
Sex-Linked Loci on the W Chromosome in the Multi-Ocellated Racerunner (Eremias multiocellata) Confirm Genetic Sex-Determination Stability in Lacertid Lizards


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
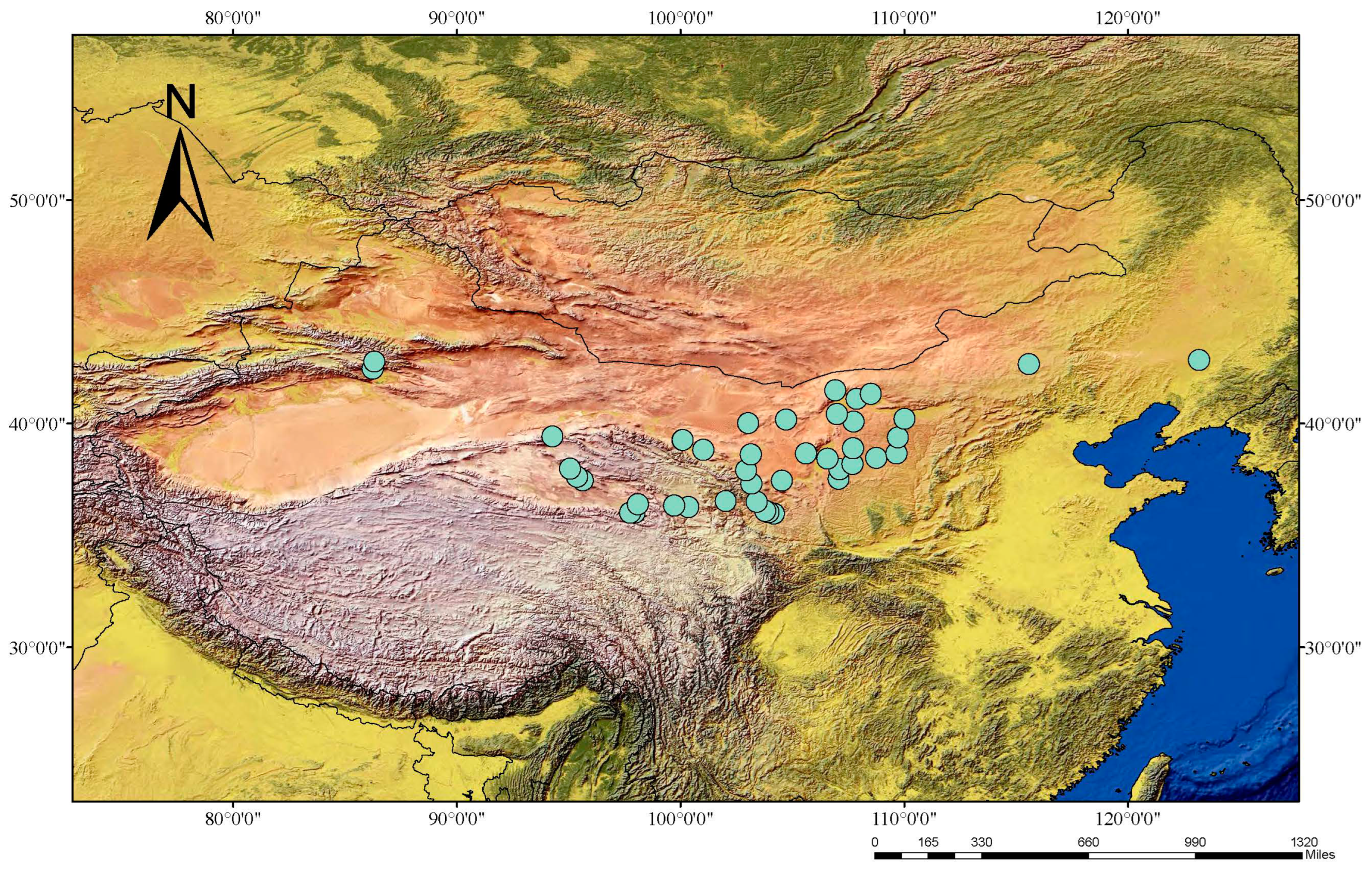
2.1. Sampling and Preparation
2.2. DNA Extraction and Genotyping by Sequencing
2.3. Filtering and SNP Calling
2.4. Screening Sex-Linked Markers
2.5. Confirmation and Assignment of Sex-Specific Markers
2.6. PCR Validation
2.7. Sex-Linked Markers Assignment
3. Results
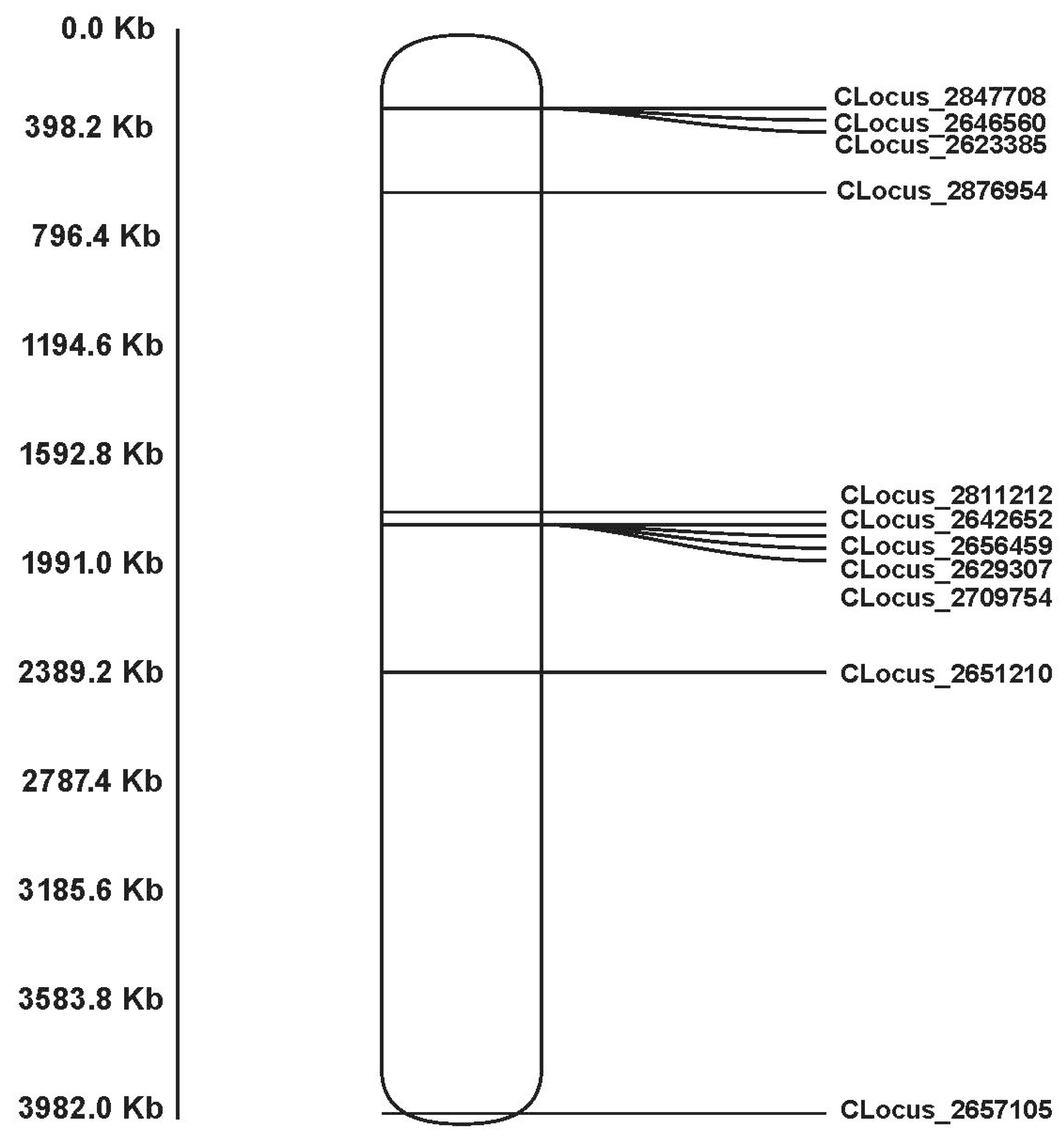
3.1. GBS Data Analyses and Sex-Linked Loci Screening
3.2. Sex-Linked Marker Screening
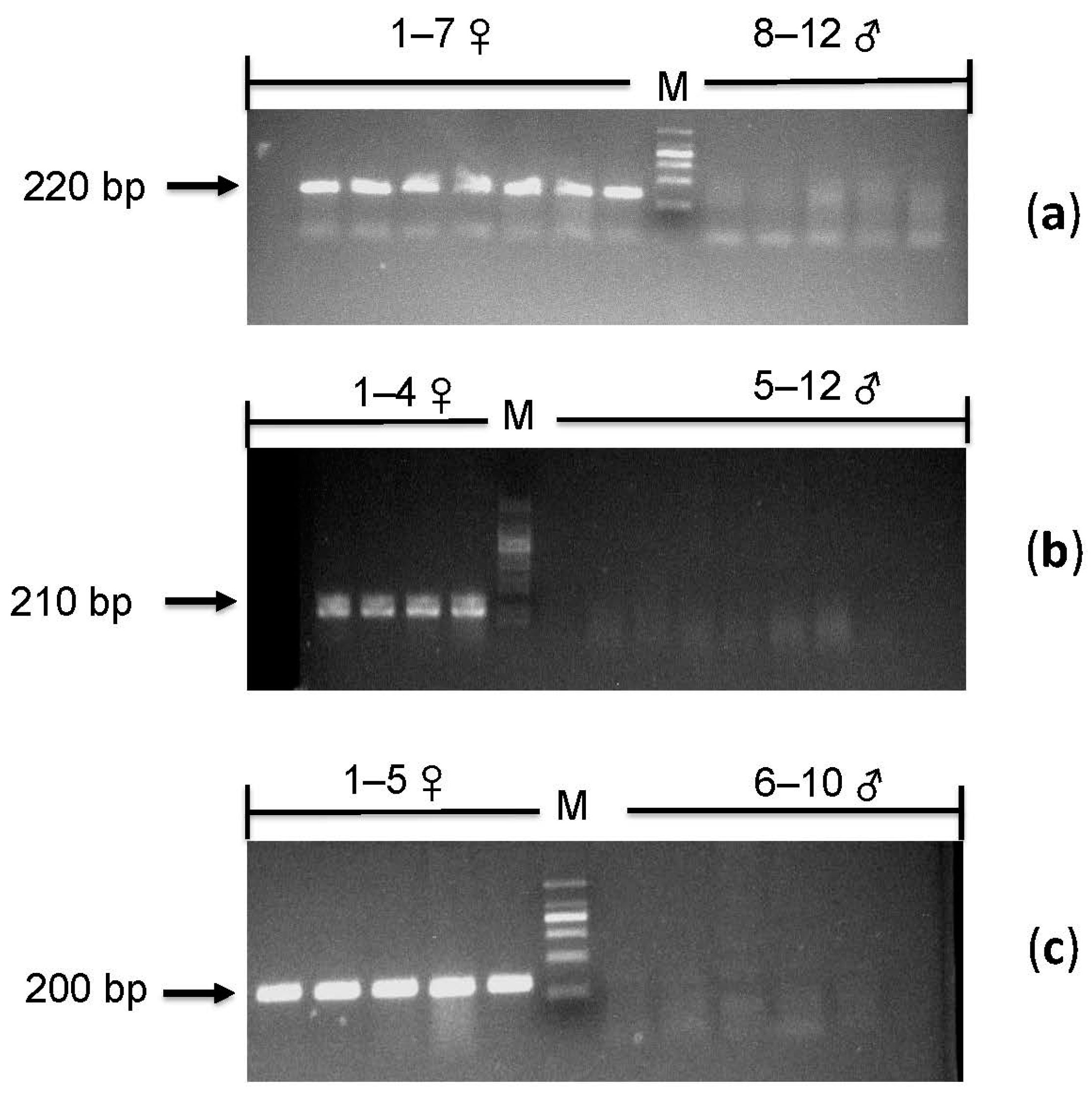
3.3. Validation of the Sex-Linked Marker
3.4. PCR Verification
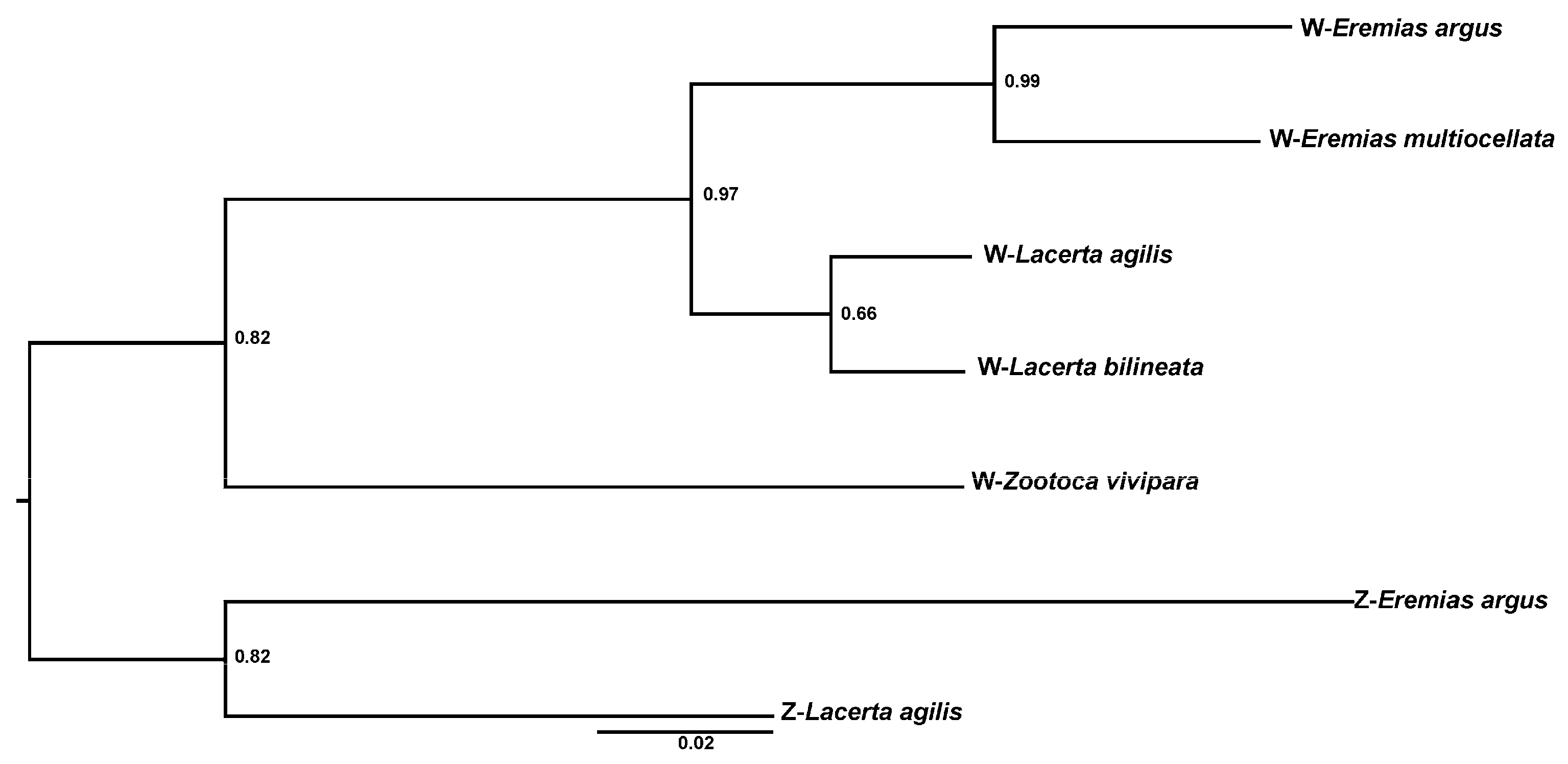
3.5. Sex-Linked Markers Assignment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Craves, J.A.M. Weird animal genomes and the evolution of vertebrate sex and sex chromosomes. Annu. Rev. Genet. 2008, 42, 565–586. [Google Scholar] [CrossRef]
- Sarre, S.D.; Ezaz, T.; Georges, A. Transitions between sex-determining systems in reptiles and amphibians. Annu. Rev. Genom. Hum. Genet. 2011, 12, 391–406. [Google Scholar] [CrossRef] [Green Version]
- Pokorna, M.; Kratochvil, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J. Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Ezaz, T.; Stiglec, R.; Veyrunes, F.; Marshall Graves, J.A. Relationships between vertebrate ZW and XY sex chromosome systems. Curr. Biol. 2006, 16, R736–R743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altmanova, M.; Rovatsos, M.; Pokorna, M.J.; Vesely, M.; Wagner, F.; Kratochvil, L. All iguana families with the exception of basilisks share sex chromosomes. Zoology 2018, 126, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.M.I.; Sarre, S.D.; Gleeson, D.; Georges, A.; Ezaz, T. Did lizards follow unique pathways in sex chromosome evolution? Genes 2018, 9, 239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiggins, J.M.; Santoyo-Brito, E.; Scales, J.B.; Fox, S.F. Gene dose indicates presence of sex chromosomes in collared lizards (Crotaphytus collaris), a species with temperature-influenced sex determination. Herpetologica 2020, 76, 27–30. [Google Scholar] [CrossRef]
- Tang, X.L.; Yue, F.; Yan, X.F.; Zhang, D.J.; Xin, Y.; Wang, C.; Chen, Q. Effects of gestation temperature on offspring sex and maternal reproduction in a viviparous lizard (Eremias multiocellata) living at high altitude. J. Therm. Biol. 2012, 37, 438–444. [Google Scholar] [CrossRef]
- Hill, P.L.; Burridge, C.P.; Ezaz, T.; Wapstra, E. Conservation of sex-linked markers among conspecific populations of a viviparous skink, Niveoscincus ocellatus, exhibiting genetic and temperature-dependent sex determination. Genome Biol. Evol. 2018, 10, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Shine, R.; Warner, D.A.; Radder, R. Windows of embryonic sexual lability in two lizard species with environmental sex determination. Ecology 2007, 88, 1781–1788. [Google Scholar] [CrossRef]
- Quinn, A.E.; Sarre, S.D.; Ezaz, T.; Graves, J.A.M.; Georges, A. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol. Lett. 2011, 7, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.M.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Quinn, A.E.; Sarre, S.D.; O’Meally, D.; Georges, A.; Graves, J.A.M. Molecular marker suggests rapid changes of sex-determining mechanisms in Australian dragon lizards. Chromosome Res. 2009, 17, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Mezzasalma, M.; Guarino, F.M.; Odierna, G. Lizards as model organisms of sex chromosome evolution: What we really know from a systematic distribution of available data? Genes 2021, 12, 1341. [Google Scholar] [CrossRef]
- Burbrink, F.T.; Grazziotin, F.G.; Pyron, R.A.; Cundall, D.; Donnellan, S.; Irish, F.; Keogh, J.S.; Kraus, F.; Murphy, R.W.; Noonan, B.; et al. Interrogating genomic-scale data for squamata (Lizards, Snakes, and Amphisbaenians) shows no support for key traditional morphological relationships. Syst. Biol. 2020, 69, 502–520. [Google Scholar] [CrossRef]
- Pen, I.; Uller, T.; Feldmeyer, B.; Harts, A.; While, G.M.; Wapstra, E. Climate-driven population divergence in sex-determining systems. Nature 2010, 468, 436–438. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukic, J.; Mrugala, A.; Suwala, G.; Lymberakis, P.; Kratochvil, L. Little evidence for switches to environmental sex determination and turnover of sex chromosomes in lacertid lizards. Sci. Rep. 2019, 9, 7832. [Google Scholar] [CrossRef] [Green Version]
- Rovatsos, M.; Vukic, J.; Altmanova, M.; Pokorna, M.J.; Moravec, J.; Kratochvil, L. Conservation of sex chromosomes in lacertid lizards. Mol. Ecol. 2016, 25, 3120–3126. [Google Scholar] [CrossRef]
- Pokorna, M.; Kratochvil, L.; Kejnovsky, E. Microsatellite distribution on sex chromosomes at different stages of heteromorphism and heterochromatinization in two lizard species (Squamata: Eublepharidae: Coleonyx elegans and Lacertidae: Eremias velox). BMC Genet. 2011, 12, 90. [Google Scholar] [CrossRef] [Green Version]
- Orlova, V.F.; Poyarkov, N.A., Jr.; Chirikova, M.A.; Nazarov, R.A.; Munkhbaatar, M.; Munkhbayar, K.; Terbish, K. MtDNA differentiation and taxonomy of Central Asian racerunners of Eremias multiocellata-E. przewalskii species complex (Squamata, Lacertidae). Zootaxa 2017, 4282, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Zeng, X.M.; Chen, B.; Wang, Y.Z. The research on the karyotypes of six species in the genus Eremias from China. Yi Chuan 2004, 26, 669–675. [Google Scholar]
- Wang, C.; Tang, X.; Xin, Y.; Yue, F.; Yan, X.; Liu, B.; An, B.; Wang, X.; Chen, Q. Identification of sex chromosomes by means of comparative genomic hybridization in a lizard, Eremias multiocellata. Zoolog. Sci. 2015, 32, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.J.; Tang, X.L.; Yue, F.; Chen, Z.; Li, R.D.; Chen, Q. Effect of gestation temperature on sexual and morphological phenotypes of offspring in a viviparous lizard, Eremias multiocellata. J. Therm. Biol. 2010, 35, 129–133. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukic, J.; Kratochvil, L. Mammalian X homolog acts as sex chromosome in lacertid lizards. Heredity 2016, 117, 8–13. [Google Scholar] [CrossRef]
- Charlesworth, D. The timing of genetic degeneration of sex chromosomes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2021, 376, 20200093. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamble, T.; Zarkower, D. Identification of sex-specific molecular markers using restriction site associated DNA sequencing (RAD-seq). Mol. Ecol. Resour. 2014, 14, 902–913. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Xia, Y.; Yue, B.; Zeng, X. Assigning the sex-specific markers via genotyping-by-sequencing onto the Y chromosome for a torrent frog Amolops mantzorum. Genes 2020, 11, 727. [Google Scholar] [CrossRef]
- Jeffries, D.L.; Lavanchy, G.; Sermier, R.; Sredl, M.J.; Miura, I.; Borzee, A.; Barrow, L.N.; Canestrelli, D.; Crochet, P.-A.; Dufresnes, C.; et al. A rapid rate of sex-chromosome turnover and non-random transitions in true frogs. Nat. Commun. 2018, 9, 4088. [Google Scholar] [CrossRef] [Green Version]
- Brelsford, A.; Lavanchy, G.; Sermier, R.; Rausch, A.; Perrin, N. Identifying homomorphic sex chromosomes from wild-caught adults with limited genomic resources. Mol. Ecol. Resour. 2017, 17, 752–759. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Luo, W.; Xia, Y.; Zeng, X. Using sex-linked markers via genotyping-by-sequencing to identify XX/XY Sex chromosomes in the spiny frog (Quasipaa boulengeri). Genes 2022, 13, 575. [Google Scholar] [CrossRef]
- Myers, E.A.; Mulcahy, D.G.; Falk, B.; Johnson, K.; Carbi, M.; de Queiroz, K. Interspecific gene flow and mitochondrial genome capture during the radiation of Jamaican Anolis lizards (Squamata; Iguanidae). Syst. Biol. 2022, 71, 501–511. [Google Scholar] [CrossRef]
- Finger, N.; Farleigh, K.; Bracken, J.T.; Leache, A.D.; Francois, O.; Yang, Z.; Flouri, T.; Charran, T.; Jezkova, T.; Williams, D.A.; et al. Genome-scale data reveal deep lineage divergence and a complex demographic history in the Texas horned lizard (Phrynosoma cornutum) throughout the southwestern and central United States. Genome Biol. Evol. 2022, 14, evab260. [Google Scholar] [CrossRef] [PubMed]
- Harolow, P. A harmless technique for sexing hatchiling lizards. Herpetol. Rev. 1996, 27, 71–72. [Google Scholar]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Catchen, J.M.; Amores, A.; Hohenlohe, P.; Cresko, W.; Postlethwait, J.H. Stacks: Building and genotyping loci de novo from short-read sequences. G3 2011, 1, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlesworth, D.; Mank, J.E. The birds and the bees and the flowers and the trees: Lessons from genetic mapping of sex determination in plants and animals. Genetics 2010, 186, 9–31. [Google Scholar] [CrossRef] [Green Version]
- Ping, J.; Xia, Y.; Ran, J.; Zeng, X. Heterogeneous evolution of sex chromosomes in the torrent frog genus Amolops. Int. J. Mol. Sci. 2022, 23, 11146. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Du, J.; Yang, L.; Liang, Q.; Yang, M.; Zhou, X.; Du, W. Chromosome-level genome assembly and population genomics of Mongolian racerunner (Eremias argus) provide insights into high-altitude adaptation in lizards. BMC Biol. 2023, 21, 40. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Holleley, C.E.; Sarre, S.D.; O’Meally, D.; Georges, A. Sex reversal in reptiles: Reproductive oddity or powerful driver of evolutionary change? Sex. Dev. 2016, 10, 279–287. [Google Scholar] [CrossRef]
- Nielsen, S.V.; Guzman-Mendez, I.A.; Gamble, T.; Blumer, M.; Pinto, B.J.; Kratochvil, L.; Rovatsos, M. Escaping the evolutionary trap? Sex chromosome turnover in basilisks and related lizards (Corytophanidae: Squamata). Biol. Lett. 2019, 15, 20190498. [Google Scholar] [CrossRef]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction Site-Associated DNA Sequencing (RAD-seq) Reveals an Extraordinary Number of Transitions among Gecko Sex-Determining Systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [Green Version]
- Keating, S.E.; Blumer, M.; Grismer, L.L.; Lin, A.; Nielsen, S.V.; Thura, M.K.; Wood, P.L.; Quah, E.S.H.; Gamble, T. Sex chromosome turnover in bent-toed geckos (Cyrtodactylus). Genes 2021, 12, 116. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Sequence (5′–3′) | Annealing Temperature (°C) | Length (bp) |
|---|---|---|---|
| 2648560 | Forward: CGAAACATTTGACTTCTGACGC | 55 | 298 |
| Reverse: TCCGACCTTATATGCCTCTTCT | |||
| 2847708 | Forward: AGCTCTCCCCAATCATCACC | 55 | 298 |
| Reverse: AGAAAGCAGGAAGGGGAGTG | |||
| 2811212 | Forward: GTCCAACATCCTTTGCCCAA | 55 | 298 |
| Reverse: CACCATCAACCTCCTCCCTT | |||
| 2811108 | Forward: TCTTCTGATGGCCTTGGACC | 55 | 298 |
| Reverse: GGCATCCAAACATCTCACCA | |||
| 2645069 | Forward: AGCATCTTCTTCCCTCTCTTCT | 55 | 298 |
| Reverse: TGGGATCTGGATTGGCTGTT | |||
| 2642652 | Forward: AGGATGTGGGAGGCTGTTTT | 55 | 298 |
| Reverse: TGCAACTGTGGACTGAAACG | |||
| 2628103 | Forward: AGCCTCTTCTTCCACTTCCA | 55 | 298 |
| Reverse: AGTGTGTGATGGAGATGGGG | |||
| 2629307 | Forward: TGCCTCCCATCACAGTTCTT | 55 | 298 |
| Reverse: TACAGCAGGACAAAAGGGCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chu, Z.; Wang, Z.; Zheng, Y.; Xia, Y.; Guo, X. Sex-Linked Loci on the W Chromosome in the Multi-Ocellated Racerunner (Eremias multiocellata) Confirm Genetic Sex-Determination Stability in Lacertid Lizards. Animals 2023, 13, 2180. https://doi.org/10.3390/ani13132180
Chu Z, Wang Z, Zheng Y, Xia Y, Guo X. Sex-Linked Loci on the W Chromosome in the Multi-Ocellated Racerunner (Eremias multiocellata) Confirm Genetic Sex-Determination Stability in Lacertid Lizards. Animals. 2023; 13(13):2180. https://doi.org/10.3390/ani13132180
Chicago/Turabian StyleChu, Zhangqing, Ziwen Wang, Yuchi Zheng, Yun Xia, and Xianguang Guo. 2023. "Sex-Linked Loci on the W Chromosome in the Multi-Ocellated Racerunner (Eremias multiocellata) Confirm Genetic Sex-Determination Stability in Lacertid Lizards" Animals 13, no. 13: 2180. https://doi.org/10.3390/ani13132180