
Intermittent and Mild Cold Stimulation Maintains Immune Function Stability through Increasing the Levels of Intestinal Barrier Genes of Broilers

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection
2.3. Evaluation of Broiler Production Performance
2.4. RNA Extraction and Reverse Transcription
2.5. Quantitative Real-Time PCR (qRT-PCR) Analysis
2.6. Western Blot Analysis
2.7. Elisa Detection
2.8. Statistics and Analysis
3. Results
3.1. Production Performance of Broilers
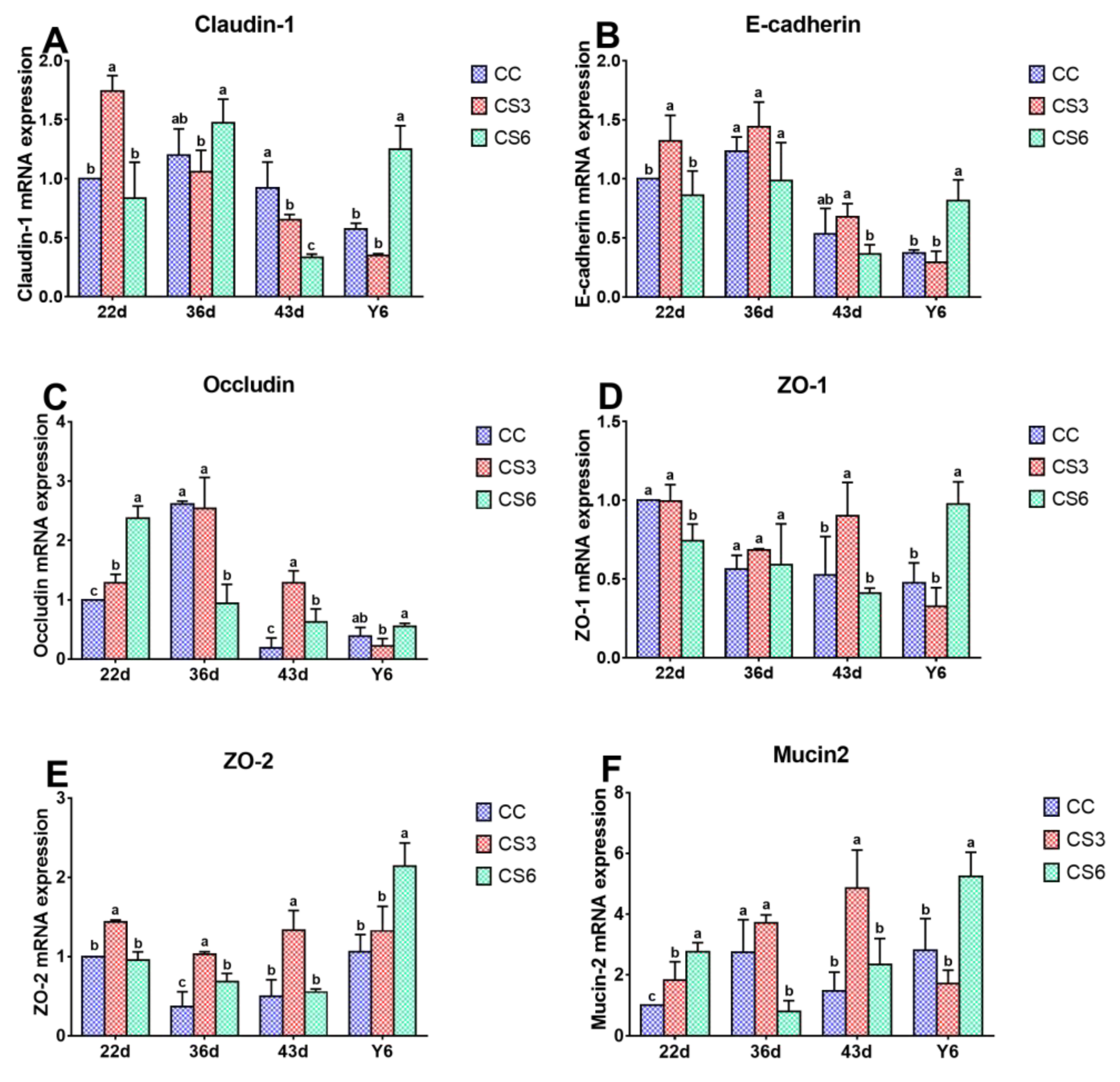
3.2. mRNA Levels of Intestinal Barrier Genes in the Duodenum
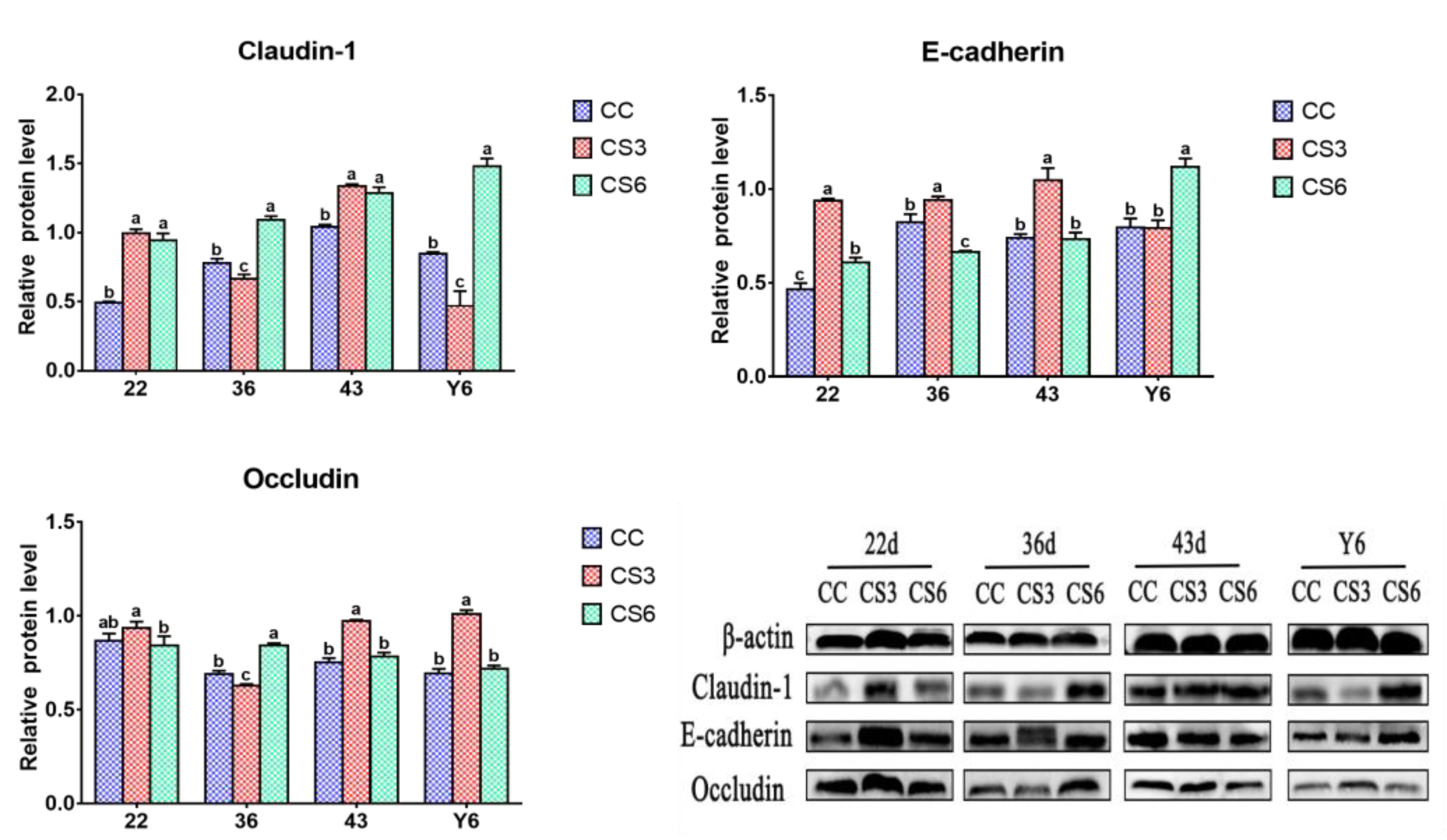
3.3. Protein Levels of Intestinal Barrier Genes in the Duodenum
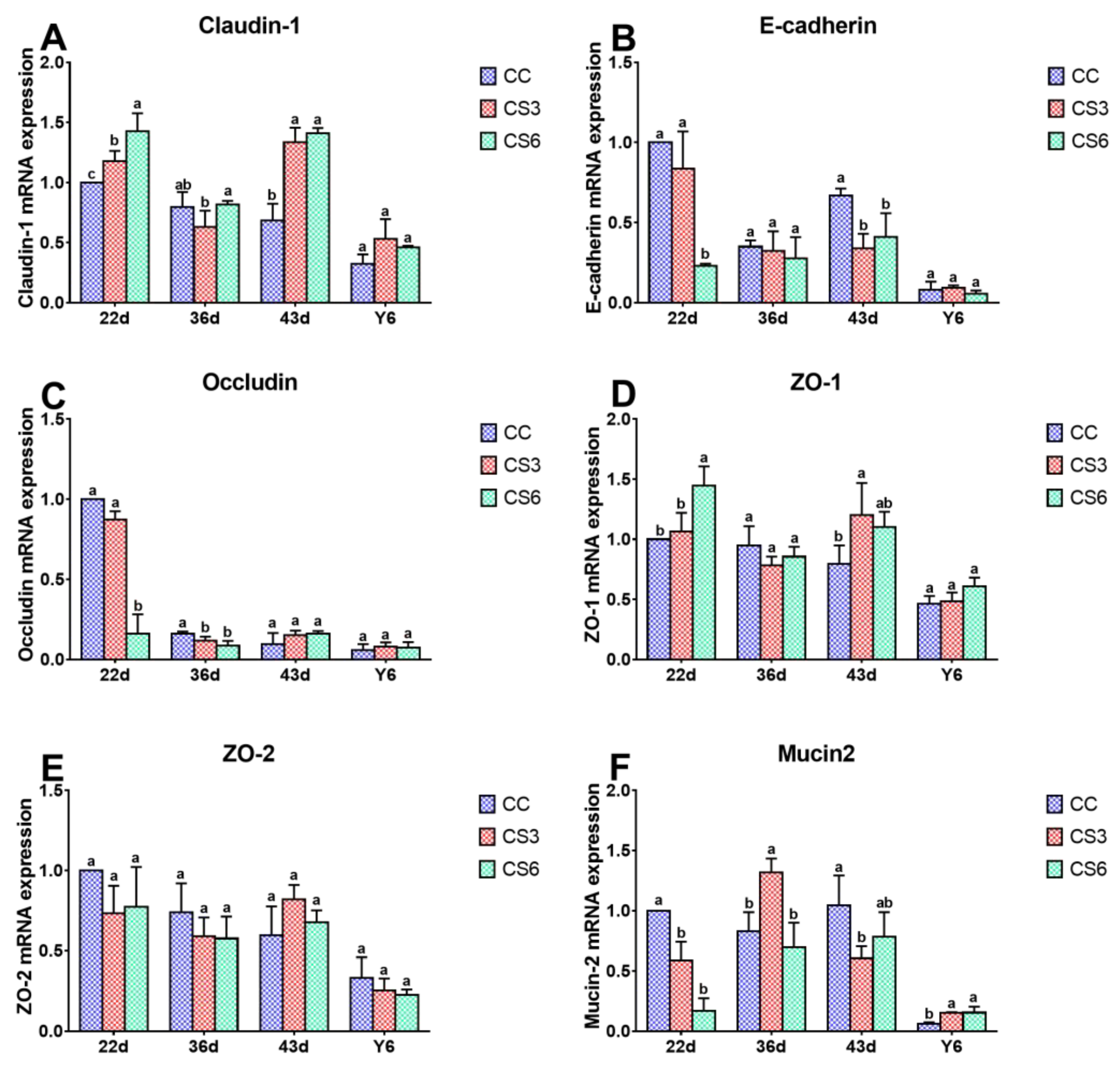
3.4. mRNA Levels of Intestinal Barrier Genes in the Jejunum
3.5. Protein Levels of Intestinal Barrier Genes in the Jejunum
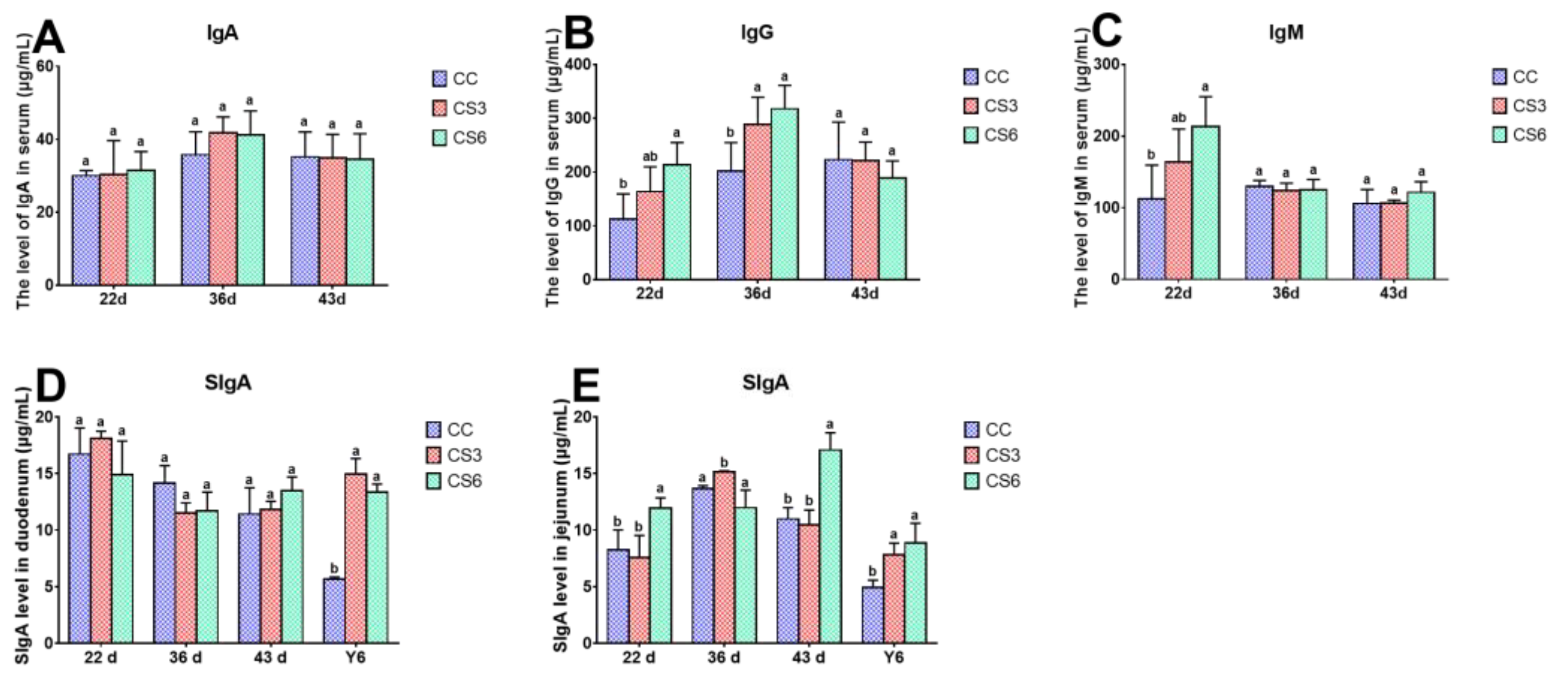
3.6. Levels of Immunoglobulin in Serum, Duodenum and Jejunum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aarif, O.; Shergojry, S.A.; Dar, S.A.; Khan, N.; Mir, N.A.; Sheikh, A.A. Impact of Cold Stress on Blood Biochemical and Immune Status in Male and Female Vanaraja Chickens. Indian J. Anim. Res. 2014, 48, 139–142. [Google Scholar] [CrossRef]
- Su, Y.Y.; Zhang, X.; Xin, H.W.; Li, S.; Li, J.F.; Zhang, R.X.; Li, X.; Li, J.H.; Bao, J. Effects of prior cold stimulation on inflammatory and immune regulation in ileum of cold-stressed broilers. Poult. Sci. 2018, 97, 4228–4237. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.Y.; Cheng, H.W. Warm perches: A novel approach for reducing cold stress effect on production, plasma hormones, and immunity in laying hens. Poult. Sci. 2021, 100, 101294. [Google Scholar] [CrossRef]
- Hirata, Y.; Broquet, A.H.; Menchén, L.; Kagnoff, M.F. Activation of Innate Immune Defense Mechanisms by Signaling through RIG-I/IPS-1 in Intestinal Epithelial Cells. J. Immunol. 2007, 179, 5425–5432. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.N.F.; Souza, G.T.; Schaffner, D.; Oliveira, T.C.M.; Maciel, J.F.; Souza, E.L.; Magnani, M. Changes in thermo-tolerance and survival under simulated gastrointestinal conditions of Salmonella Enteritidis PT4 and Salmonella Typhimurium PT4 in chicken breast meat after exposure to sequential stresses. Int. J. Food Microbiol. 2017, 251, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Theerawatanasirikul, S.; Koomkrong, N.; Kayan, A.; Boonkaewwan, C. Intestinal barrier and mucosal immunity in broilers, Thai Betong, and native Thai Praduhangdum chickens. Turk. J. Vet. Anim. Sci. 2017, 41, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Brandtzaeg, P. The gut as communicator between environment and host: Immunological consequences. Eur. J. Pharmacol. 2011, 668, S16–S32. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Ulluwishewa, D.; Anderson, R.C.; McNabb, W.C.; Moughan, P.J.; Wells, J.M.; Roy, N.C. Regulation of tight junction permeability by intestinal bacteria and dietary components. J. Nutr. 2011, 141, 769–776. [Google Scholar] [CrossRef] [Green Version]
- Fanning, A.S.; Jameson, B.J.; Jesaitis, L.A.; Anderson, J.M. The Tight Junction Protein ZO-1 Establishes a Link between the Transmembrane Protein Occludin and the Actin Cytoskeleton. J. Biol. Chem. 1998, 273, 29745–29753. [Google Scholar] [CrossRef] [Green Version]
- Catalioto, R.M.; Maggi, C.A.; Giuliani, S. Intestinal epithelial barrier dysfunction in disease and possible therapeutical interventions. Curr. Med. Chem. 2011, 18, 398–426. [Google Scholar] [CrossRef]
- Krause, G.; Winkler, L.; Mueller, S.L.; Haseloff, R.F.; Piontek, J.; Blasig, I.E. Structure and function of claudins. Biochim. Biophys. Acta 2008, 1778, 631–645. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Guo, Y.; Wang, Z. beta-1,3/1,6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult. Sci. 2013, 92, 1764–1773. [Google Scholar] [CrossRef] [PubMed]
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.L.; Sakai, M.; Sa, L.R.; Ferreira, A.J.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.F.; Chen, Y.P.; Chen, R.; Su, Y.; Zhang, R.Q.; He, Q.F.; Wang, K.; Wen, C.; Zhou, Y.M. Dietary mannan oligosaccharide ameliorates cyclic heat stress-induced damages on intestinal oxidative status and barrier integrity of broilers. Poult. Sci. 2019, 98, 4767–4776. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Han, H.; Zhang, L.; Wang, T. Bisdemethoxycurcumin attenuates lipopolysaccharide-induced intestinal damage through improving barrier integrity, suppressing inflammation, and modulating gut microbiota in broilers. J. Anim. Sci. 2021, 99, skab296. [Google Scholar] [CrossRef]
- Yang, T.; Liu, B.; Wang, Y.; Huang, X.; Yan, Z.; Jiang, Q.; Chen, Q. Ellagic Acid Improves Antioxidant Capacity and Intestinal Barrier Function of Heat-Stressed Broilers via Regulating Gut Microbiota. Animals 2022, 12, 1180. [Google Scholar] [CrossRef]
- Zhao, F.Q.; Zhang, Z.W.; Yao, H.D.; Wang, L.L.; Liu, T.; Yu, X.Y.; Li, S.; Xu, S.W. Effects of cold stress on mRNA expression of immunoglobulin and cytokine in the small intestine of broilers. Res. Vet. Sci. 2013, 95, 146–155. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2015, 10, e0138975. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.T.; Li, S.; Zhao, N.; Xing, L.; Gong, R.X.; Li, T.T.; Zhang, S.J.; Li, J.H.; Bao, J. Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum. Animals 2022, 12, 3260. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Chen, Z. The protective effect of gamma-aminobutyric acid on the development of immune function in chickens under heat stress. J. Anim. Physiol. Anim. Nutr. 2016, 100, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Xue, G.; Yin, J.; Zhao, N.; Liu, Y.; Fu, Y.; Zhang, R.; Bao, J.; Li, J. Intermittent mild cold stimulation improves the immunity and cold resistance of spleens in broilers. Poult. Sci. 2021, 100, 101492. [Google Scholar] [CrossRef] [PubMed]
- Leeson, S. Nutritional considerations of poultry during heat stress. World’s Poult. Sci. J. 1986, 42, 69–81. [Google Scholar] [CrossRef]
- Mashaly, M.M.; Hendricks, G.L., 3rd; Kalama, M.A.; Gehad, A.E.; Abbas, A.O.; Patterson, P.H. Effect of heat stress on production parameters and immune responses of commercial laying hens. Poult. Sci. 2004, 83, 889–894. [Google Scholar] [CrossRef]
- Sagher, B.M. The effect of cold stress on muscle growth in young chicks. Growth 1975, 39, 281–288. [Google Scholar]
- Blahová, J.; Dobšíková, R.; Straková, E.; Suchý, P. Effect of Low Environmental Temperature on Performance and Blood System in Broiler Chickens (Gallus domesticus). Acta Vet. Brno 2007, 76, S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Shinder, D.L.; Luger, D.; Rusal, M.; Rzepakovsky, V.; Bresler, V.; Yahav, S. Early age cold conditioning in broiler chickens(Gallus domesticus): Thermotolerance and growth responses. J. Therm. Biol. 2002, 27, 517–523. [Google Scholar] [CrossRef]
- Dokladny, K.; Moseley, P.L.; Ma, T.Y. Physiologically relevant increase in temperature causes an increase in intestinal epithelial tight junction permeability. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G204–G212. [Google Scholar] [CrossRef]
- Zhou, H.J.; Kong, L.L.; Zhu, L.X.; Hu, X.Y.; Busye, J.; Song, Z.G. Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef]
- Umeda, K.; Ikenouchi, J.; Katahira-Tayama, S.; Furuse, K.; Sasaki, H.; Nakayama, M.; Matsui, T.; Tsukita, S.; Furuse, M.; Tsukita, S. ZO-1 and ZO-2 independently determine where claudins are polymerized in tight-junction strand formation. Cell 2006, 126, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Cano, A.; Perez-Moreno, M.A.; Rodrigo, I.; Locascio, A.; Blanco, M.J.; del Barrio, M.G.; Portillo, F.; Nieto, M.A. The transcription factor snail controls epithelial-mesenchymal transitions by repressing E-cadherin expression. Nat. Cell Biol. 2000, 2, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shao, D.; Wu, S.; Song, Z.; Shi, S. Heat stress-induced intestinal barrier damage and dimethylglycine alleviates via improving the metabolism function of microbiota gut brain axis. Ecotoxicol. Environ. Saf. 2022, 244, 114053. [Google Scholar] [CrossRef]
- Fihn, B.M.; Sjoqvist, A.; Jodal, M. Permeability of the rat small intestinal epithelium along the villus-crypt axis: Effects of glucose transport. Gastroenterology 2000, 119, 1029–1036. [Google Scholar] [CrossRef] [PubMed]
- Woof, J.M.; Kerr, M.A. The function of immunoglobulin A in immunity. J. Pathol. 2006, 208, 270–282. [Google Scholar] [CrossRef] [Green Version]
- Carr, D.J.; Woolley, T.W.; Blalock, J.E. Phentolamine but not propranolol blocks the immunopotentiating effect of cold stress on antigen-specific IgM production in mice orally immunized with sheep red blood cells. Brain Behav. Immun. 1992, 6, 50–63. [Google Scholar] [CrossRef]
- Park, S.O.; Hwangbo, J.; Ryu, C.M.; Park, B.S.; Chae, H.S.; Choi, H.C.; Kang, H.K.; Seo, O.S.; Choi, Y.H. Effects of extreme heat stress on growth performance, lymphoid organ, IgG and cecum microflora of broiler chickens. Int. J. Agric. Biol. 2013, 15, 120. [Google Scholar]
- Thaxton, P. Influence of temperature on the immune response of birds. Poult. Sci. 1978, 57, 1430–1440. [Google Scholar] [CrossRef]
- Chen, W.; Yin, C.; Li, J.; Sun, W.; Li, Y.; Wang, C.; Pi, Y.; Cordero, G.; Li, X.; Jiang, X. Stimbiotics Supplementation Promotes Growth Performance by Improving Plasma Immunoglobulin and IGF-1 Levels and Regulating Gut Microbiota Composition in Weaned Piglets. Biology 2023, 12, 441. [Google Scholar] [CrossRef]
- Lin, H.; Lin, J.; Pan, T.; Li, T.; Jiang, H.; Fang, Y.; Wang, Y.; Wu, F.; Huang, J.; Zhang, H.; et al. Polymeric immunoglobulin receptor deficiency exacerbates autoimmune hepatitis by inducing intestinal dysbiosis and barrier dysfunction. Cell Death Dis. 2023, 14, 68. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reagent | Usage |
|---|---|
| 5× gDNA Eraser Buffer | 2.0 µL |
| gDNA Eraser | 1.0 µL |
| Total RNA | 1 µg |
| RNase Free dH2O up to | 10.0 µL |
| 42 °C Water-bath for 2 min | |
| 5× PrimeScript Buffer 2 | 4.0 µL |
| RT Primer Mix | 1.0 µL |
| PrimeScript RT Enzyme Mix I | 1.0 µL |
| RNase Free dH2O | 4.0 µL |
| 37 °C Water-bath for 15 min, then 85 °C Water-bath for 5 s | |
| Gene | Gene Reference Sequence | Primer Sequences (5′-3′) |
|---|---|---|
| β-actin | NM_205518.1 | Forward: CACCACAGCCGAGAGAGAAAT |
| Reverse: TGACCATCAGGGAGTTCATAGC | ||
| Claudin-1 | NM_001013611.2 | Forward: TGGAGGATGACCAGGTGAAGA |
| Reverse: CGAGCCACTCTGTTGCCATA | ||
| E-cadherin | NM 001039258.2 | Forward: GACAGGGACATGAGGCAGAA |
| Reverse: GCCGTGACAATGCCATTCTC | ||
| Occludin | NM 205128.1 | Forward: TCATCGCCTCCATCGTCTAC |
| Reverse: TCTTACTGCGCGTCTTCTGG | ||
| ZO-1 | XM 413773.4 | Forward: TGTAGCCACAGCAAGAGGTG |
| Reverse: CTGGAATGGCTCCTTGTGGT | ||
| ZO-2 | XM_025144669.1 | Forward: CGGCAGCTATCAGACCACTC |
| Reverse: CACAGACCAGCAAGCCTACAG | ||
| Mucin2 | XM_421035 | Forward: CAGCACCAACTTCTCAGTTC |
| Reverse: TCTGCAGCCACACATTCTTT |
| CC | CS3 | CS6 | p Value | |
|---|---|---|---|---|
| Feed conversion ratio | 2.01 ± 0.07 a | 1.84 ± 0.06 b | 1.98 ± 0.07 a | 0.000 |
| Daily feed intake (kg) | 1.21 ± 0.37 a | 1.21 ± 0.36 a | 1.24 ± 0.40 a | 0.163 |
| Daily weight gain (kg) | 0.60 ± 0.14 b | 0.65 ± 0.15 a | 0.62 ± 0.14 b | 0.008 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xing, L.; Li, T.; Zhang, Y.; Bao, J.; Wei, H.; Li, J. Intermittent and Mild Cold Stimulation Maintains Immune Function Stability through Increasing the Levels of Intestinal Barrier Genes of Broilers. Animals 2023, 13, 2138. https://doi.org/10.3390/ani13132138
Xing L, Li T, Zhang Y, Bao J, Wei H, Li J. Intermittent and Mild Cold Stimulation Maintains Immune Function Stability through Increasing the Levels of Intestinal Barrier Genes of Broilers. Animals. 2023; 13(13):2138. https://doi.org/10.3390/ani13132138
Chicago/Turabian StyleXing, Lu, Tingting Li, Yong Zhang, Jun Bao, Haidong Wei, and Jianhong Li. 2023. "Intermittent and Mild Cold Stimulation Maintains Immune Function Stability through Increasing the Levels of Intestinal Barrier Genes of Broilers" Animals 13, no. 13: 2138. https://doi.org/10.3390/ani13132138