Seasonal Changes in the Antioxidant Activity and Biochemical Parameters of Goat Milk

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Climatic Conditions
2.2. Goat Milk Sampling
2.3. Measurement of the Total Amount of Water-Soluble Antioxidants
2.4. Determination of Vitamin A and Carotene according to the Method of Kondrakhin I.P. [28]
2.5. Determination of Vitamin C in Milk with 2,6-Dichlorophenolindophenol
2.6. Determination of the Biochemical Parameters of Milk
3. Results
3.1. Biochemical Composition of Goat Milk Depending on the Season
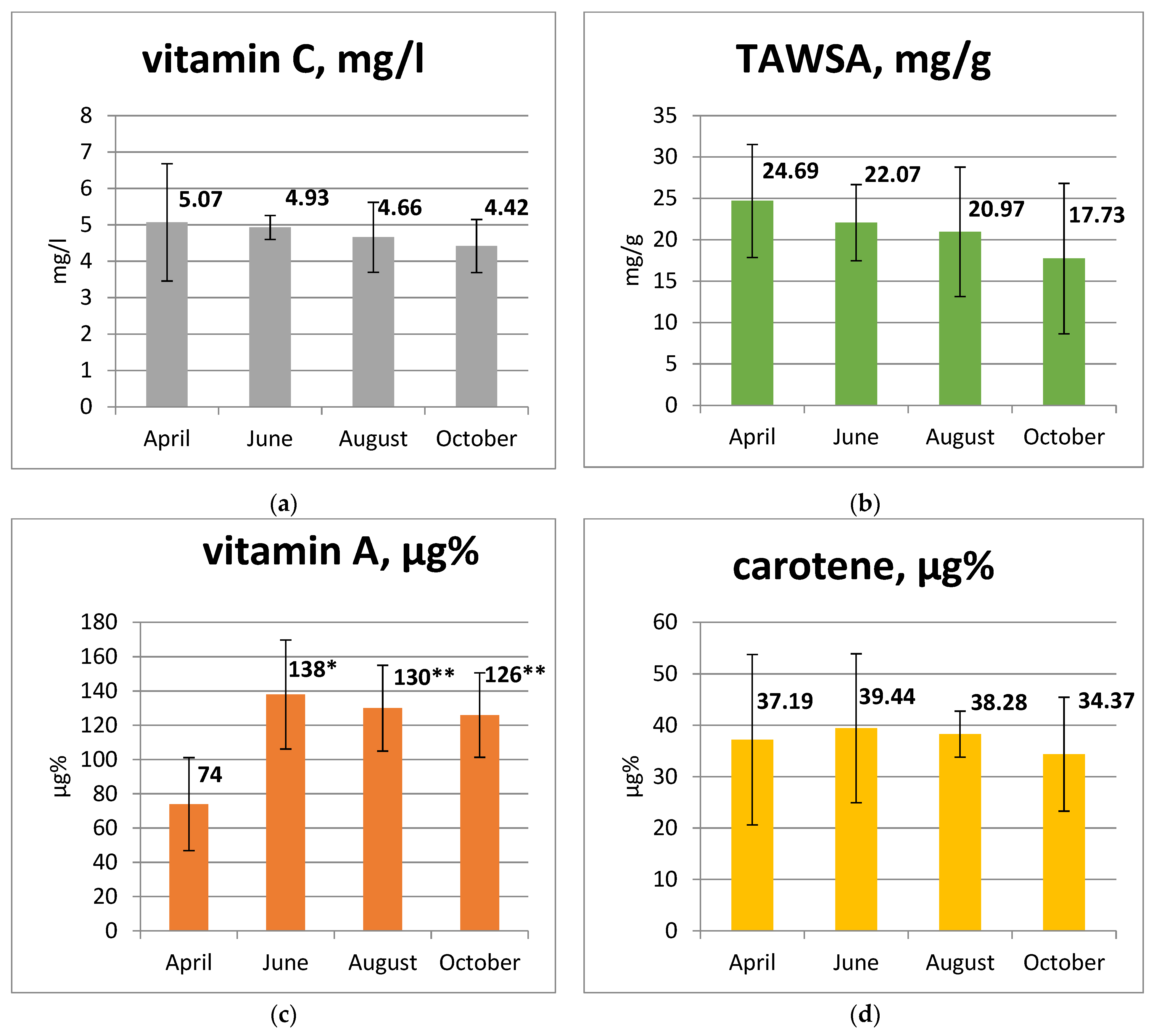
3.2. Antioxidant Composition of Goat Milk Depending on the Season
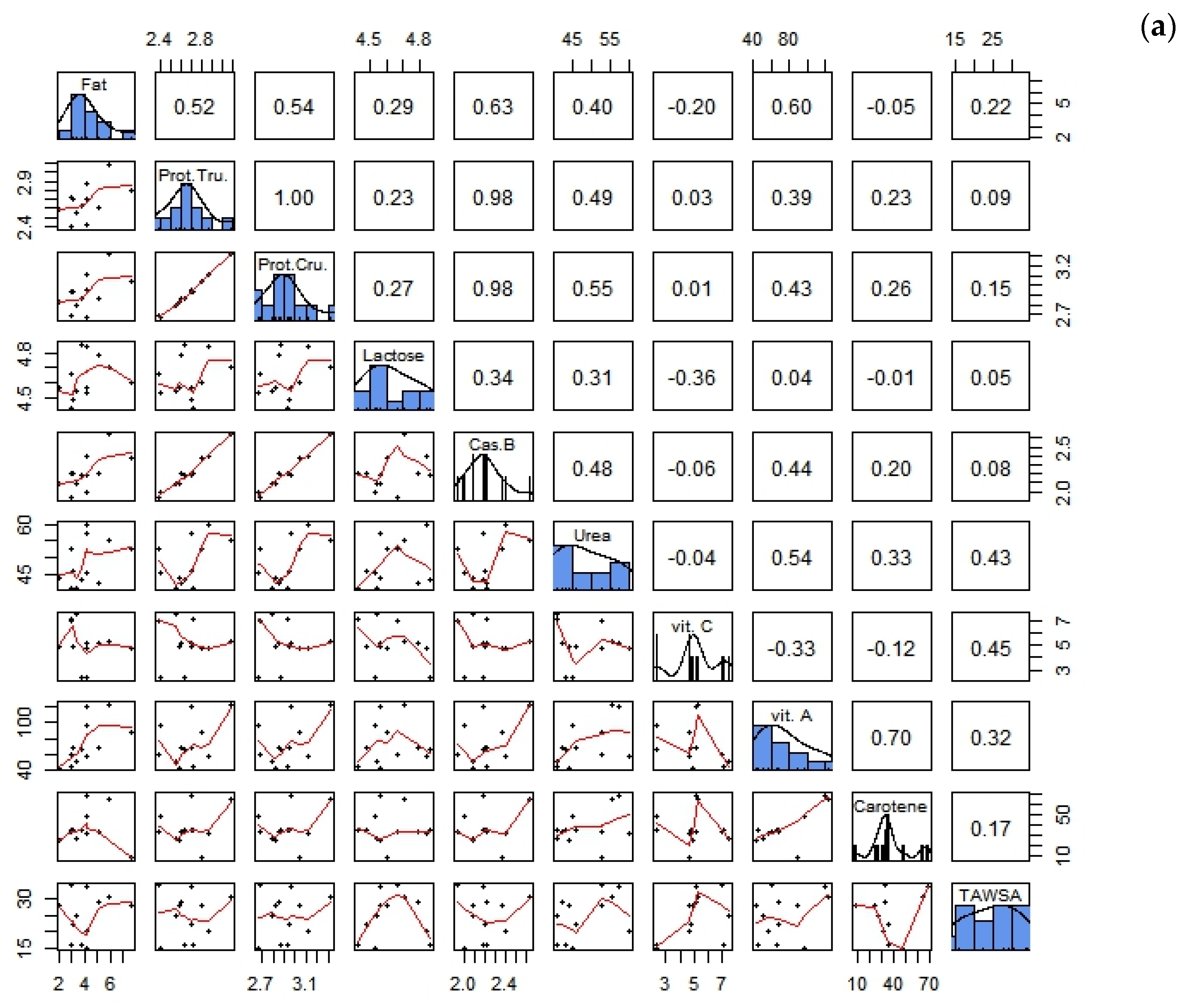
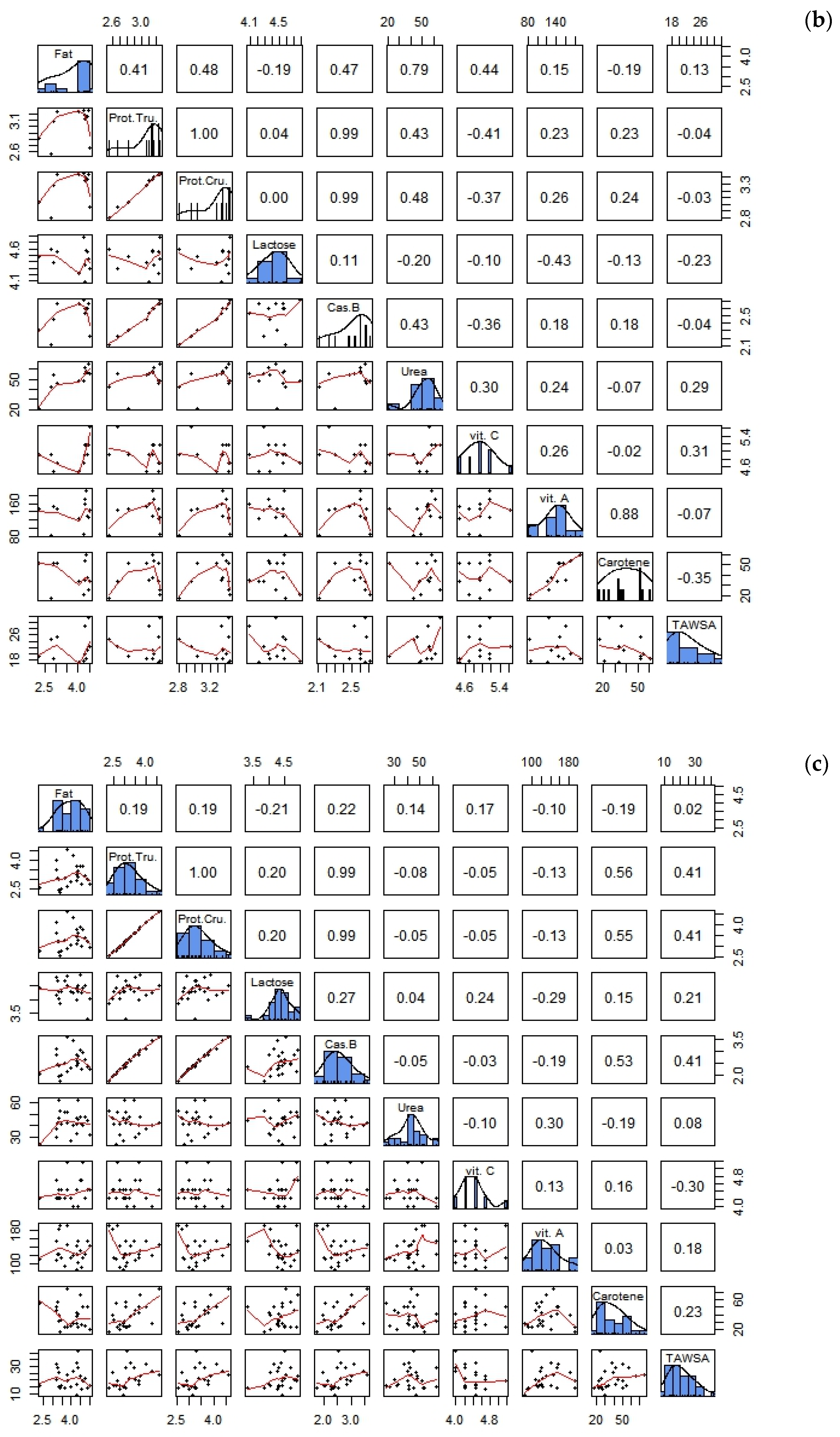
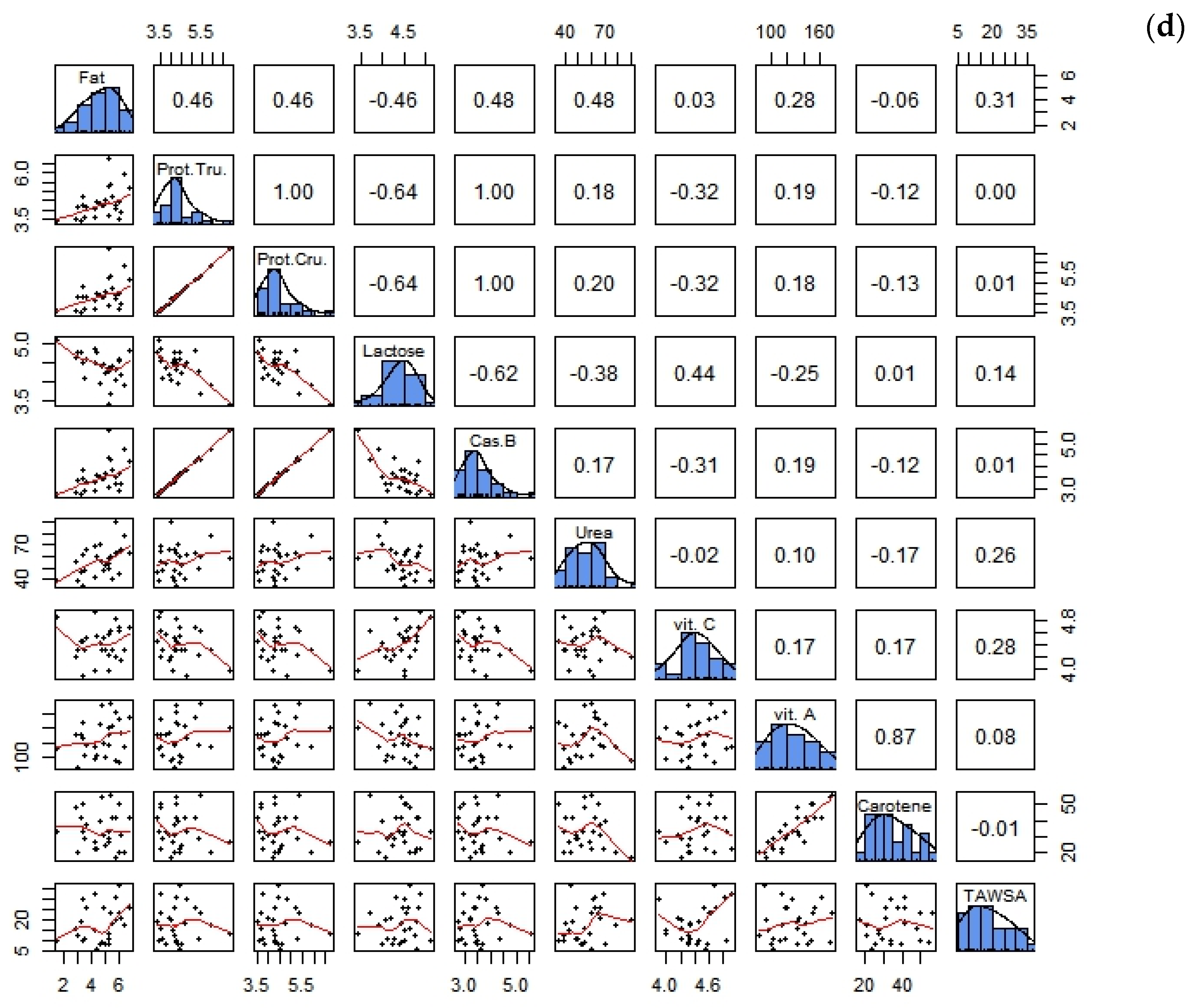
3.3. Correlations between Biochemical Parameters and the Antioxidant Composition of Goat Milk
4. Discussion
4.1. Comparability of Our Results with the Data Obtained Earlier
4.2. Physiological and Biochemical Mechanisms of the Adaptation of Goats to Heat Stress
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, I.T.; Nadeem, M.; Imran, M.; Ullah, R.; Ajmal, M.; Jaspal, M.H. Antioxidant properties of Milk and dairy products: A comprehensive review of the current knowledge. Lipids Health Dis. 2019, 18, 41. [Google Scholar] [CrossRef]
- Mercha, I.; Lakram, N.; Kabbour, M.R.; Bouksaim, M.; Zkhiri, F.; El Maadoudi, E.H. Probiotic and technological features of Enterococcus and Weissella isolates from camel milk characterised by an Argane feeding regimen. Arch. Microbiol. 2020, 202, 2207–2219. [Google Scholar] [CrossRef] [PubMed]
- Lad, S.S.; Aparnathi, K.; Mehta, B.; Velpula, S. Goat Milk in Human Nutrition and Health—A Review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1781–1792. [Google Scholar] [CrossRef]
- Kljajevic, N.V.; Tomasevic, I.B.; Miloradovic, Z.N.; Nedeljkovic, A.; Miocinovic, J.B.; Jovanovic, S.T. Seasonal variations of Saanen goat milk composition and the impact of climatic conditions. J. Food Sci. Technol. 2018, 55, 299–303. [Google Scholar] [CrossRef]
- Chauhan, S.S.; Rashamol, V.P.; Bagath, M.; Sejian, V.; Dunshea, F.R. Impacts of heat stress on immune responses and oxidative stress in farm animals and nutritional strategies for amelioration. Int. J. Biometeorol. 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Sejian, V.; Silpa, M.V.; Nair, M.R.R.; Devaraj, C.; Krishnan, G.; Bagath, M.; Chauhan, S.S.; Suganthi, R.U.; Fonseca, V.F.C.; König, S.; et al. Heat Stress and Goat Welfare: Adaptation and Production Considerations. Animals 2021, 11, 1021. [Google Scholar] [CrossRef]
- Habashy, W.S.; Milfort, M.C.; Rekaya, R.; Aggrey, S.E. Cellular antioxidant enzyme activity and biomarkers for oxidative stress are affected by heat stress. Int. J. Biometeorol. 2019, 63, 1569–1584. [Google Scholar] [CrossRef]
- Lima, A.R.C.; Silveira, R.M.F.; Castro, M.S.M.; De Vecchi, L.B.; Fernandes, M.H.M.D.R.; De Resende, K.T. Relationship between thermal environment, thermoregulatory responses and energy metabolism in goats: A comprehensive review. J. Therm. Biol. 2022, 109, 103324. [Google Scholar] [CrossRef]
- Sejian, V.; Bhatta, R.; Gaughan, J.; Dunshea, F.R.; Lacetera, N. Review: Adaptation of animals to heat stress. Animal 2018, 12, s431–s444. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Mondal, T. Heat stress and thermoregulatory responses of goats: A review. Biol. Rhythm. Res. 2021, 52, 407–433. [Google Scholar] [CrossRef]
- Milewski, K.Z.S.; Antoszkiewicz, Z.; Tański, A.S.Z. Impact of production season on the chemical composition and health properties of goat milk and rennet cheese. Emir. J. Food Agric. 2018, 30, 107–114. [Google Scholar] [CrossRef]
- Khan, I.T.; Bule, M.; Ullah, R.; Nadeem, M.; Asif, S.; Niaz, K. The antioxidant components of milk and their role in processing, ripening, and storage: Functional food. Vet.-World 2019, 12, 12–33. [Google Scholar] [CrossRef]
- Medeiros, G.K.; Queiroga, R.C.; Costa, W.K.; Gadelha, C.A.; Lacerda, R.R.E.; Lacerda, J.T.; Pinto, L.S.; Braganhol, E.; Teixeira, F.C.; Barbosa, P.P.D.S.; et al. Proteomic of goat milk whey and its bacteriostatic and antitumour potential. Int. J. Biol. Macromol. 2018, 113, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Fernandez, J.; Diaz-Castro, J.; Alférez, M.J.; Boesch, C.; Nestares, T.; López-Aliaga, I. Fermented goat milk improves antioxidant status and protects from oxidative damage to biomolecules during anemia recovery. J. Sci. Food Agric. 2017, 97, 1433–1442. [Google Scholar] [CrossRef] [PubMed]
- Manuelian, C.L.; Maggiolino, A.; De Marchi, M.; Claps, S.; Esposito, L.; Rufrano, D.; Casalino, E.; Tateo, A.; Neglia, G.; De Palo, P. Comparison of Mineral, Metabolic, and Oxidative Profile of Saanen Goat during Lactation with Different Mediterranean Breed Clusters under the Same Environmental Conditions. Animals 2020, 10, 432. [Google Scholar] [CrossRef]
- Shuvarikov, A.S.; Pastukh, O.N.; Zhukova, E.V.; Zheltova, O.A. The quality of milk of goats of Saanen, Alpine and Nubian breeds. IOP Conf. Series Earth Environ. Sci. 2021, 640, 032031. [Google Scholar] [CrossRef]
- Battini, M.; Barbieri, S.; Fioni, L.; Mattiello, S. Feasibility and validity of animal-based indicators for on-farm welfare assessment of thermal stress in dairy goats. Int. J. Biometeorol. 2016, 60, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Mayer, H.K.; Fiechter, G. Physical and chemical characteristics of sheep and goat milk in Austria. Int. Dairy J. 2012, 24, 57–63. [Google Scholar] [CrossRef]
- Li, S.; Delger, M.; Dave, A.; Singh, H.; Ye, A. Seasonal Variations in the Composition and Physicochemical Characteristics of Sheep and Goat Milks. Foods 2022, 11, 1737. [Google Scholar] [CrossRef]
- Margatho, G.; Rodríguez-Estévez, V.; Medeiros, L.; Simões, J. Seasonal variation of Serrana goat milk contents in mountain graz-ing system for cheese manufacture. Rev. Med. Vet. 2018, 169, 157–165. [Google Scholar]
- Zhukova, E.V.; Pastukh, O.N. Technological properties of milk depending on the season of the year. In Proceedings of the Scientific Developments and Innovations in Solving the Priority Tasks of Modern Zootechnics Conference, Kursk, Russia, 11 March 2021. [Google Scholar]
- Chávez-Servín, J.L.; Andrade-Montemayor, H.M.; Vázquez, C.V.; Barreyro, A.A.; García-Gasca, T.; Martínez, R.A.F.; Ramírez, A.M.O.; de la Torre-Carbot, K. Effects of feeding system, heat treatment and season on phenolic compounds and antioxidant capacity in goat milk, whey and cheese. Small Rumin. Res. 2018, 160, 54–58. [Google Scholar] [CrossRef]
- Hein, L.; Sørensen, L.; Kargo, M.; Buitenhuis, A. Genetic analysis of predicted fatty acid profiles of milk from Danish Holstein and Danish Jersey cattle populations. J. Dairy Sci. 2018, 101, 2148–2157. [Google Scholar] [CrossRef] [PubMed]
- Sarangi, S. Adaptability of goats to heat stress: A review. Pharma Innov. J. 2018, 7, 1114–1126. [Google Scholar]
- Collier, R.J.; Collier, J.L.; Rhoads, R.P.; Baumgard, L.H. Invited Review: Genes Involved in the Bovine Heat Stress Response. J. Dairy Sci. 2008, 91, 445–454. [Google Scholar] [CrossRef]
- Chamurliev, N.G. Functional state and morphological and biochemical parameters of blood during adaptation of dairy goats to the conditions of a sharply continental climate. In Proceedings of the Innovative Technologies in the Agro-Industrial Complex in Modern Economic Conditions Conference, Volgograd, Russia, 10–12 February 2021; pp. 309–313. [Google Scholar]
- Savina, A.A.; Voronina, O.A.; Bogolyubova, N.V.; Zaitsev, S.Y. Amperometric Detection of Antioxidant Activity of Model and Biological Fluids. Mosc. Univ. Chem. Bull. 2020, 75, 340–346. [Google Scholar] [CrossRef]
- Nadvodnyuk, A.I. Methods of Veterinary Clinical Laboratory Diagnostics: Handbook; Kolos: Moscow, Russia, 2004; pp. 186–188. [Google Scholar]
- Da Silva, T.L.; Aguiar-Oliveira, E.; Mazalli, M.R.; Kamimura, E.S.; Maldonado, R.R. Comparison between titrimetric and spectrophotometric methods for quantification of vitamin C. Food Chem. 2017, 224, 92–96. [Google Scholar] [CrossRef]
- Grelet, C.; Bastin, C.; Gelé, M.; Davière, J.-B.; Johan, M.; Werner, A.; Reding, R.; Fernandez Pierna, J.A.; Colinet, F.G.; Dardenne, P.; et al. Development of Fourier transform mid-infrared calibrations to predict acetone, β-hydroxybutyrate, and citrate contents in bovine milk through a European dairy network. J. Dairy Sci. 2016, 99, 4816–4825. [Google Scholar] [CrossRef]
- Kumar, H.; Yadav, D.; Kumar, N.; Seth, R.; Goyal, A.K. Nutritional and nutraceutical properties of goat milk-a review. Indian J. Dairy Sci. 2016, 69, 513–518. [Google Scholar]
- Choe, E.; Min, D.B. Mechanisms of Antioxidants in the Oxidation of Foods. Compr. Rev. Food Sci. Food Saf. 2009, 8, 345–358. [Google Scholar] [CrossRef]
- Ahmed, A.S.; El-Bassiony, T.; Elmalt, L.M.; Ibrahim, H.R. Identification of potent antioxidant bioactive peptides from goat milk proteins. Food Res. Int. 2015, 74, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Lenis Sanin, Y.; Zuluaga Cabrera, A.M.; Tarazona Morales, A.M. Adaptive responses to thermal stress in mammals. Rev. De Med. Vet. 2016, 31, 121–135. [Google Scholar]
- Huckabee, W.E. Relationships of pyruvate and lactate during anaerobic metabolism—I: Effects of infusion of pyruvate or glucose and of hyperventilation. J. Clin. Investig. 1958, 37, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Archana, P.R.; Aleena, J.; Pragna, P.; Vidya, M.K.; Abdul Niyas, P.A.; Bagath, M.; Krishnan, G.; Manimaran, A.; Beena, V.; Kurien, E.K.; et al. Role of Heat Shock Proteins in Livestock Adaptation to Heat Stress. J. Dairy Veter-Anim. Res. 2017, 5, 00127. [Google Scholar] [CrossRef]
- Abioja, M.; Logunleko, M.; Majekodunmi, B.; Adekunle, E.; Shittu, O.; Odeyemi, A.; Nwosu, E.; Oke, O.; Iyasere, O.; Abiona, J.; et al. Roles of candidate genes in the adaptation of goats to heat stress: A review. Small Rumin. Res. 2022, 218, 106878. [Google Scholar] [CrossRef]
- Cai, J.; Zhao, F.-Q.; Liu, J.-X.; Wang, D.-M. Local Mammary Glucose Supply Regulates Availability and Intracellular Metabolic Pathways of Glucose in the Mammary Gland of Lactating Dairy Goats Under Malnutrition of Energy. Front. Physiol. 2018, 9, 1467. [Google Scholar] [CrossRef] [PubMed]
- Sadovnikova, A.; Garcia, S.C.; Hovey, R.C. A Comparative Review of the Extrinsic and Intrinsic Factors Regulating Lactose Synthesis. J. Mammary Gland. Biol. Neoplasia 2021, 26, 197–215. [Google Scholar] [CrossRef]
- Lin, X.; Luo, J.; Zhang, L.; Zhu, J. MicroRNAs Synergistically Regulate Milk Fat Synthesis in Mammary Gland Epithelial Cells of Dairy Goats. Gene Expr. 2013, 16, 1–13. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | April | June | August | October |
|---|---|---|---|---|
| Mass fraction of fat MFF (%) | 4.14 ± 0.43 | 3.80 ± 0.23 | 3.94 ± 0.13 | 4.65 ± 0.24 |
| Mass fraction of true protein MFTP-1 (%) | 2.67 ± 0.06 | 3.06 ± 0.07 (p ≥ 0.01) * | 3.17 ± 0.12 (p ≥ 0.05) * | 4.37 ± 0.15 |
| Mass fraction of crude protein MFCP-2 (%) | 2.92 ± 0.05 (p ≥ 0.01) * | 3.28 ± 0.06 (p ≥ 0.001) * | 3.37 ± 0.11 (p ≥ 0.01) * | 4.44 ± 0.14 |
| Lactose (%) | 4.63 ± 0.04 | 4.44 ± 0.05 | 4.35 ± 0.07 | 4.41 ± 0.07 |
| Dry skimmed milk matter DSMM (%) | 8.17 ± 0.10 (p ≥ 0.05) * | 8.42 ± 0.10 (p ≥ 0.001) * | 8.39 ± 0.16 (p ≥ 0.001) * | 9.75 ± 0.14 |
| Total dry matter TDM (%) | 12.33 ± 0.47 | 12.24 ± 0.26 | 12.34 ± 0.21 (p ≥ 0.001) * | 14.24 ± 0.31 |
| Caseins (%) | 2.21 ± 0.06 (p ≥ 0.01) * | 2.51 ± 0.06 (p ≥ 0.001) * | 2.56 ± 0.09 (p ≥ 0.001) * | 3.55 ± 0.12 (p ≥ 0.001) * |
| Aceton (mM/L) | 0.15 ± 0.03 (p ≥ 0.001) * | 0.05 ± 0.03 | 0.08 ± 0.02 | 0 & |
| Betahydroxybutyrate BHB (mM/L) | 0.06 ± 0.01 (p ≥ 0.01) * | 0.01 ± 0.02 | 0.03 ± 0.01 (p ≥ 0.001) * | 0 & |
| Urea (mg/100 mL) | 48.51 ± 0.94 (p ≥ 0.001) * | 51.08 ± 3.31 | 43.01 ± 2.01 | 56.40 ± 2.49 |
| Freezing point (°C) | −0.547.25 ± 0.54 (p ≥ 0.001) * | −0.554 ± 0.22 | −0.562 ± 0.19 | −0.567 ± 0.24 |
| pH (a.u. #) | 6.60 ± 0.02 (p ≥ 0.001) * | 6.57 ± 0.02 | 6.53 ± 0.03 (p ≥ 0.001) * | 6.61 ± 0.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voronina, O.A.; Zaitsev, S.Y.; Savina, A.A.; Rykov, R.A.; Kolesnik, N.S. Seasonal Changes in the Antioxidant Activity and Biochemical Parameters of Goat Milk. Animals 2023, 13, 1706. https://doi.org/10.3390/ani13101706
Voronina OA, Zaitsev SY, Savina AA, Rykov RA, Kolesnik NS. Seasonal Changes in the Antioxidant Activity and Biochemical Parameters of Goat Milk. Animals. 2023; 13(10):1706. https://doi.org/10.3390/ani13101706
Chicago/Turabian StyleVoronina, Oksana A., Sergei Yu. Zaitsev, Anastasia A. Savina, Roman A. Rykov, and Nikita S. Kolesnik. 2023. "Seasonal Changes in the Antioxidant Activity and Biochemical Parameters of Goat Milk" Animals 13, no. 10: 1706. https://doi.org/10.3390/ani13101706