (Sedar city), env. Qishlaq Aji | ||
| ZISP—18936 1–13 | 37.271453 | 60.245347 | Turkmenistan, station Takyr | ||
| ZISP—19177 1–15 | 37.371190 | 60.077820 | Turkmenistan, 33 km west of Tejen on the highway to Ashgabat | ||
| ZISP—16612 | 37.261158 | 67.307407 | Uzbekistan, Kara-su near Termez | ||
| ZISP—18925 1–27 | 39.015630 | 56.253282 | Turkmenistan, 4 km northwest of Kizyl-Arvat | ||
| IZUAS—1473 1–6 | 40.682777 | 64.010022 | Uzbekistan, 40 km to the west of Ayagitma (Kyzylkum) | ||
| IZUAS—1474 1–13 | 40.813600 | 64.336812 | Uzbekistan, 20 km to the northwest of Ayagitma | ||
| IZUAS—1574 1–2 | 40.682777 | 64.010022 | Uzbekistan, 20 km to the west of Ayagitma | ||
| A. loricatus | ZMMU R-10679 | 40.283611 | 69.623962 | Tajikistan, Leninabad, old fortress. | |
| ZMMU R-16041 | 40.289225 | 70.426283 | Tajikistan, Ferghana Valley, env. Kanibadam | ||
| ZMMU R-11424 1–5 | 40.319699 | 71.024406 | Uzbekistan, Ferghana Valley, env. Kokand city, Sarykurgan village on the Sokh river. Uzbek-Kyrgyz border. | ||
| ZMMU R-16105 1–6 | 40.535786 | 70.940439 | Uzbekistan, Ferghana Valley, environs of Kokand | ||
| ZMMU R-16106 1–3 | 40.331407 | 71.021404 | Uzbekistan, Fergana Valley, the bank of the Sokh River. Sarykurgan. | ||
| ZMMU R-16104 1–8 | 40.535786 | 70.940439 | Uzbekistan, Ferghana Valley, environs of Kokand | ||
| A. pipiens | ZMMU R-12915 | 44.05 | 69 | 337 | Kazakhstan, western part of the Moinkum Sands |
| ZMMU R-5878 1–9 | 42.757384 | 107.550913 | Mongolia, Galbyn-Gobi (southern), Tavan-Aldyn-Khudun | ||
| ZMMU R-5490 1–2 | 45.659435 | 91.154084 | Mongolia, Kobdos aimag, 15 km NW of the Ushig outpost | ||
| ZMMU R-6088 1–4 | 45.467223 | 106.260296 | Mongolia, Middle Gobi aimag, Bayan-Dov | ||
| ZMMU R-12602 | 44.140570 | 103.715861 | Mongolia, South Gobi aimag, Bayan-zag | ||
| ZMMU R-5881 1–3 | - | - | Mongolia | ||
| ZMMU R-13053 | 45.351402 | 96.647005 | Mongolia, Gobi-Altai aimag, Tsagan-Tolgoin-Bulak, about 30 km on the road from Tsogt to Dzakhoy | ||
| ZMMU R-4253 1–2 | 43.151441 | 109.145179 | Mongolia, Dornogob, 30 km south of Khuvsgul somon, on the road to Khatan-Bulak | ||
| ZMMU R-4924 1–3 | 43.229632 | 101.047568 | Mongolia, South Gobi aimag, vicinity of Gurvan-Tes | ||
| ZMMU R-5491 1–3 | 48.212680 | 102.217947 | Mongolia, Gobi-Altai aimag, Khatsovmiin-bulag, about 50 km S-SE from Khair-Khan | ||
| ZMMU R-5381 1–5 | 42.333333 | 103.25 | 1200 | Mongolia, South Gobi aimag, southern slope of the Mt. Baga-Ula | |
| ZMMU R-5489 | 43.128934 | 97.975124 | Mongolia, Bayan Khongor aimag, Shara-Khulsny-Bulak | ||
| ZMMU R-4920 1–4 | 43.251052 | 100.004418 | Mongolia, South Gobi aimag, Naryn-Bulak | ||
| ZMMU R-6090 | 42.533333 | 106.85 | Mongolia, 5 km to the west from Ganz-Mod-Khuduk | ||
| ZMMU R-5492 1–2 | 45.659435 | 91.154084 | Mongolia, Kobdos aimag, 10 km NW of the Ushig outpost | ||
| ZMMU R-5880 1–2 | 43.716667 | 108.633333 | Mongolia, Dornogovi aimag, in the ruins of a monastery | ||
| ZMMU R-6089 1–2 | 44.25 | 109.283333 | Mongolia, Dornogovi aimag | ||
| ZMMU R-5488 | 46.699222 | 91.069969 | Mongolia, Kobdos aimag, 22 km southwest of the Gurvan Khukhet outpost | ||
| ZMMU R-5254 | 43.667070 | 102.933058 | Mongolia, 150 km NW from Bayan-Leg (northern slope of the Nem-Get ridge) | ||
| ZMMU R-4921 | 43.321384 | 99.329913 | Mongolia, vicinity of the Ekhiin-Gol station | ||
| ZMMU R-5385 | 42.066667 | 104.416667 | 1250 | Mongolia, South Gobi aimag, 5 km NW from the Jal-Shand-Khuduk well | |
| ZMMU R-6056 | 43 | 107.666667 | Mongolia, Aimak Umnegovi | ||
| ZMMU R-7172 | 49.615853 | 116.993728 | China/Mongolia, South Gobi aimag, eastern tip of Bordzingiin-Gobi, 12 km west of Genghis Khan Wall | ||
| ZMMU R-3908 | 43.836195 | 106.549735 | Mongolia, South Gobi aimag, 10 km north of the southeastern tip of the Khurkhe-Ula ridge (an extension of the Gobi Altai) | ||
| ZMMU R-5829 25 | 48.967206 | 89.963372 | Mongolia, the ruins of the monastery Ulgii | ||
| ZMMU R-5382 1–3 | 41.833333 | 104.666667 | 1050 | Mongolia, South Gobi aimag, northern tip of the Dzamyn-Khuren-Eds sands, 10 km SE from the Bichit-Usny-Khuduk well | |
| ZMMU R-4922 | 43.251052 | 100.004418 | Mongolia, Naryn-Dats-Bulak | ||
| ZMMU R-4925 1–2 | 43.229632 | 101.047568 | Mongolia, South Gobi aimag, vicinity of Gurvan-Tes | ||
| ZMMU R-4923 317–318 | 43.909167 | 9.912222 | Mongolia, Haichin-2 | ||
| ZMMU R-12914 | 43.73 | 79.46 | Kazakhstan | ||
| ZMMU R-9571 | 44.052888 | 62.711996 | Kazakhstan, Kzyl-Orda region, 80 km to the northeast from Chaban-Kazgan, northern Kyzyl-Kum | ||
| ZMMU R-9570 1–6 | 43.535957 | 58.688738 | Uzbekistan, at the foot of the Chink near Sudachey Lake, in the lower reaches of the Amu-Darya | ||
| ZMMU R-14716 | 44.94747 | 68.57365 | Kazakhstan, the edge of the floodplain of the Chu River | ||
| ZMMU R-5920 | 43.423373 | 78.993863 | Kazakhstan, Alma-Ata region, 40 km W-SW Chundzha village | ||
| ZMMU R-336 | 45.657446 | 58.602648 | Kazakhstan, NW shore of the Aral Sea, Ust-Urt, Kaska-dzhol tract | ||
| ZMMU R-11423 1 | 43.870681 | 77.051736 | Kazakhstan, surroundings of Kapchagan | ||
| ZMMU R-115 1–3 | 45.915034 | 73.613855 | Kazakhstan, Cape Korzhun-Tubek, western shore of Lake Balkhash | ||
| ZMMU R-118 1–3 | 45.474858 | 73.612777 | Kazakhstan, Zhambyl region, Mynaral tract, western shore of the lake. Balkhash | ||
| ZMMU R-9569 1–2 | 5.252309 | 63.423455 | Kazakhstan, Northern Kyzyl-Kum, 50–70 km SW-W of Dzhusaly station | ||
| ZMMU R-9566 | 44.481389 | 64.168056 | Kazakhstan, Kum-Kala, Karmakchinsky district, Kzyl-Orda region | ||
| ZMMU R-9565 | 45.505092 | 61.612670 | Kazakhstan, Erembet-Tunbugut, Karmakchinsky district, Kzyl-Orda region | ||
| ZMMU R-9567 | 45.597886 | 63.072142 | Kazakhstan, 21 km W from Tunbugut, Karmakchinsky district, Kzyl-Orda region. | ||
| ZMMU R-12917 | 46.733333 | 75.366667 | Kazakhstan, north of Lake Balkhash, kishlak Ortaderessin | ||
| ZMMU R-13488 | 44.066667 | 76.983333 | Kazakhstan, Almaty region, about 18 km northwest of Kapchagay, at the foot of the rocks on the banks of the Ili River | ||
| ZMMU R-10682 1–2 | 50.430058 | 79.896721 | Kazakhstan, 25 km west of Semipalatinsk | ||
| ZMMU R-4293 1–9 | 50.423652 | 80.254928 | Kazakhstan, Semipalatinsk | ||
| ZMMU R-5896 | 40.45 | 54.5667 | Turkmenistan, Tersakkan tract, about 200 km NE from Krasnovodsk | ||
| ZMMU R-9564 1–2 | 43.616389 | 61.926667 | Kazakhstan, Western Kyzyl-Kum, Chaban Kazgan well | ||
| ZMMU R-10472 | 48.144205 | 46.856669 | Russia, Astrakhan region, Big Bogdo mountain | ||
| ZMMU R-9214 | 48.144205 | 46.856669 | Russia, Astrakhan region, Big Bogdo mountain | ||
| ZMMU R-10849 1–2 | 48.206567 | 46.970051 | Russia, Astrakhan region, Bogdinsky-Baskungaksky state reserve, steppe between Surikovskaya and Kordonnaya beams | ||
| ZMMU R-4621 | 48.144205 | 46.856669 | Russia, lower Volga, surroundings of Lake Baskunchak, Mount Bogdo. | ||
| ZMMU R-117 1–3 | 44.759161 | 74.432668 | Kazakhstan, Tai-Aral island in the south of Lake Balkhash. | ||
| ZMMU R-113 1–5 | 48.144205 | 46.856669 | Russia, Kyrgyz steppe (Big Bogdo) | ||
| ZMMU R-12568 | 44.95 | 63.616667 | Kazakhstan | ||
| ZMMU R-12569 | 45.605099 | 59.110612 | Kazakhstan, Aral Sea, Barsakelmes island | ||
| ZMMU R-6835 1–3 | 47.616760 | 80.420510 | Kazakhstan, Semipalatinsk region, right bank of the Ayaguz river, 40 km downstream from the city of Ayaguz | ||
| ZMMU R-4903 1–2 | 48.144205 | 46.856669 | Russia, lower Volga, surroundings of Lake Baskunchak, Mount Bogdo. | ||
| ZMMU R-111 | Expedition to the Persian Gulf | ||||
| ZMMU R-15912 | 43.681 | 108.441 | Mongolia, Dornogob aimag, Ungetiin uul | ||
| ZMMU R-15910 1–2 | 43.608 | 108.237 | Mongolia, Dornogob aimag, southeast of Suykhent | ||
| ZMMU R-15426 1–8 | 42.381003 | 57.370581 | Uzbekistan, Ustyurt plateau, northern shore of Sarykamysh lake | ||
| IZASU 120754—120759 | 42.997157 | 56.080082 | Uzbekistan, Ustyurt | ||
| A. przewalskii | ZMMU R-16103 | - | - | China | |
| ZISP 8271 | 37.354466 | 82.945839 | China, Niyadarya | ||
| ZISP 8250 | 37.110833 | 79.920833 | China, Chotan | ||
| ZISP 8267 | 35.810567 | 83.832288 | China, Kara-Sai | ||
| ZISP 15690 1–3 | 42.112944 | 92.112376 | China, coast from Bugas to Lukchun | ||
| ZISP 8258 1–3 | 41.531399 | 79.496704 | China, Chilan-su | ||
| A. szczerbaki | ZMMU R-12087 | 42.323907 | 59.180661 | Turkmenistan, Tashauz region, Kunya-Urgench | |
| ZISP 18885 7779–7784 | 41.650710 | 59.905444 | Turkmenistan, Takhta village on the Amur Darya | ||
| ZISP 18908 1870–1874 | 41.650710 | 59.905444 | Turkmenistan, Tatauzhskaya region, Takhta village | ||
| ZISP 18519 1–6 | 42.323393 | 59.162816 | Turkmenistan, the vicinity of the village of Kunya-Urgench | ||
| A. tadjikiensis | ZMMU R-16039 | - | - | Turkmenistan | |
| ZISP 18884 1824 | 37.595005 | 68.655830 | Tajikistan, Kurchantyubinsk region, 7 km from the village of Kyzyl-kakha down the Vakhsh river. | ||
| ZISP 18675 | 38.219290 | 69.126832 | Tajikistan, Valley of the Vakhsh River, near Kolkhozabad |
Appendix B
| ZMMU_RAN_032_b_A_loricatus_COI_mtDNA | OR298052 |
| ZMMU_RAN_030_b_A_tokobajevi_COI_mtDNA | OR298053 |
| ZMMU_RAN_155_A_tokobajevi_COI_mtDNA | OR298054 |
| ZMMU_RAN_156_A_tokobajevi_COI_mtDNA | OR298055 |
| ZMMU_RAN_157_A_tokobajevi_COI_mtDNA | OR298056 |
| ZMMU_RAN_160_A_tokobajevi_COI_mtDNA | OR298057 |
| ZMMU_RAN_161_A_tokobajevi_COI_mtDNA | OR298058 |
| ZMMU_R_11384_1_A_tokobajevi_COI_mtDNA | OR298059 |
| ZMMU_R_11384_2_A_tokobajevi_COI_mtDNA | OR298060 |
| ZMMU_R_15426_1_A_pipiens_COI_mtDNA | OR298061 |
| ZMMU_R_15426_5_A_pipiens_COI_mtDNA | OR298062 |
| ZMMU_RAN_4932_laevis_COI_mtDNA | OR298063 |
| ZMMU_R_A_Fergana_COI_mtDNA | OR298064 |
| ZMMU_RAN_4917_emilia_COI_mtDNA | OR298065 |
| ZMMU_RAN_4918_emilia_COI_mtDNA | OR298066 |
| ZMMU_RAN_4924_emilia_COI_mtDNA | OR298067 |
| ZMMU_R_15426_2_A_pipiens_COI_mtDNA | OR298068 |
| ZMMU_R_15426_3_A_pipiens_COI_mtDNA | OR298069 |
| ZMMU_R_15426_4_A_pipiens_COI_mtDNA | OR298070 |
| ZMMU_RAN_2337a_A_przewalskii_COI_mtDNA | OR298071 |
| ZMMU_R_13488_A_pipiens_COI_mtDNA | OR298072 |
| ZMMU_RAN_031_b_A_pipiens_COI_mtDNA | OR298073 |
| ZMMU_R_12914_A_pipiens_COI_mtDNA | OR298074 |
| ZMMU_R_L_36_A_pipiens_COI_mtDNA | OR298075 |
| ZMMU_THD6_A_pipiens_COI_mtDNA | OR298076 |
| ZMMU_R_12915_1_A_pipiens_COI_mtDNA | OR298077 |
| ZMMU_R_12915_2_A_pipiens_COI_mtDNA | OR298078 |
| ZMMU_R_12815_A_pipiens_COI_mtDNA | OR298079 |
| ZMMU_RAN_1426_A_pipiens_COI_mtDNA | OR298080 |
| ZMMU_R_06_Mediodactylus | OR298081 |
| ZMMU_R_160381_laevis_2_COI_mtDNA | OR298082 |
| ZMMU_R_160382_laevis_2_COI_mtDNA | OR298083 |
| ZMMU_RAN_001_laevis_1_COI_mtDNA | OR298084 |
| ZMMU_RAN_002_laevis_1_COI_mtDNA | OR298085 |
| ZMMU_RAN_003_laevis_1_COI_mtDNA | OR298086 |
| ZMMU_R_160_tokobajevi_COI_mtDNA | OR298087 |
| ZMMU_RAN_004_tadjikiensis_COI_mtDNA | OR298088 |
| ZMMU_R_16039_tadjikiensis_COI_mtDNA | OR298089 |
| ZMMU_R_16040_loricatus_COI_mtDNA | OR298090 |
| ZMMU_R_16041_loricatus_COI_mtDNA | OR298091 |
| ZMMU_R_12816_pipiens_COI_mtDNA | OR298092 |
| ZMMU_R_15912_pipiens_COI_mtDNA | OR298093 |
| ZMMU_R_15910_1_pipiens_COI_mtDNA | OR298094 |
| ZMMU_R_15910_2_pipiens_COI_mtDNA | OR298095 |
References
- Sindaco, R.; Jeremcenko, V.K. The Reptiles of the Western Palearctic. In Annotated Checklist and Distributional Atlas of the Turtles, Crocodiles, Amphisbaenians and Lizards of Europe, North Africa, Middle East and Central Asia; Edizioni Belvedere: Latina, Italy, 2008; Volume 1, p. 580. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2022-2. 2022. Available online: https://www.iucnredlist.org (accessed on 2 July 2023).
- Bondarenko, K.A. Transformation of Socio-Cultural Factors Impacting on the external labour migration in Uzbekistan. Spat. Econ. Pros-Transtvennaya Ekon. 2020, 3, 76–108. [Google Scholar] [CrossRef]
- Nazarov, R.A.; Nuridjanov, D.A.; Vashetko, E.V. The current state of endemic reptilian species of the Fergana Valley and possible measures for their conservation. Contemporary Problems of Conservation of Rare, Disappearing Lowland Animals of Uzbekistan; Tashkent, Uzbekistan. 2016, pp. 141–149. Available online: https://istina.msu.ru/publications/article/32600194/ (accessed on 2 July 2023).
- Nazarov, R.A.; Radjabizadeh, M.; Poyarkov, N.A.; Ananjeva, N.B.; Melnikov, D.A.; Pouyani, E.R. A new species of frog-eyed gecko, genus Teratoscincus strauch, 1863 (Squamata: Sauria: Sphaerodactylidae), from central Iran. Russ. J. Herpetol. 2017, 24, 4. [Google Scholar] [CrossRef]
- Bykova, E.A.; Abduraupov, T.V.; Gritsina, M.A.; Esipov, A.V.; Dehkonov, S.I. Current status of rare and endangered species of vertebrates in Fergana region of Uzbekistan. In Actual Problems of Mathematics and Natural Sciences: Materials of the International Scientific-Practical Conference, Petropavlovsk-Barnaul-Surgut-Novosibirsk; Surgut State Pedagogical University: Surgut, Russia, 2022; pp. 257–263. [Google Scholar]
- Pallas, P. Zoographia Russo-Asiatics, Sistens Omnium Animalium in Extenso Imperio Rossico et Adjacentibus Maribus Observatorum Recensionem, Domicilia, Mores et Descriptiones, Anatomen Atque Icones Plurimorum; Caes. Academiae Scientarum: Petropoli, Russia, 1827; Volume 3, p. 426. [Google Scholar]
- Fitzinger, L.J.F.J. Systema Reptilium; Fasciculus Primus; Braumüller et Seidel: Wien, Austria, 1843. [Google Scholar]
- Szczerbak, N.N.; Golubev, M.L. The Gecko Fauna of the USSR and Adjacent Regions; Nauka Dymka: Kiev, Ukraine, 1986; p. 232. [Google Scholar]
- Anderson, S.C. The Lizards of Iran. Society for the Study of Amphibians and Reptiles. In Contributions to Herpetology; Ithaca: New York, NY, USA, 1999; Volume 15, p. 450. [Google Scholar]
- Khan, M.Z.; Mahmood, N. Current population status of diurnal lizards of Karachi, Pakistan. Russ. J. Herpetol. 2003, 10, 241–244. [Google Scholar]
- Bauer, A.M.; Masroor, R.; Titus-Mcquillan, J.; Heinicke, M.P.; Daza, J.D.; Jackman, T.R. A preliminary phylogeny of the Palearctic naked-toed geckos (Reptilia: Squamata: Gekkonidae) with taxonomic implications. Zootaxa 2013, 3599, 301–324. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, I.; Bauer, A.M.; Jackman, T.R.; Karanth, P. Cryptic species and Miocene diversification of palaearctic naked-toed geckos (Squamata: Gekkonidae) in the Indian dry zone. Zool. Scr. 2014, 43, 455–471. [Google Scholar] [CrossRef]
- Gamble, T.; Greenbaum, E.; Jackman, T.R.; Bauer, A.M. Into the light: Diurnality has evolved multiple times in geckos. Biol. J. Linn. Soc. 2015, 115, 896–910. [Google Scholar] [CrossRef]
- Bondarenko, D.A.; Chelintsev, N.G. A comparative estimation of different methods of the line transect census of desert reptiles. Bulletin of Moscow Society of Naturalists. Biol. Ser. 1996, 101, 26–35. [Google Scholar]
- Bondarenko, D.A. “Raspredelenie i plotnost naseleniya sredneaziatskoy cherepahiv Tsentralnyh Kyzylkumah” [The Distribution and Population Density of the Central Asian Tortoise in Central Kizil-Kum (Uzbekistan)]. The Bulletin of Moscow Society of Naturalists. Biol. Ser. 1994, 99, 22–27. [Google Scholar]
- Bondarenko, D.A.; Peregontsev, E.A.; Mukhtar, G.B. Current state of steppe tortoise (Agrionemys horsfieldii Gray, 1844) populations in land-scapes of southern Kazakhstan. Russ. J. Ecol. 2008, 39, 210. [Google Scholar] [CrossRef]
- Bondarenko, D.A.; Peregontsev, E.A. Reptile communities of the Karakalpakian Ustyurt (Uzbekistan). Curr. Stud. Herpetol. 2018, 18, 13–26. [Google Scholar] [CrossRef]
- Hillis, D.M.; Moritz, C.; Mable, B.K. Molecular Systematics; Sinauer: Sunderland, MA, USA, 1996; Volume 2. [Google Scholar]
- Hebert, P.D.; Gregory, T.R. The promise of DNA barcoding for taxonomy. Syst. Biol. 2005, 54, 852–859. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Rodriguez, J.J.; Whitfield, J.B.; Deans, A.R.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D. Extreme diversity of tropical parasitoid wasps exposed by iterative integration of natural history, DNA barcoding, morphology, and collections. Proc. Natl. Acad. Sci. USA 2008, 105, 12359–12364. [Google Scholar] [CrossRef] [PubMed]
- Solovyeva, E.N.; Poyarkov, N.A.; Dunaev, E.A.; Duysebayeva, T.N.; Bannikova, A.A. Molecular differentiation and taxonomy of the sunwatcher toad-headed agama species complex Phrynocephalus superspecies helioscopus (Pallas 1771) (Reptilia: Agamidae). Russ. J. Genet. 2011, 47, 842–856. [Google Scholar] [CrossRef]
- Nazarov, R.A.; Poyarkov, N.A.; Orlov, N.L.; Phung, T.M.; Nguyen, T.T.; Hoang, D.M.; Ziegler, T. Two new cryptic species of the Cyrtodactylus irregularis complex (Squamata: Gekkonidae) from southern Vietnam. Zootaxa 2012, 3302, 1–24. [Google Scholar] [CrossRef]
- Nagy, Z.T.; Sonet, G.; Glaw, F.; Vences, M. First large-scale DNA barcoding assessment of reptiles in the biodiversity hotspot of Madagascar, based on newly designed COI primers. PLoS ONE 2012, 7, e34506. [Google Scholar] [CrossRef]
- Ivanova, N.V.; Zemlak, T.S.; Hanner, R.H.; Hebert, P.D. Universal primer cocktails for fish DNA barcoding. Mol. Ecol. Notes 2007, 7, 544–548. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post–analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.O.; Grismer, L.L. A standardized and statistically defensible framework for quantitative morphological analyses in taxonomic studies. Zootaxa 2021, 5023, 293–300. [Google Scholar] [CrossRef]
- Bogdanov, O.P. Fauna Uzbekskoj SSR. T. 1. Zemnovodnye i Presmykajuschiesya 1. [The Fauna of the Uzbek SSR. T. 1. Amphibians and Rep-Tiles]; Izdatelstvo Akademii Nauk Uzbekskoy SSR [Uzbek SSR Academy Sciences Publishing House]: Tashkent, Uzbekistan, 1960; p. 260. [Google Scholar]
- Lichtenstein, M.H.C. Verzeichniss der Doubletten des zoologischen Museums der Königl. In Universität zu Berlin nebst Beschreibung vieler bisher unbekannter Arten von Säugethieren, Vögeln, Amphibien und Fischen; Königl. Preuss. Akad. Wiss. T. Trautwein: Berlin, Germany, 1823; p. 118. [Google Scholar]
- Lehmann, A. Alexander Lehmann’s Reise nach Buchara und Samarkand in den Jahren 1841 und 1842 (Nach den hinterlas-senen Schriften desselben bearbeitet, und mit Anmerkungen versehen von G. v. Helmersen); Nebst einem zoologischen An-haenge von J.F.Brandt.; Kaiserlichen Akademie der Wissenschaften: Wien, Austria, 1852. [Google Scholar]
- Brandt, J.F. Zoologischer Anhang. In Alexander Lehmann’s Reise Nach Buchara und Samarkand in den Jahren 1841 und 1842; Beitr. z. Kenntn. d. Russ. Reiches; Kaiserlichen Akademie der Wissenschaften: Wien, Austria, 1852; p. 299. [Google Scholar]
- Nikolsky, A.M. Reptiles and amphibians of the Turkestan general-governorship (Herpetologia Turanica). AP Fedchenko, Travel to the Turkestan of the member-founder of the AP Fedchenko Society, accomplished on behalf of the Imperial Society of Amateurs of natural history, anthropology, and ethnography. Zoogeographical Investig. 1899, 23, 21–84. [Google Scholar]
- Nikolsky, A.M. Reptiles and amphibians of Russian Empire (Herpetologica Russica). Mem. De L’academie Imp. Des Sci. De St.-Petersbour 1905, 17, 518. [Google Scholar]
- Nikolsky, A.M. The Fauna of Russia and Adjacent Countries. Reptiles (Reptilia), Vol. 1. Chelonia and Sauria; Academia of Science: Petrograd, Russia, 1915; p. 534. [Google Scholar]
- Nikolsky, A.M. Fauna of Russia and Adjacent Countries. Reptiles (Reptilia) Vol. 2. Ophidia; Academia of Science: Petrograd, Russia, 1916; p. 247. [Google Scholar]
- Nikolsky, A.M. Fauna of Russia and Adjacent Countries. Amphibia; Academia of Science: Petrograd, Russia, 1918; p. 344. [Google Scholar]
- Fedchenko, A.P. Puteshestvie v Turkestan [Travels in Turkestan]. Turkestan Vedemosti 1870, 15, 91–92. [Google Scholar]
- Fedchenko, A.P. Report of the Turkestan scientific expedition of the society (from 1869 to April 15, 1870). Izv. Imp. Obs. Lubit. Estestvozn. Antropol. I Etnogr. 1871, 8, 135–190. [Google Scholar]
- Fedchenko, A.P. Voyage to Turkistan. CH. II. In Kokan Khanate; Izvestia Imperatorskogo O-va Lyubitelei Estestvoznania, Antropologii, i Etnografii: Moscow, Russia, 1875; Volume 7, p. 176. [Google Scholar]
- Strauch, A.A. Bemerkungen über die Geckoniden-Sammlung im zoologischen Museum der Kaiserlichen Akademie der Wissenschaften zu St. Petersburg. Mémoires De L’académie Impériale Des Sci. De St. Pétersbourg Ser. 1887, 7, 1–72. [Google Scholar]
- Vashetko, E.V. Ecology of rapid racerunner in Ferghana Valley. Zool. Zhurnal 1972, 51, 153–155. [Google Scholar]
- Vashetko, E.V.; Kamalova, Z.Y. Turtles, Lizards. In Pozvonochnye Zhivotnye Ferganskoi Doliny [Vertebrate Animals of the Ferghana Valley]; FAN Publ.: Tashkent, Uzbekistan, 1974; pp. 60–74. [Google Scholar]
- Vashetko, E.V.; Kamalova, Z.Y. On the Problem Creating a Reserve for the Conservation Endemic Reptile Species in the Ferghana Valley. In Okhrana Zhivotnogo Mira i Rastenii Uzbekistana: Tezisy Konferentsii [Conservation of the Flora and Fauna of Uzbekistan]; Fan Publ.: Tashkent, Uzbekistan, 1978; pp. 16–17. [Google Scholar]
- Yadgarov, T. Snakes. In Vertebrate Animals of Fergana Valley; FAN: Tashkent, Uzbekistan, 1974; pp. 75–84. [Google Scholar]
- Yadgarov, T.; Allabergenov, K. On conservation of gene pools of the vertebrate animals fauna of the Yazyavan Desert. Uzb. Biol. J. 1990, 2, 59–61. [Google Scholar]
- Tsaruk, O.I. Toad-heads agamas (Phrynocephalus) of Uzbekistan. The state of populations and prospects for conservation. In Reserves of Uzbekistan; Chinor ENK: Tashkent, Uzbekistan, 2001; Issue 3; pp. 72–77. [Google Scholar]
- Chikin, Y.A. Sohranenie Bioraznoobraziya Central’noy Azii, [Conservation of the Biodiversity in the Central Asia]; Zoohobbi: Alma-Ata, Kazakhstan, 1998; Volume 3. [Google Scholar]
- Chikin, Y.A. Protected reptile species from sands of Fergana. In Proceedings of the 1st meeting of the Nikolsky Herpetological Sociality, Puschino, Russia, 4–7 December 2000; pp. 327–328. [Google Scholar]
- Bondarenko, D.A.; Peregontsev, E.A. Distribution of the Central Asian Tortoise Agrionemys horsfieldii (Gray, 1844) in Uzbekistan (range, re-gional and landscape distribution, populations density). Curr. Stud. Herpetol. 2017, 17, 3–4, 124–146. [Google Scholar]
- Nazarov, R.A.; Abduraupov, T.V. New data on distribution and conservation of endemic reptile species in the Fergana Valley. In Proceedings of the Problems of Herpetology: Program and Abstracts of the VIII Congress of the A. M. Nikolsky Herpetological Society (NHS) of the Russian Academy of Sciences «Current Herpetological Research in Eurasia», Moscow, Russia, 3–9 October 2021; pp. 188–189. [Google Scholar]
- Abdulakimovna, M.M.; Narkuziyevich, A.O. Review of literature on Helmintofauna of Rodenta in the territory of the Republic of Uzbekistan. Gospod. I Innow. 2022, 21, 245–249. [Google Scholar]










| Survey Point Number | Alsophylax ferganensis sp. nov. | Alsophylax emilia sp. nov. | Alsophylax loricatus | Teratoscincus rustamowi (Szczerbak, 1979) | Phrynocephalus helioscopus saidalievi (Sattorov, 1981) | Phrynocephalus strauchi | Eremias scripta pherganensis |
|---|---|---|---|---|---|---|---|
| 1 | 5.25 | - | - | - | 0.1 | - | - |
| 2 | - | - | 11 | - | - | - | - |
| 3 | - | - | - | 54 | - | 6.8 | 7.7 |
| 4 | - | - | - | 25.3 | - | 3.2 | 6.8 |
| 5 | - | - | - | 58.5 | - | 9.6 | |
| 6 | - | - | - | 36.3 | - | 2.4 | 2.7 |
| 7 | - | - | - | - | 2.3 | - | - |
| 8 | - | 3.4 | - | - | 3.4 | - | - |
| 9 | - | 4.5 | - | - | 4.2 | - | - |
| 10 | - | - | - | - | 3.7 | - | - |
| 11 | - | - | - | - | 4.1 | 1.6 | - |
| 12 | - | - | - | 14.7 | - | 3 | - |
| Holotype | Paratypes | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ZMMU Re-17532 | UZZI RE–19075 | UZZI RE–19077 | UZZI RE–19076 | ZMMU Re-17533 | UZZI RE–19078 | ZMMU Re-17534 | ZMMU Re-17535 | ZMMU Re-17536 | ZMMU Re-17537 | ZMMU Re-17538 | ZMMU Re-17539 | |
| Field No. | RAN 4358 | RAN 4359 | RAN 4360 | RAN 4361 | RAN 4362 | RAN 4363 | RAN 4364 | RAN 4365 | RAN 4369 | RAN 4370 | RAN 4371 | RAN 4372 |
| SEX | m | f | m | f | f | m | f | f | f | m | m | f |
| SVL | 29.6 | 31.5 | 28.5 | 29.3 | 24.9 | 27.7 | 27.2 | 28.8 | 30.3 | 30 | 25.2 | 29 |
| TL | 36.9 | - | 29.6 | - | - | - | - | 28.7 | 32.9 | 37.7 | 32.1 | 23.7 |
| HL | 8.1 | 7.9 | 8 | 7.8 | 8 | 7.4 | 7.6 | 7.2 | 7.8 | 7.9 | 7 | 7.9 |
| HW | 4.6 | 4 | 4.2 | 3.9 | 3.5 | 4 | 4 | 3.8 | 4 | 3.9 | 3.5 | 3.7 |
| HH | 2.7 | 2.8 | 3 | 2.5 | 2.5 | 2.3 | 2.5 | 2.6 | 2.4 | 2.5 | 2 | 2 |
| SnEye | 3 | 2.7 | 2.7 | 2.6 | 2.3 | 2.7 | 2.5 | 2.4 | 2.5 | 2.5 | 2.4 | 2.4 |
| EyeEar | 2.7 | 2.2 | 2.4 | 2.6 | 2.1 | 2.1 | 2.4 | 2.2 | 2.4 | 2.5 | 2.1 | 2.2 |
| Orb D | 2 | 2 | 1.8 | 1.9 | 1.6 | 1.6 | 1.8 | 1.8 | 1.8 | 1.9 | 1.7 | 1.9 |
| LS | 4.7 | 4.3 | 4.6 | 4.5 | 4.4 | 4.4 | 4.6 | 4.6 | 4.7 | 4.3 | 4.2 | 4.5 |
| ForeaL | 4.3 | 4 | 4 | 3.8 | 3.5 | 3.8 | 3.9 | 4 | 4.1 | 3.8 | 3.5 | 3.9 |
| FemurL | 5.9 | 5.3 | 5.9 | 5.8 | 5.4 | 5.5 | 5.3 | 5.4 | 5.7 | 5.3 | 5.4 | 5.3 |
| CrusL | 5.9 | 5.6 | 5.6 | 5.5 | 4.9 | 5.1 | 5.2 | 5.1 | 6 | 5 | 5 | 5.6 |
| LD4A | 2.3 | 2.1 | 1.8 | 2.1 | 2.3 | 2.4 | 2.2 | 2.3 | 1.9 | 2.5 | 2.1 | 2.3 |
| LD4P | 3.1 | 3 | 2.7 | 3.1 | 3.2 | 2.9 | 3.1 | 3 | 2.8 | 3.3 | 2.9 | 3.3 |
| SL (r/l) | 7/7 | 7/7 | 7/7 | 7/7 | 7/7 | 7/7 | 7/7 | 7/8 | 7/8 | 7/7 | 8/6 | 7/7 |
| IL (r/l) | 5/5 | 5/6 | 5/5 | 5/5 | 5/5 | 5/5 | 5/5 | 6/6 | 5/6 | 5/5 | 6/5 | 5/5 |
| SEH | 18 | 18 | 22 | 21 | 20 | 19 | 18 | 20 | 19 | 19 | 20 | 19 |
| LF4 | 14 | 14 | 14 | 15 | 15 | 13 | 13 | 15 | 13 | 13 | 12 | 14 |
| LT4 | 18 | 15 | 16 | 15 | 14 | 15 | 15 | 17 | 16 | 14 | 15 | 18 |
| SLB | 86 | 87 | 86 | 83 | 89 | 88 | 89 | 97 | 85 | 84 | 94 | 94 |
| PP | 7 | - | 6 | 7 | - | 6 | 7 | 8 | 7 | 6 | 7 | 7 |
| Spurs (r/l) | 2/2 | 3/3 | 3/2 | 2/3 | 3/3 | 3/3 | 2/2 | 3/3 | 3/3 | 3/3 | 2/2 | 2/2 |
| Sq | 44 | 48 | 50 | 49 | 44 | 45 | 47 | 47 | 47 | 48 | 47 | 51 |
| Alsophylax cf. laevis | Alsophylax laevis | Alsophylax loricatus | Alsophylax pipiens | Alsophylax cf. pipiens | Alsophylax przewalskii | Alsophylax szczerbaki | Alsophylax tadjikiensis | Alsophylax ferganensis | |
|---|---|---|---|---|---|---|---|---|---|
| N | 21 | 60 | 25 | 46 | 107 | 19 | 17 | 3 | 12 |
| SVL | |||||||||
| mean | 31.97 | 32.55 | 30.74 | 33.14 | 32.00 | 30.35 | 28.26 | 27.10 | 28.50 |
| ±SD | 1.65 | 2.34 | 3.02 | 3.06 | 2.47 | 1.58 | 1.27 | 2.07 | 1.50 |
| range | 27.2–35.2 | 26.6–38.7 | 24.6–40.9 | 27.4–41.5 | 26.2–39.8 | 26.59–33.5 | 26.3–32.4 | 24–29.1 | 24.9–31.5 |
| TL | |||||||||
| mean | 31.10 | 33.96 | 35.91 | 33.68 | 32.57 | 38.18 | 33.31 | 25.30 | 30.43 |
| ±SDSD | 4.13 | 2.74 | 3.60 | 2.69 | 3.53 | 3.81 | 3.56 | 0.00 | 2.98 |
| range | 24–42.6 | 25–42 | 27.5–44.6 | 23.3–43.5 | 19–42.8 | 31.19–45.3 | 23.7–42.3 | 25.30 | 23.7–37.7 |
| HL | |||||||||
| mean | 8.86 | 8.87 | 8.32 | 9.07 | 8.89 | 8.37 | 7.98 | 7.53 | 7.72 |
| ±SDSD | 0.44 | 0.55 | 0.91 | 0.79 | 0.63 | 0.32 | 0.30 | 0.49 | 0.28 |
| range | 7.9–9.7 | 7.4–10.3 | 7–10.7 | 7.5–10.9 | 7.2–11 | 7.02–8.93 | 7.3–8.8 | 6.8–8 | 7–8.1 |
| HW | |||||||||
| mean | 5.02 | 4.94 | 4.44 | 5.12 | 5.14 | 4.82 | 4.24 | 3.90 | 3.93 |
| ±SD | 0.32 | 0.36 | 0.48 | 0.52 | 0.45 | 0.38 | 0.20 | 0.13 | 0.21 |
| range | 4.4–5.7 | 3.9–5.9 | 3.6–5.7 | 3.6–6.7 | 4–6.5 | 4.2–5.8 | 3.7–4.7 | 3.7–3.9 | 3.5–4.6 |
| HH | |||||||||
| mean | 3.18 | 3.06 | 2.70 | 3.31 | 3.32 | 3.36 | 2.69 | 2.50 | 2.48 |
| ±SD | 0.32 | 0.28 | 0.32 | 0.35 | 0.36 | 0.41 | 0.16 | 0.13 | 0.21 |
| range | 2.3–4.2 | 2.4–4.2 | 2–3.51 | 2.2–4.1 | 2.2–4.5 | 2.5–4.33 | 2.2–3 | 2.7–2.4 | 2–3 |
| SnEye | |||||||||
| mean | 2.92 | 2.86 | 2.80 | 2.98 | 2.91 | 2.74 | 2.52 | 2.43 | 2.56 |
| ±SD | 0.22 | 0.18 | 0.30 | 0.30 | 0.22 | 0.15 | 0.09 | 0.09 | 0.15 |
| range | 2.5–3.4 | 2.3–3.4 | 2.3–3.6 | 2.4–3.8 | 2.4–3.6 | 2.19–3 | 2.3–2.7 | 2.3–2.5 | 2.3–3 |
| Orb D | |||||||||
| mean | 2.18 | 2.05 | 1.82 | 2.15 | 2.15 | 1.72 | 1.74 | 1.80 | 1.82 |
| ±SD | 0.15 | 0.11 | 0.29 | 0.18 | 0.17 | 0.23 | 0.08 | 0.20 | 0.10 |
| range | 1.9–2.6 | 1.6–2.5 | 1.26–2.7 | 1.69–2.8 | 1.6–2.7 | 1.18–2 | 1.5–1.9 | 1.5–2.1 | 1.6–2 |
| LS | |||||||||
| mean | 5.11 | 4.70 | 4.40 | 4.93 | 4.72 | 4.39 | 3.83 | 3.93 | 4.48 |
| ±SD | 0.40 | 0.31 | 0.70 | 0.53 | 0.40 | 0.33 | 0.13 | 0.16 | 0.14 |
| range | 4.2–6.7 | 3.6–5.3 | 3.5–6 | 3.8–6.2 | 3.6–6.4 | 3.52–5 | 3.5–4.2 | 3.7–4 | 4.2–4.7 |
| ForeaL | |||||||||
| mean | 4.40 | 3.85 | 3.49 | 4.15 | 4.00 | 3.51 | 3.15 | 3.37 | 3.88 |
| ±SD | 0.29 | 0.26 | 0.59 | 0.45 | 0.31 | 0.27 | 0.14 | 0.18 | 0.17 |
| range | 3.8–5.6 | 3–4.7 | 2.7–5 | 3–5.5 | 2.9–5 | 3.1–4.11 | 3–3.4 | 3.1–3.4 | 3.5–4.3 |
| FemurL | |||||||||
| mean | 5.89 | 5.29 | 5.19 | 5.91 | 5.74 | 5.17 | 4.45 | 4.53 | 5.52 |
| ±SD | 0.35 | 0.32 | 0.74 | 0.65 | 0.44 | 0.29 | 0.22 | 0.11 | 0.21 |
| range | 5.2–6.6 | 4.2–6.1 | 4–7.2 | 4.4–7.9 | 4.5–7.3 | 4.5–5.71 | 4.1–5.2 | 4.7–4.4 | 5.3–5.9 |
| CrusL | |||||||||
| mean | 5.70 | 5.22 | 4.73 | 5.60 | 5.27 | 4.84 | 4.31 | 4.50 | 5.38 |
| ±SD | 0.34 | 0.36 | 0.80 | 0.57 | 0.48 | 0.34 | 0.18 | 0.07 | 0.33 |
| range | 5–6.5 | 4–6 | 3.6–6.8 | 4.3–7.1 | 3.9–6.7 | 4.1–5.49 | 3.9–4.7 | 4.4–4.5 | 4.9–6 |
| LD4A | |||||||||
| mean | 2.20 | 2.21 | 2.52 | 2.76 | 2.81 | 2.50 | 2.26 | 1.90 | 2.19 |
| ±SD | 0.17 | 0.17 | 0.40 | 0.31 | 0.26 | 0.18 | 0.21 | 0.07 | 0.16 |
| range | 1.9–2.6 | 1.8–2.7 | 1.8–3.6 | 2.1–3.6 | 2–3.5 | 2.1–2.9 | 1.5–2.8 | 1.9–1.8 | 1.8–2.5 |
| LD4P | |||||||||
| mean | 2.97 | 2.92 | 3.45 | 3.67 | 3.58 | 3.65 | 3.19 | 2.33 | 3.03 |
| ±SD | 0.24 | 0.19 | 0.39 | 0.38 | 0.33 | 0.25 | 0.16 | 0.18 | 0.15 |
| range | 2.2–3.7 | 2.3–3.4 | 2.1–4.4 | 2.7–4.41 | 2.7–4.4 | 2.75–4.17 | 2.9–3.6 | 2.2–2.2 | 2.7–3.3 |
| EyeEar | |||||||||
| mean | 2.61 | 2.49 | 2.46 | 2.70 | 2.65 | 2.48 | 2.55 | 2.47 | 2.33 |
| ±SD | 0.23 | 0.21 | 0.23 | 0.28 | 0.25 | 0.17 | 0.14 | 0.11 | 0.18 |
| range | 2–3 | 2–3 | 2–3 | 2.1–3.4 | 1.9–3.6 | 2.2–2.89 | 2.3–2.8 | 2.3–2.5 | 2.1–2.7 |
| Holotype | Paratypes | ||
|---|---|---|---|
| ZMMU Re-17544 | UZZI RE–19079 | ZMMU Re-17545 | |
| Field No. | RAN 4917 | RAN 4918 | RAN 4924 |
| SEX | m | m | m |
| SVL | 35 | 32.6 | 31.5 |
| TL | 40.2 | 15.3 regenerated | 33.1 |
| HL | 9.7 | 9.2 | 9 |
| HW | 5.8 | 5.2 | 5.1 |
| HH | 2.7 | 2.8 | 2.6 |
| SnEye | 3 | 2.8 | 3 |
| EyeEar | 2.9 | 2.8 | 2.6 |
| Orb D | 2.1 | 2.2 | 2.2 |
| LS | 5.8 | 5.4 | 5.2 |
| ForeaL | 5.1 | 4.7 | 4.6 |
| FemurL | 6.5 | 6.4 | 5.3 |
| CrusL | 6.5 | 6.0 | 5.7 |
| LD4A | 2.5 | 2.2 | 2.2 |
| LD4P | 3 | 2.6 | 2.6 |
| SL (r/l) | 8/8 | 7/7 | 8/9 |
| IL (r/l) | 6/7 | 6/6 | 7/6 |
| SEH | 22 | 22 | 22 |
| LF4 | 11 | 12 | 11 |
| LT4 | 13 | 14 | 14 |
| SLB | 97 | 102 | 104 |
| PP | 8 | 9 | 7 |
| Spurs (r/l) | 2/2 | 2/3 | 2/2 |
| Sq | 65 | 64 | 59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarov, R.A.; Abduraupov, T.V.; Shepelya, E.Y.; Gritsina, M.A.; Melnikov, D.A.; Buehler, M.D.; Lapin, J.D.; Poyarkov, N.A.; Grismer, J.L. The Fergana Valley Is an Isolate of Biodiversity: A Discussion of the Endemic Herpetofauna and Description of Two New Species of Alsophylax (Sauria: Gekkonidae) from Eastern Uzbekistan. Animals 2023, 13, 2516. https://doi.org/10.3390/ani13152516
Nazarov RA, Abduraupov TV, Shepelya EY, Gritsina MA, Melnikov DA, Buehler MD, Lapin JD, Poyarkov NA, Grismer JL. The Fergana Valley Is an Isolate of Biodiversity: A Discussion of the Endemic Herpetofauna and Description of Two New Species of Alsophylax (Sauria: Gekkonidae) from Eastern Uzbekistan. Animals. 2023; 13(15):2516. https://doi.org/10.3390/ani13152516
Chicago/Turabian StyleNazarov, Roman A., Timur V. Abduraupov, Evgeniya Yu. Shepelya, Mariya A. Gritsina, Daniel A. Melnikov, Matthew D. Buehler, Jack D. Lapin, Nikolay A. Poyarkov, and Jesse L. Grismer. 2023. "The Fergana Valley Is an Isolate of Biodiversity: A Discussion of the Endemic Herpetofauna and Description of Two New Species of Alsophylax (Sauria: Gekkonidae) from Eastern Uzbekistan" Animals 13, no. 15: 2516. https://doi.org/10.3390/ani13152516