
Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum
 and
and 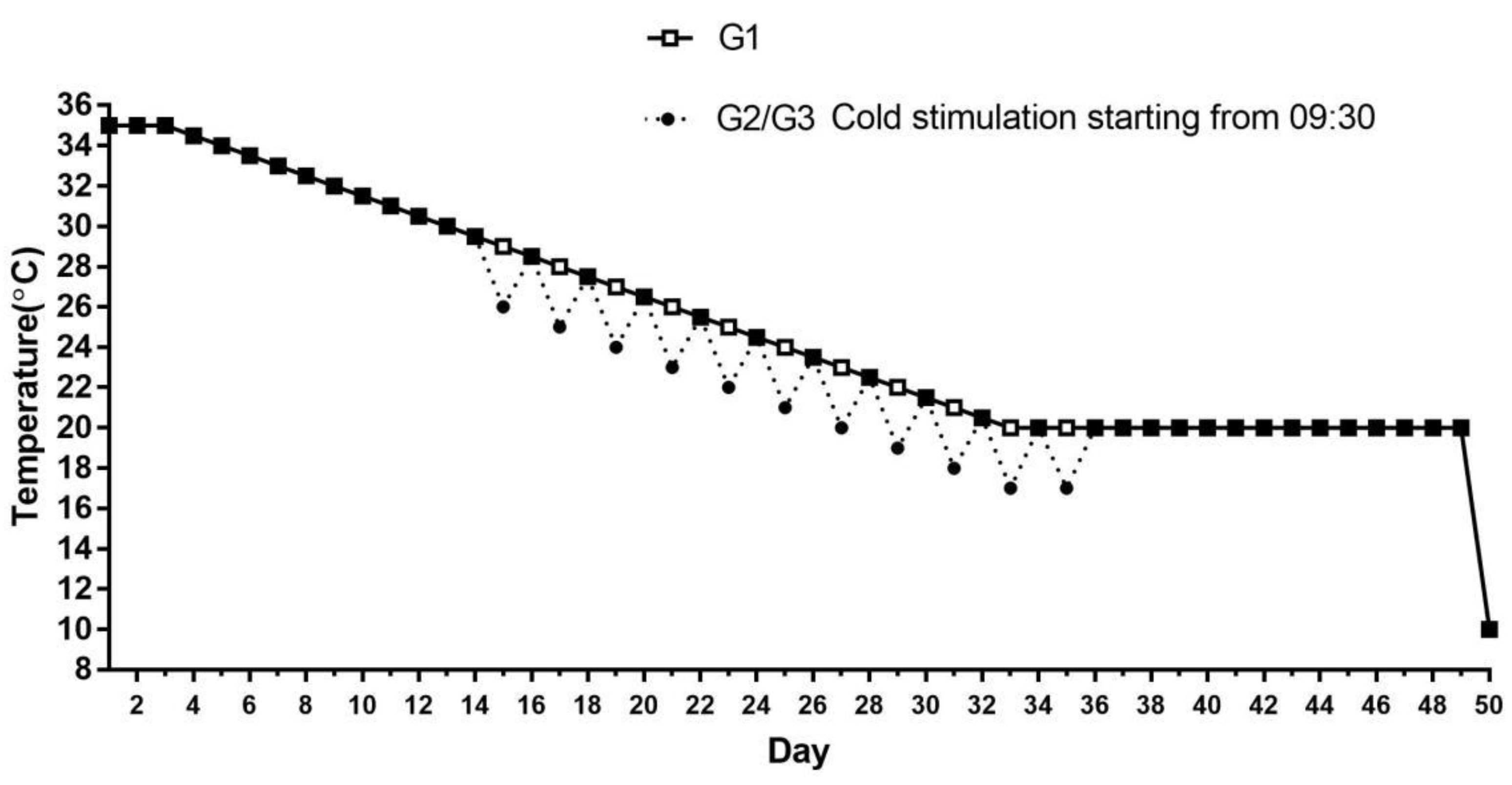{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Care and Experimental Design
2.2. RNA Extraction and Reverse Transcription
2.3. Quantitative Real-Time PCR Analysis
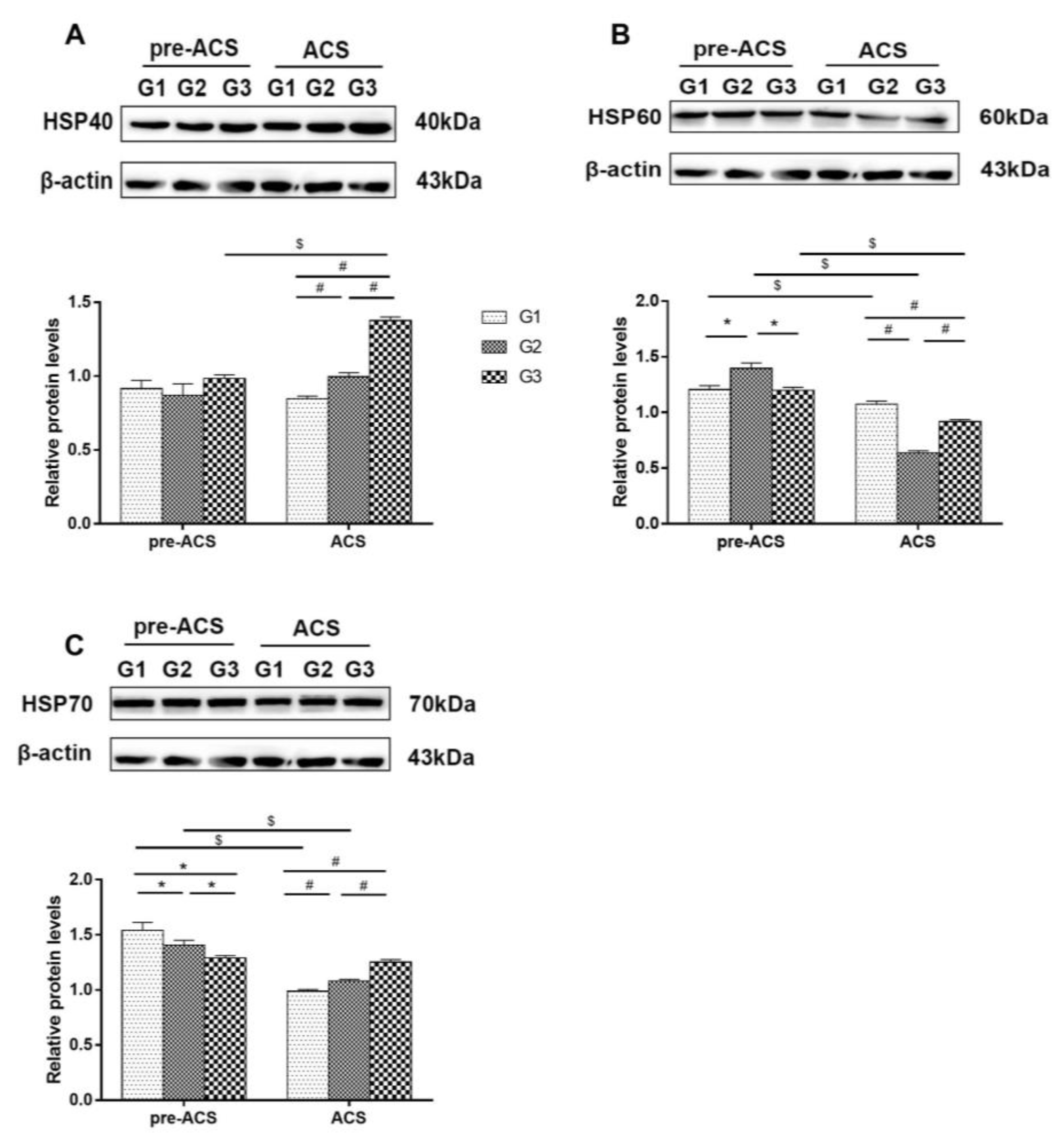
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
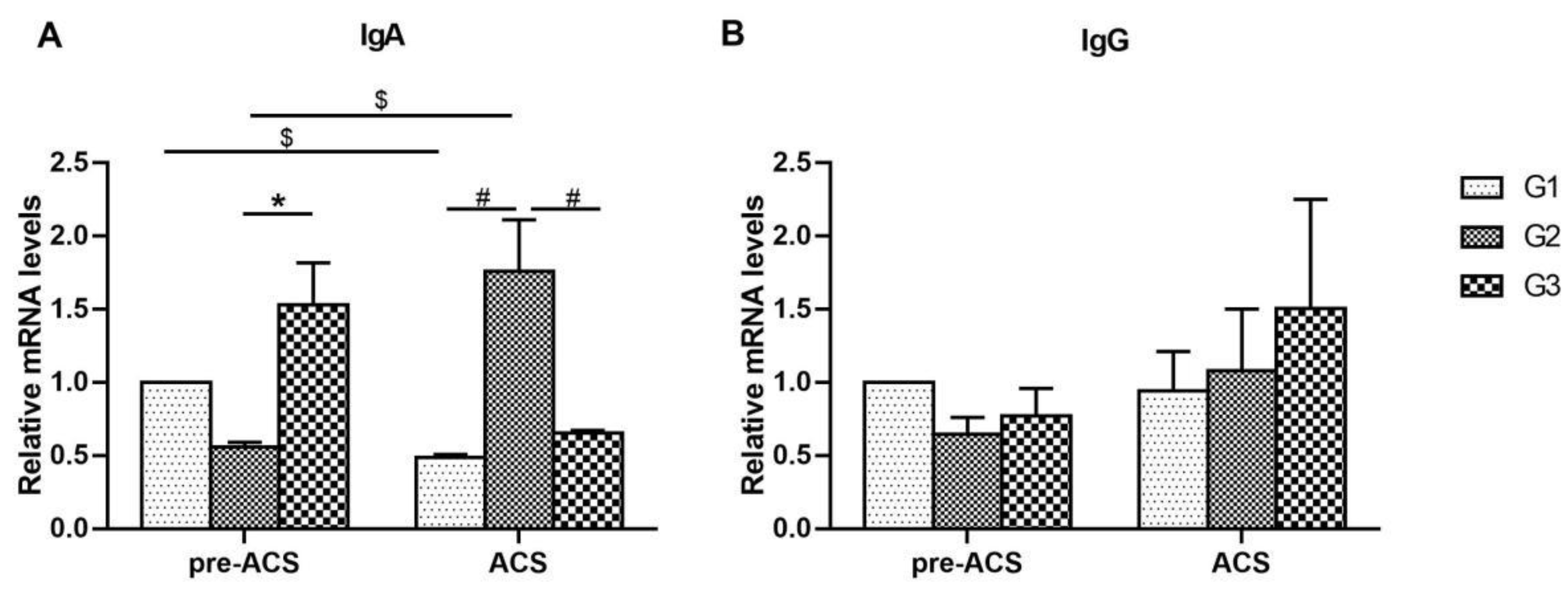
3.1. Relative Expression Levels of Immunoglobulins in Ileum Tissue
3.2. Relative Expression Levels of Cytokines in Ileum Tissue
3.3. Relative Expression Levels of Toll-Like Receptors in Ileum Tissue
3.4. Relative Expression Levels of Intestinal Barrier Genes in Ileum Tissue
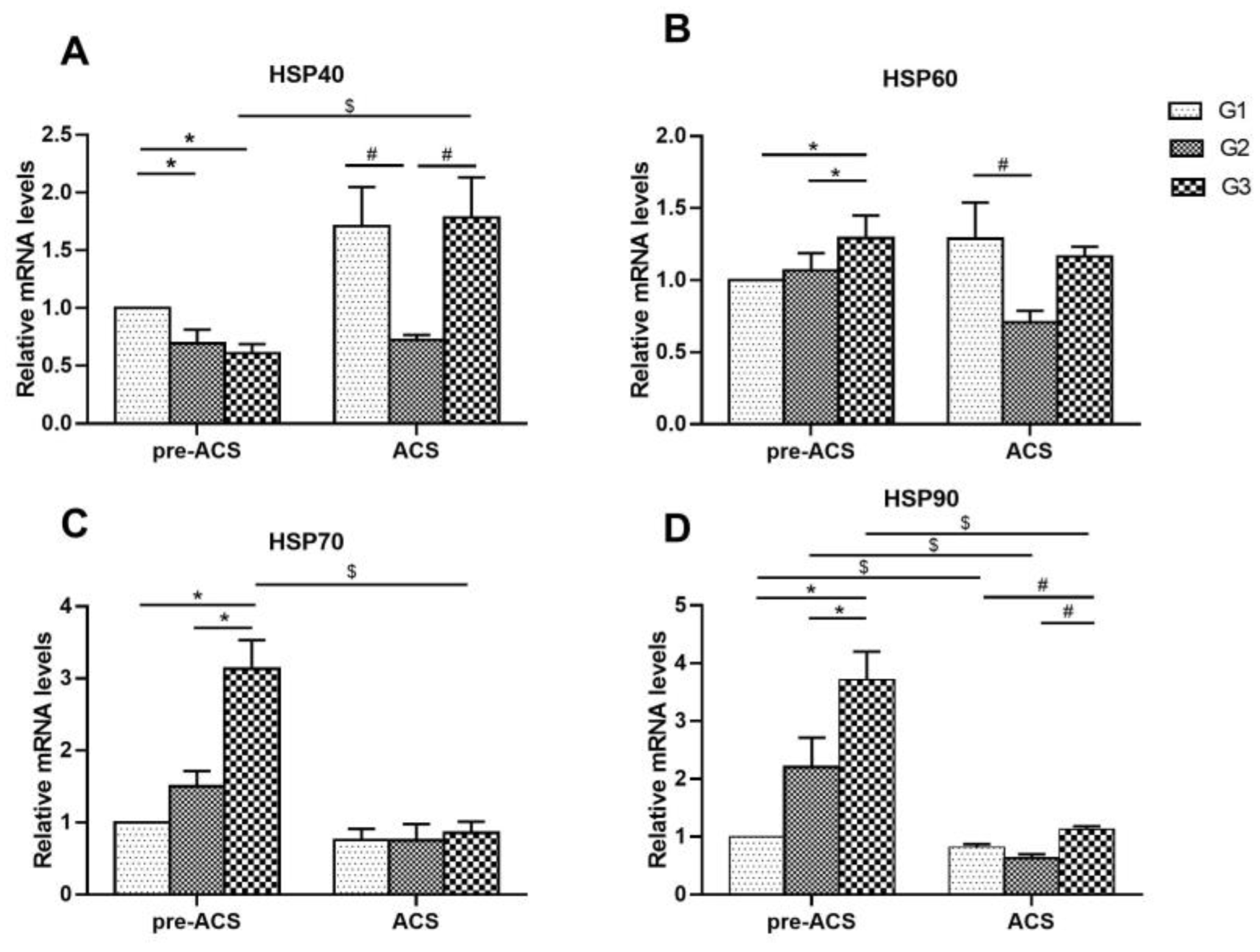
3.5. Relative Expression Levels of Heat Shock Proteins in Ileum Tissue
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nyuiadzi, D.; Berri, C.; Dusart, L.; Travel, A.; Méda, B.; Bouvarel, I.; Guilloteau, L.A.; Chartrin, P.; Coustham, V.; Praud, C.; et al. Short cold exposures during incubation and postnatal cold temperature affect performance, breast meat quality, and welfare parameters in broiler chickens. Poult. Sci. 2020, 99, 857–868. [Google Scholar] [CrossRef]
- Xie, S.S.; Yang, X.K.; Gao, Y.H.; Jiao, W.J.; Li, X.H.; Li, Y.J.; Ning, Z.H. Performance differences of Rhode Island Red, Bashang Long-tail Chicken, and their reciprocal crossbreds under natural cold stress. Asian-Australas J. Anim. Sci. 2017, 30, 1507–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarif, O.; Shergojry, S.A.; Dar, S.A.; Khan, N.; Mir, N.A.; Sheikh, A.A. Impact of cold stress on blood biochemical and immune status in male and female vanaraja chickens. Indian J. Anim. Res. 2014, 48, 139–142. [Google Scholar] [CrossRef]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. The effect of cold stress on the pathogenesis of necrotic enteritis in broiler chicks. Avian Pathol. 2015, 44, 430–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.Y.; Cheng, H.W. Warm perches: A novel approach for reducing cold stress effect on production, plasma hormones, and immunity in laying hens. Poult. Sci. 2021, 100, 101294. [Google Scholar] [CrossRef] [PubMed]
- Spinu, M.; Degen, A.A. Effect of cold stress on performance and immune responses of Bedouin and White Leghorn hens. Br. Poult. Sci. 1993, 34, 177–185. [Google Scholar] [CrossRef]
- Su, Y.Y.; Wei, H.D.; Bi, Y.J.; Wang, Y.A.; Zhao, P.; Zhang, R.X.; Li, J.H.; Bao, J. Pre-cold acclimation improves the immune function of trachea and resistance to cold stress in broilers. J. Cell. Physiol. 2018, 234, 7198–7212. [Google Scholar] [CrossRef]
- Liu, Y.H.; Xue, G.; Li, S.; Fu, Y.J.; Yin, J.W.; Zhang, R.X.; Li, J.H. Effect of Intermittent and Mild Cold Stimulation on the Immune Function of Bursa in Broilers. Animals 2020, 10, 1275. [Google Scholar] [CrossRef]
- Xue, G.; Yin, J.W.; Zhao, N.; Liu, Y.H.; Fu, Y.J.; Zhang, R.X.; Bao, J.; Li, J.H. Intermittent mild cold stimulation improves the immunity and cold resistance of spleens in broilers. Poult. Sci. 2021, 100, 101492. [Google Scholar] [CrossRef]
- Li, J.H.; Huang, F.F.; Li, X.; Su, Y.Y.; Li, H.T.; Bao, J. Effects of intermittent cold stimulation on antioxidant capacity and mRNA expression in broilers. Livest. Sci. 2017, 204, 110–114. [Google Scholar] [CrossRef]
- Shinder, D.; Luger, D.; Rusal, M.; Rzepakosky, V.; Bresler, V.; Yahav, S. Early age cold conditioning in broiler chickens (Gallus domesticus): Thermotolerance and growth responses. J. Therm. Biol. 2002, 27, 517–523. [Google Scholar] [CrossRef]
- Su, Y.Y.; Li, S.; Xin, H.W.; Li, J.F.; Li, X.; Zhang, R.X.; Li, J.H.; Bao, J. Proper cold stimulation starting at an earlier age can enhance immunity and improve adaptability to cold stress in broilers. Poult. Sci. 2020, 99, 129–141. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.; Greene, E.; Ishola, P.; Huff, G.; Donoghue, A.; Bottje, W.; Dridi, S. Chronic Mild Cold Conditioning Modulates the Expression of Hypothalamic Neuropeptide and Intermediary Metabolic-Related Genes and Improves Growth Performances in Young Chicks. PLoS ONE 2015, 10, e0142319. [Google Scholar] [CrossRef] [PubMed]
- Vordenbäumen, S.; Otte, J.-M. Role of Antimicrobial Peptides in Inflammatory Bowel Disease. Polymers 2011, 3, 2010–2017. [Google Scholar] [CrossRef]
- Wu, B.Y.; Guo, H.R.; Cui, H.M.; Peng, X.; Fang, J.; Zuo, Z.C.; Deng, J.L.; Wang, X.; Hang, J.Y. Pathway underlying small intestine apoptosis by dietary nickel chloride in broiler chickens. Chem. Biol. Interact. 2016, 24, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Simon, K.; Arts, J.A.J.; de Vries Reilingh, G.; Kemp, B.; Lammers, A. Effects of early life dextran sulfate sodium administration on pathology and immune response in broilers and layers. Poult. Sci. 2016, 95, 1529–1542. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke 2018, 13, 612–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selecká, E.; Levkut, M., Jr.; Revajová, V.; Levkutová, M.; Karaffová, V.; Ševčíková, Z.; Herich, R.; Levkut, M. Research Note: Immunocompetent cells in blood and intestine after administration of Lacto-Immuno-Vital in drinking water of broiler chickens. Poult. Sci. 2021, 100, 101282. [Google Scholar] [CrossRef]
- Olfati, A.; Mojtahedin, A.; Sadeghi, T.; Akbari, M.; Martinez, P.F. Comparison of growth performance and immune responses of broiler chicks reared under heat stress, cold stress and thermoneutral conditions. Span. J. Agric. Res. 2018, 16, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.D.; Zhang, R.X.; Su, Y.Y.; Bi, Y.J.; Li, X.; Zhang, X.; Li, J.H.; Bao, J. Effects of Acute Cold Stress after Long-Term Cold Stimulation on Antioxidant Status, Heat Shock Proteins, Inflammation and Immune Cytokines in Broiler Heart. Front. Physiol. 2018, 9, 1589. [Google Scholar] [CrossRef]
- Su, Y.Y.; Zhang, X.; Xin, H.W.; Li, S.; Li, J.H.; Bao, J. Effects of prior cold stimulation on inflammatory and immune regulation in ileum of cold-stressed broilers. Poult. Sci. 2018, 97, 4228–4237. [Google Scholar] [CrossRef]
- Perelman, J.M.; Pirogov, A.B.; Naumov, D.E.; Gassan, D.A.; Afanaseva, E.Y.; Kotova, O.O. Dynamics of Cell Pattern and Profile of Bronchial Cytokines in Response to Acute Cold Exposure in Asthma Patients with Cold Airway Hyperresponsiveness. Am. J. Respir. Crit. Care Med. 2020, 201, A7394. [Google Scholar] [CrossRef]
- Yildirim, N.C.; Yurekli, M. The effect of adrenomedullin and cold stress on interleukin-6 levels in some rat tissues. Clin. Exp. Immunol. 2010, 161, 171–175. [Google Scholar] [CrossRef]
- Vargovic, P.; Laukova, M.; Ukropec, J.; Manz, G.; Kvetnansky, R. Prior Repeated Stress Attenuates Cold-Induced Immunomodulation Associated with "Browning" in Mesenteric Fat of Rats. Cell. Mol. Neurobiol. 2018, 38, 349–361. [Google Scholar] [CrossRef]
- Takeuchi, O.; Akira, S. Pattern Recognition.n Receptors and Inflammation. Cell 2010, 140, 805–820. [Google Scholar] [CrossRef] [Green Version]
- Velová, H.; Gutowska-Ding, M.W.; Burt, D.W.; Vinkler, M. Toll-like Receptor Evolution in Birds: Gene Duplication, Pseudogenization, and Diversifying Selection. Mol. Biol. Evol. 2018, 35, 2170–2184. [Google Scholar] [CrossRef] [Green Version]
- Gan, F.; Liu, Q.; Liu, Y.; Huang, D.; Pan, C.; Song, S.; Huang, K. Lycium barbarum polysaccharides improve CCl 4 -induced liver fibrosis, inflammatory response and TLRs/NF-kB signaling pathway expression in wistar rats. Life Sci. 2018, 192, 205–212. [Google Scholar] [CrossRef]
- He, Z.Z.; Li, J.J.; Mei, Y.; Lv, Y.; Zhu, F.; Liu, R.; Wang, X.; Yao, F.H.; Zhang, L. The role and difference of TLR2 and TLR4 in rhabdomyolysis induced acute kidney injury in mice. Int. J. Clin. Exp. Pathol. 2018, 11, 1054–1061. [Google Scholar]
- Vandana, G.D.; Bagath, M.; Sejian, V.; Krishnan, G.; Beena, V.; Bhatta, R. Summer season induced heat stress impact on the expression patterns of different toll-like receptor genes in Malabari goats. Biol. Rhythm Res. 2019, 50, 446–482. [Google Scholar] [CrossRef]
- Li, S.; Li, J.H.; Liu, Y.H.; Li, C.; Zhang, R.X.; Bao, J. Effects of Intermittent Mild Cold Stimulation on mRNA Expression of Immunoglobulins, Cytokines, and Toll-Like Receptors in the Small Intestine of Broilers. Molecules 2020, 10, 1492. [Google Scholar] [CrossRef]
- Brownlie, R.; Allan, B. Avian toll-like receptors. Cell Tissue Res. 2011, 343, 121–130. [Google Scholar] [CrossRef]
- Theerawatanasirikul, S.; Koomkrong, N.; Kayan, A.; Boonkaewwan, C. Intestinal barrier and mucosal immunity in broilers, Thai Betong, and native Thai Praduhangdum chickens. Turk. J. Vet. Anim. Sci. 2017, 41, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Lynn, K.S.; Peterson, R.J.; Koval, M. Ruffles and spikes: Control of tight junction morphology and permeability by claudins. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183339. [Google Scholar] [CrossRef] [PubMed]
- Assimakopoulos, S.F.; Akinosoglou, K.; de Lastic, A.L.; Skintzl, A.; Mouzaki, A.; Gogos, C. The Prognostic Value of Endotoxemia and Intestinal Barrier Biomarker ZO-1 in Bacteremic Sepsis. Am. J. Med. Sci. 2019, 359, 100–107. [Google Scholar] [CrossRef]
- Zhou, H.J.; Kong, L.L.; Zhu, L.X.; Hu, X.Y.; Busye, J.; Song, Z.G. Effects of cold stress on growth performance, serum biochemistry, intestinal barrier molecules, and adenosine monophosphate-activated protein kinase in broilers. Animal 2021, 15, 100138. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Nighot, P.; Nighot, M.; Haque, M.; Rawat, M.; Ma, T.Y. Lactobacillus acidophilus Induces a Strain-specific and Toll-like Receptor 2-Dependent Enhancement of Intestinal Epithelial Tight Junction Barrier and Protection Against Intestinal Inflammation. Am. J. Pathol. 2021, 191, 872–884. [Google Scholar] [CrossRef]
- Zhang, Q.; Eicher, S.D.; Applegate, T.J. Development of intestinal mucin 2, IgA, and polymeric Ig receptor expressions in broiler chickens and Pekin ducks. Poult. Sci. 2015, 94, 172–180. [Google Scholar] [CrossRef]
- Cobo, E.R.; Kissoon-Singh, V.; Moreau, F.; Holani, R.; Chadee, K. MUC2 Mucin and Butyrate Contribute to the Synthesis of the Antimicrobial Peptide Cathelicidin in Response to Entamoeba histolytica- and Dextran Sodium Sulfate-Induced Colitis. Infect. Immun. 2017, 85, e00905-16. [Google Scholar] [CrossRef] [Green Version]
- Uerlings, J.; Song, Z.G.; Hu, X.Y.; Wang, S.K.; Lin, H.; Buyse, J.; Everaert, N. Heat exposure affects jejunal tight junction remodeling independently of adenosine monophosphate-activated protein kinase in 9-day-old broiler chicks. Poult. Sci. 2018, 97, 3681–3690. [Google Scholar] [CrossRef]
- Tsiouris, V.; Georgopoulou, I.; Batzios, C.; Pappaioannou, N.; Ducatelle, R.; Fortomaris, P. Heat stress as a predisposing factor for necrotic enteritis in broiler chicks. Avian Pathol. 2018, 47, 1–99. [Google Scholar] [CrossRef]
- Zheng, G.; Victor Fon, G.; Meixner, W.; Creekmore, A.; Zong, Y.K.; Dame, M.; Colacino, J.; Dedhia, P.H.; Hong, S.; Wiley, J.W. Chronic stress and intestinal barrier dysfunction: Glucocorticoid receptor and transcription repressor HES1 regulate tight junction protein Claudin-1 promoter. Sci. Rep. 2017, 7, 4502. [Google Scholar] [CrossRef]
- Hao, Y.; Feng, Y.J.; Li, J.L.; Gu, X.H. Role of MAPKs in HSP70's Protection against Heat Stress-Induced Injury in Rat Small Intestine. Biomed Res. Int. 2018, 2018, 1571406. [Google Scholar] [CrossRef] [Green Version]
- Flees, J.; Rajaei-Sharifabadi, H.; Greene, E.; Beer, L.; Hargis, B.M.; Ellestad, L.; Porter, T.; Donoghue, A.; Bottje, W.G.; Dridi, S. Effect of Morinda citrifolia (Noni)-Enriched Diet on Hepatic Heat Shock Protein and Lipid Metabolism-Related Genes in Heat Stressed Broiler Chickens. Front. Physiol. 2017, 27, 919. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Tang, S.; Song, E.; Yin, B.; Wu, D.; Bao, E. Hsp70 expression induced by Co-Enzyme Q10 protected chicken myocardial cells from damage and apoptosis under in vitro heat stress. Poult. Sci. 2017, 96, 1426–1437. [Google Scholar] [CrossRef]
- Zaglool, A.W.; Roushdy, E.M.; El-Tarabany, M.S. Impact of strain and duration of thermal stress on carcass yield, metabolic hormones, immunological indices and the expression of HSP90 and Myogenin genes in broilers. Res. Vet. Sci. 2018, 122, 193–199. [Google Scholar] [CrossRef]
- Shah, S.W.A.; Chen, J.; Han, Q.; Xu, Y.; Ishfaq, M.; Teng, X.H. Ammonia inhalation impaired immune function and mitochondrial integrity in the broilers bursa of fabricius: Implication of oxidative stress and apoptosis. Ecotoxicol. Environ. Saf. 2020, 190, 110078. [Google Scholar] [CrossRef]
- Shah, S.W.A.; Chen, D.; Zhang, J.Y.; Liu, Y.L.; Ishfaq, M.; Teng, X.H. The effect of ammonia exposure on energy metabolism and mitochondrial dynamic proteins in chicken thymus: Through oxidative stress, apoptosis, and autophagy. Ecotoxicol. Environ. Saf. 2020, 206, 111413. [Google Scholar] [CrossRef]
- Zhao, F.Q.; Zhang, Z.W.; Yao, H.D.; Wang, L.L.; Liu, T.; Yu, X.Y.; Li, S.; Xu, S.W. Effects of cold stress on mRNA expression of immunoglobulin and cytokine in the small intestine of broilers. Res. Vet. Sci. 2013, 95, 146–155. [Google Scholar] [CrossRef]
- Thaxton, P. Influence of temperature on the immune response of birds. Poult. Sci. 1978, 57, 1430–1440. [Google Scholar] [CrossRef]
- Lanzarini, N.M.; Bentes, G.A.; Volotão, E.M.; Pinto, M.A. Use of chicken immunoglobulin Y in general virology. J. Immunoass. Immunochem. 2018, 39, 235–248. [Google Scholar] [CrossRef]
- Carr, D.J.J.; Woolley, T.W.; Blalock, J.E. Phentolamine but not propranolol blocks the immunopotentiating effect of cold stress on antigen-specific IgM production in mice orally immunized with sheep red blood cells. Brain Behav. Immun. 1992, 6, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Felten, S.Y.; Madden, K.S.; Bellinger, D.L.; Kruszewska, B.; Moynihan, J.A.; Felten, D.L. The role of the sympathetic nervous system in the modulation of immune responses. Advan Pharmacol. 1998, 42, 583–587. [Google Scholar] [CrossRef]
- Al-Zghoul, M.B.; Saleh, K.M.; Ababneh, M.M.K. Effects of pre-hatch thermal manipulation and post-hatch acute heat stress on the mRNA expression of interleukin-6 and genes involved in its induction pathways in 2 broiler chicken breeds. Poult. Sci. 2019, 98, 1805–1819. [Google Scholar] [CrossRef] [PubMed]
- Shieh, Y.H.; Huang, H.M.; Wang, C.C.; Lee, C.C.; Fan, C.K.; Lee, Y.L. Zerumbone enhances the Th1 response and ameliorates ovalbumin-induced Th2 responses and airway inflammation in mice. Int. Immunopharmacol. 2015, 24, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.R.; Li, S.Z.; Fang, H.G. Different Duration of Cold Stress Enhances Pro-Inflammatory Cytokines Profile and Alterations of Th1 and Th2 Type Cytokines Secretion in Serum of Wistar Rats. J. Anim. Vet. Adv. 2012, 11, 1538–1545. [Google Scholar]
- Brenner, I.K.; Castellani, J.W.; Gabaree, C.; Young, A.J.; Zamecnik, J.; Shephard, R.J.; Shek, P.N. Immune changes in humans during cold exposure: Effects of prior heating and exercise. J. Appl. Physiol. 1999, 87, 699–710. [Google Scholar] [CrossRef]
- Monroy, F.P.; Banerjee, S.K.; Duong, T.; Aviles, H. Cold stress-induced modulation of inflammatory responses and intracerebral cytokine mRNA expression in acute murine toxoplasmosis. J. Parasitol. 1999, 85, 878–886. [Google Scholar] [CrossRef]
- Alcala, A.; Osborne, B.; Allen, B.; Seaton-Terry, A.; Kirkland, T.; Whalen, M. Toll-like receptors in the mechanism of tributyltin-induced production of pro-inflammatory cytokines, IL-1β and IL-6. Toxicology 2022, 30, 153177. [Google Scholar] [CrossRef]
- Titi, L.O.; Mohammed, I.; Amoaku, W.M. Toll-like Receptor Signalling Pathways and the Pathogenesis of Retinal Diseases. Front. Ophthalmol. 2022, 2, 850394. [Google Scholar] [CrossRef]
- Wu, M.H.; Zhang, P.; Huang, X. Toll-like receptors in innate immunity and infectious diseases. Front. Med. China 2010, 4, 385–393. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Calefi, A.S.; Cruz, D.S.G.; Aloia, T.P.A.; Zager, A.; Astolfi-Ferreira, C.S.; Piantino Ferreira, J.A.; Sharif, S.; Palermo-Neto, J. Heat stress decreases expression of the cytokines, avian beta-defensins 4 and 6 and Toll-like receptor 2 in broiler chickens infected with Salmonella Enteritidis. Vet. Immunol. Immunopathol. 2017, 40, 19–28. [Google Scholar] [CrossRef]
- Basu, M.; Paichha, M.; Swain, B.; Lenka, S.S.; Singh, S.; Chakrabarti, R.; Samanta, M. Modulation of TLR2, TLR4, TLR5, NOD1 and NOD2 receptor gene expressions and their downstream signaling molecules following thermal stress in the Indian major carp catla (Catla catla). 3 Biotech 2015, 5, 1021–1030. [Google Scholar] [CrossRef]
- Varasteh, S.; Braber, S.; Akbari, P.; Garssen, J.; Fink-Gremmels, J. Differences in Susceptibility to Heat Stress along the Chicken Intestine and the Protective Effects of Galacto-Oligosaccharides. PLoS ONE 2017, 10, e0138975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreedhar, A.S.; Csermely, P. Heat shock proteins in the regulation of apoptosis: New strategies in tumor therapy: A comprehensive review. Pharmacol. Ther. 2004, 101, 227–257. [Google Scholar] [CrossRef]
- Khoso, P.A.; Yang, Z.J.; Liu, C.P.; Li, S. Selenium Deficiency Downregulates Selenoproteins and Suppresses Immune Function in Chicken Thymus. Biol. Trace Elem. Res. 2015, 167, 48–55. [Google Scholar] [CrossRef]
- Takada, M.; Otaka, M.; Takahashi, T.; Izumi, Y.; Tamaki, K.; Shibuya, T.; Sakamoto, N.; Osada, T.; Yamamoto, S.; Ishida, R.; et al. Overexpression of a 60-kDa heat shock protein enhances cytoprotective function of small intestinal epithelial cells. Life Sci. 2010, 86, 499–504. [Google Scholar] [CrossRef]
- Goloudina, A.R.; Demidov, O.N.; Garrido, C. Inhibition of HSP70: A challenging anti-cancer strategy. Cancer Lett. 2012, 325, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Puijvelde, G.H.M.; van Es, T.; Wanrooij, E.J.A.; Habets, K.L.L.; Vos, P.; Zee, R.; van Eden, W.; van Berkel, T.J.; Kuiper, J. Induction of oral tolerance to HSP60 or an HSP60-peptide activates T cell regulation and reduces atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2677–2683. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Li, S.; Zhao, N.; Xing, L.; Gong, R.; Li, T.; Zhang, S.; Li, J.; Bao, J. Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum. Animals 2022, 12, 3260. https://doi.org/10.3390/ani12233260
Liu X, Li S, Zhao N, Xing L, Gong R, Li T, Zhang S, Li J, Bao J. Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum. Animals. 2022; 12(23):3260. https://doi.org/10.3390/ani12233260
Chicago/Turabian StyleLiu, Xiaotao, Shuang Li, Ning Zhao, Lu Xing, Rixin Gong, Tingting Li, Shijie Zhang, Jianhong Li, and Jun Bao. 2022. "Effects of Acute Cold Stress after Intermittent Cold Stimulation on Immune-Related Molecules, Intestinal Barrier Genes, and Heat Shock Proteins in Broiler Ileum" Animals 12, no. 23: 3260. https://doi.org/10.3390/ani12233260