
RNAi Analysis of Potential Functions of Cyclin B3 in Reproduction of Male Oriental River Prawns (Macrobrachium nipponense)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Methods and Materials
2.1. Ethics Statement
2.2. Rapid Amplification of cDNA ends (RACE)
2.3. The qPCR Analysis
2.4. RNAi Analysis
2.5. Histological Observations
2.6. Statistical Analysis
3. Results
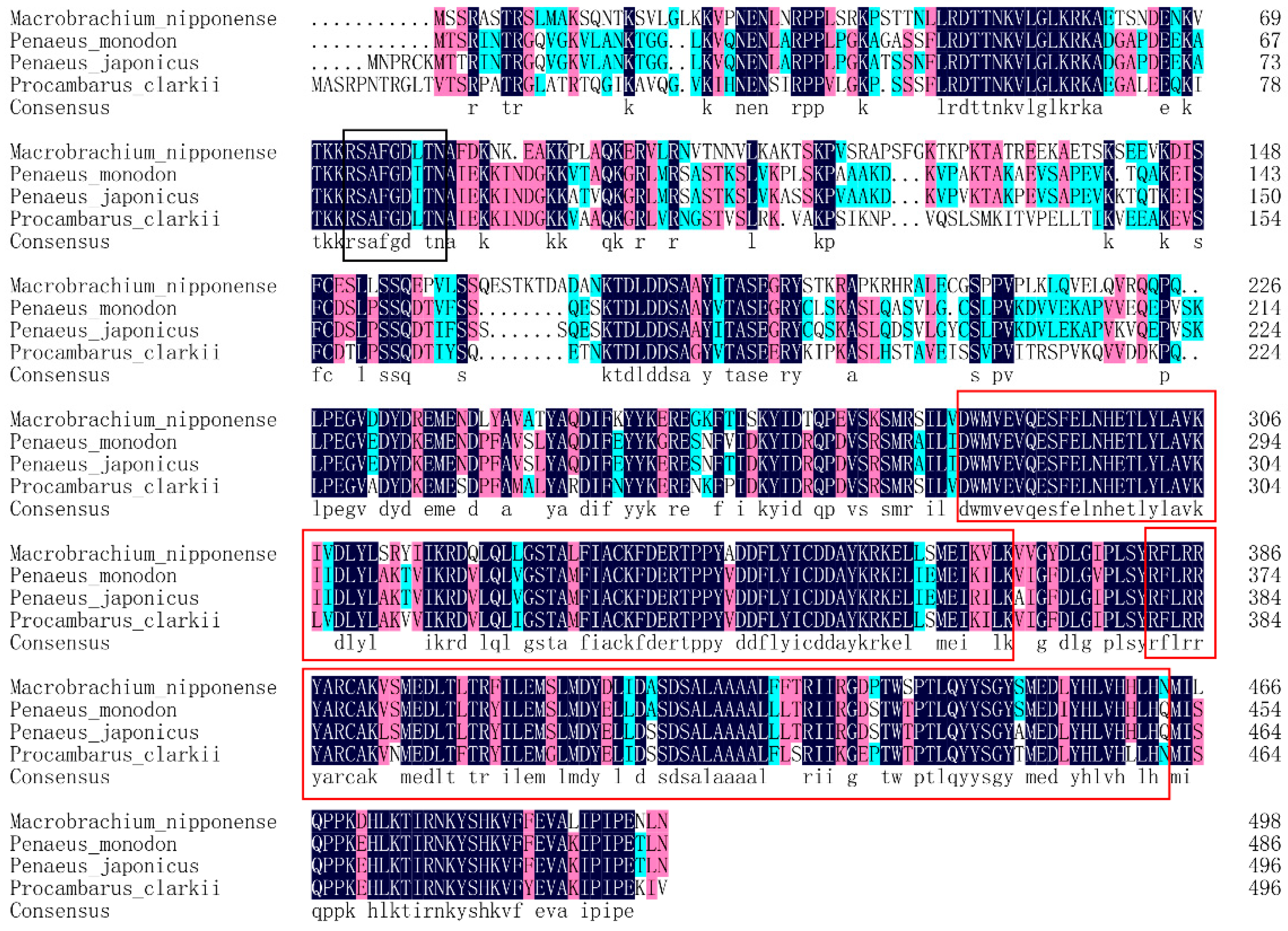
3.1. Sequence Analysis
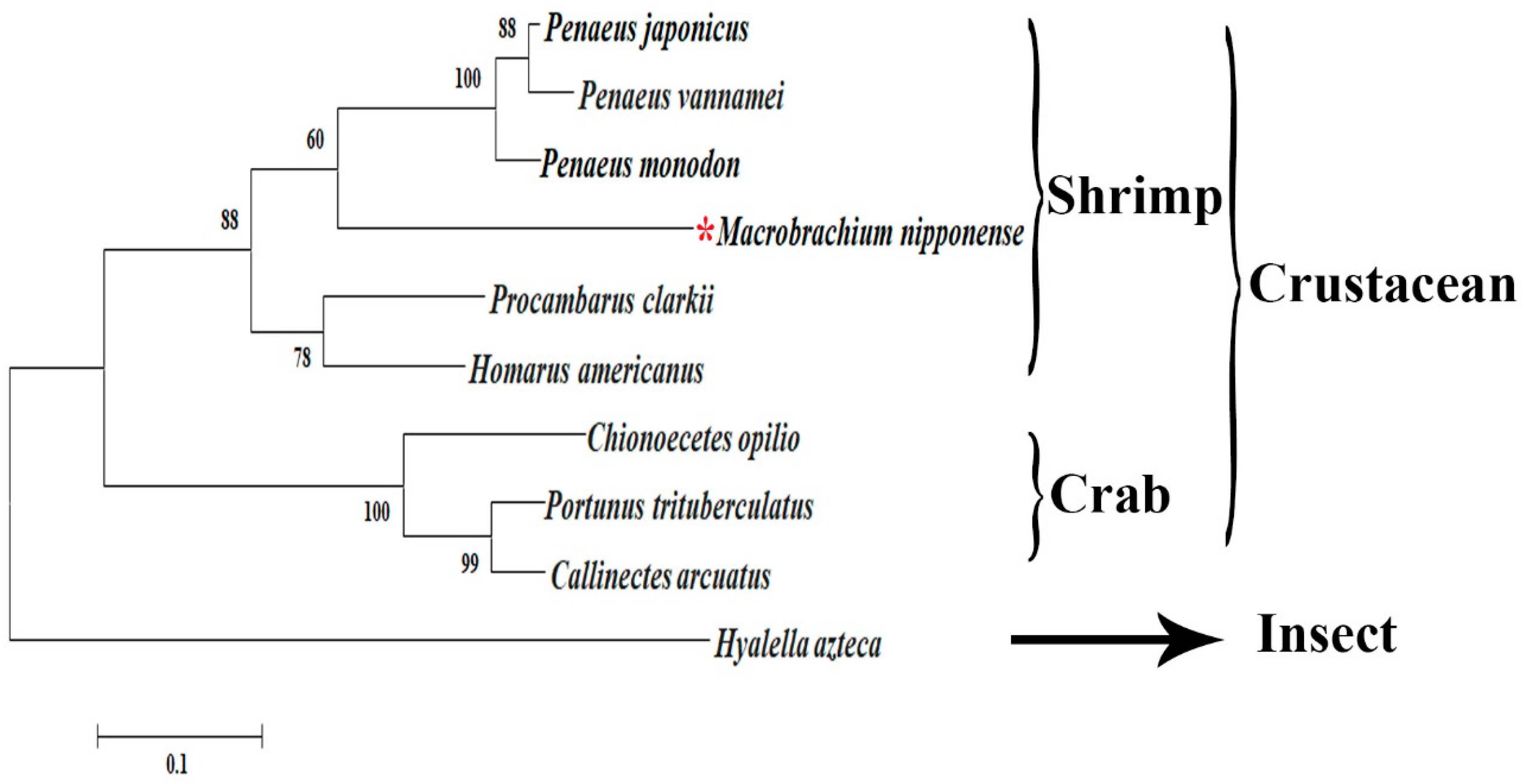
3.2. Phylogenetic Tree Analysis
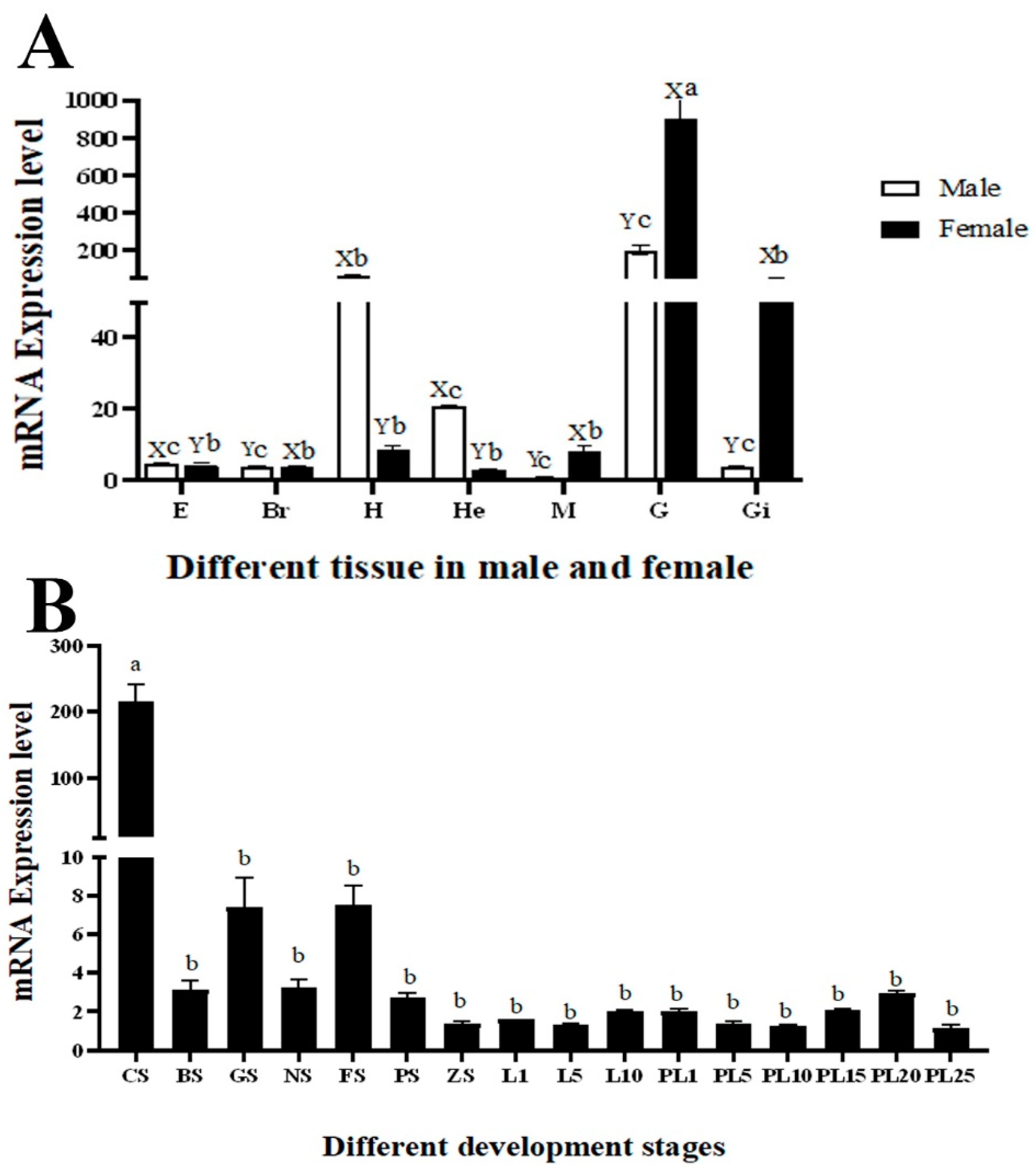
3.3. The qPCR Analysis
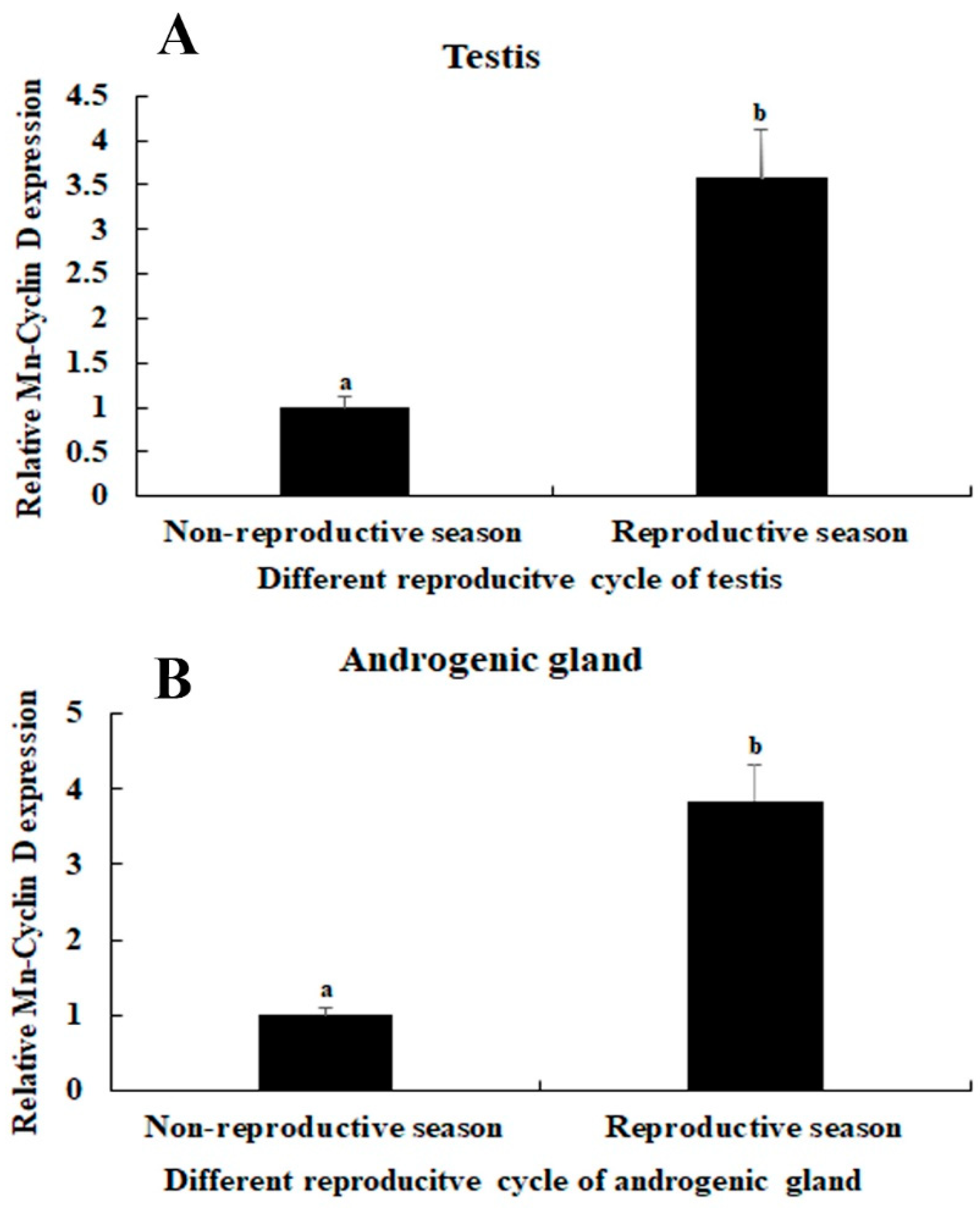
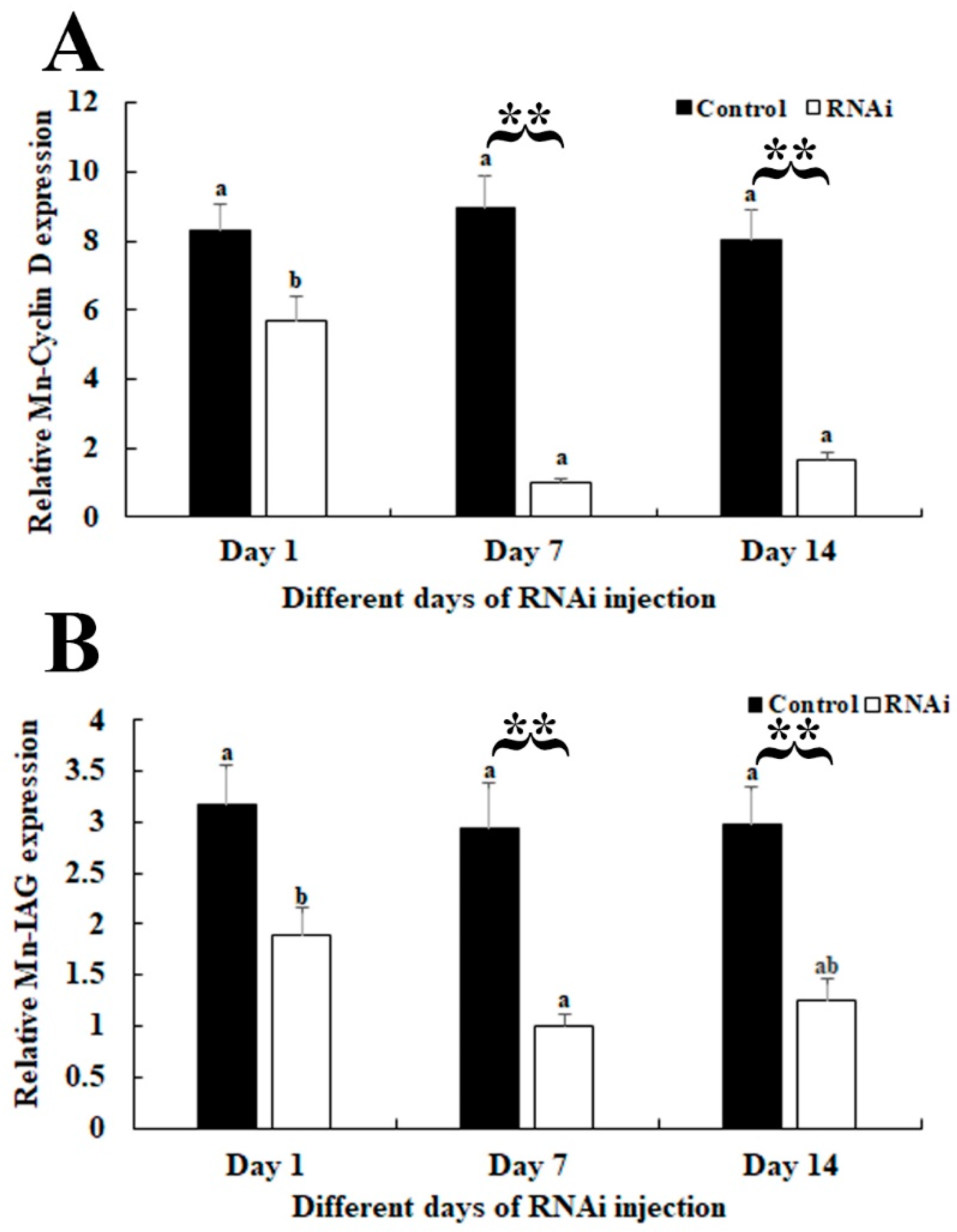
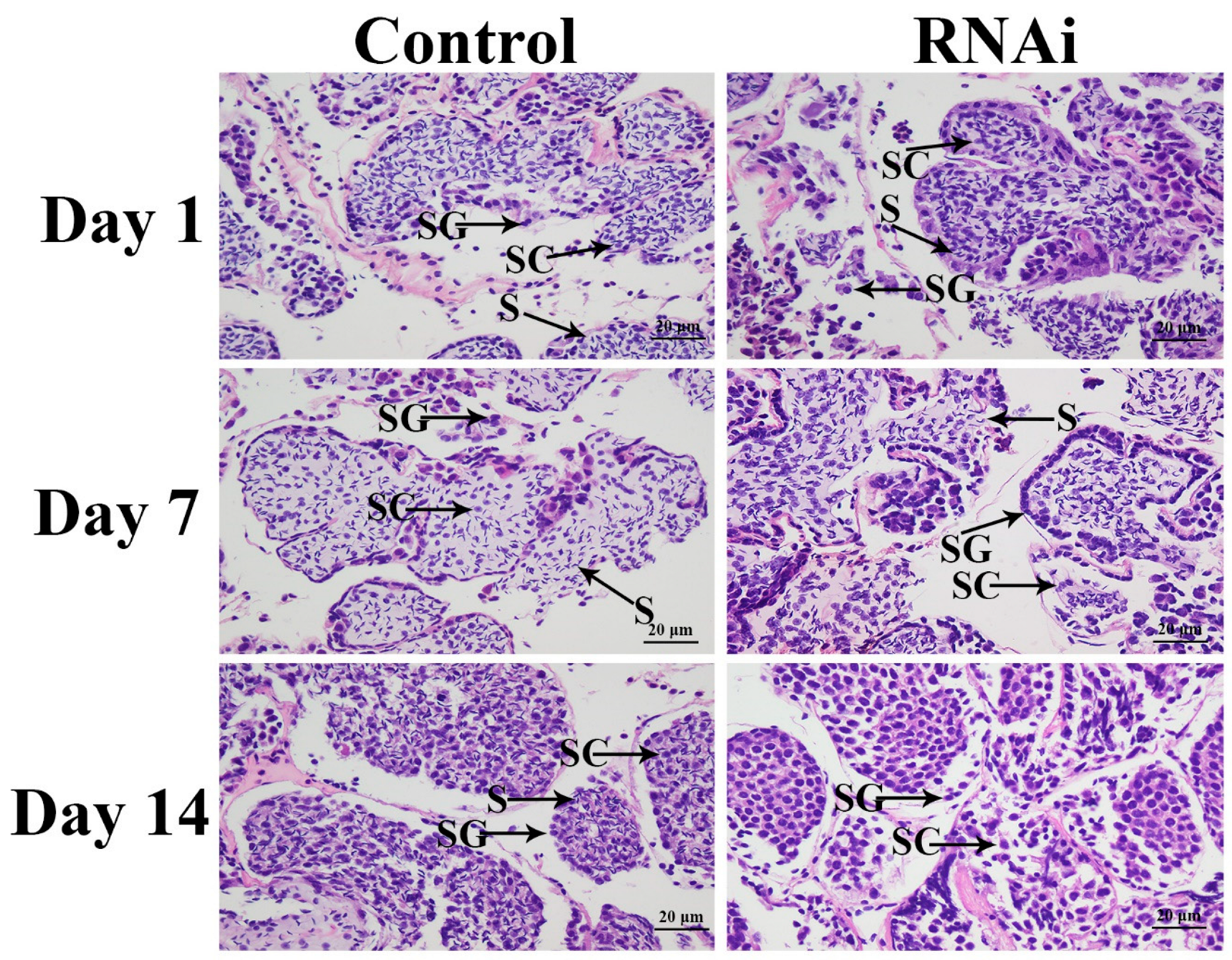
3.4. RNAi Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fu, H.T.; Jiang, S.F.; Xiong, Y.W. Current status and prospects of farming the giant river prawn (Macrobrachium rosenbergii) and the oriental river prawn (Macrobrachium nipponense) in china. Aquac. Res. 2012, 43, 993–998. [Google Scholar]
- Zhang, X.L.; Cui, L.F.; Li, S.M.; Liu, X.Z.; Han, X.; Jiang, K.Y.; Bureau of Fisheries, Ministry of Agriculture of the People's Republic of China. Fisheries economic statistics. In China Fishery Yearbook; Agricultural Press: Beijing China, 2020; Volume 24. [Google Scholar]
- Jin, S.B.; Zhang, Y.; Guang, H.H.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Histological observation of gonadal development during post-larva in oriental river prawn, Macrobrachium nipponense. Chin. J. Fish. 2016, 29, 11–16. [Google Scholar]
- Jin, S.B.; Fu, Y.; Hu, Y.N.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Identification of candidate genes of male sexual development from androgenic gland in Macrobrachium nipponense through performing long-reads and next generation transcriptome sequencing after eyestalk ablation. Sci. Rep. 2021, 11, 19855. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.B.; Fu, Y.; Hu, Y.N.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Transcriptome Profiling Analysis of the Testis After Eyestalk Ablation for Selection of the Candidate Genes Involved in the Male Sexual Development in Macrobrachium nipponense. Front. Genet. 2021, 12, 675928. [Google Scholar] [CrossRef] [PubMed]
- Khalaila, I.; Manor, R.; Weil, S.; Granot, Y.; Keller, R.; Sagi, A. The eyestalk-androgenic gland-testis endocrine axis in the crayfish Cherax quadricarinatus. Gen. Comp. Endocrinol. 2022, 127, 147–156. [Google Scholar] [CrossRef]
- Song, C.W.; Liu, L.; Hui, M.; Liu, Y.; Liu, H.R.; Cui, Z.X. Primary molecular basis of androgenic gland endocrine sex regulation revealed by transcriptome analysis in Eriocheir sinensis. J. Oceanol. Limnol. 2019, 37, 223–234. [Google Scholar] [CrossRef]
- Hopkins, P.M. The eyes have it: A brief history of crustacean neuroendocrinology. Gen. Comp. Endocrinol. 2012, 175, 357–366. [Google Scholar] [CrossRef]
- Revathi, P.; Iyapparaj, P.; Vasanthi, L.A.; Jeyanthi, S.; Krishnan, M. Impact of eyestalk ablation on the androgenic gland activity in the freshwater prawn Macrobrachium rosenbergii (De Man). World 2013, 5, 373–381. [Google Scholar]
- Treerattrakool, S.; Panyim, S.; Udomkit, A. Induction of ovarian maturation and spawning in Penaeus monodon broodstock by double-stranded RNA. Mar. Biotechnol. 2011, 13, 163–169. [Google Scholar] [CrossRef]
- Treerattrakool, S.; Chartthai, C.; Phromma-in, N.; Panyim, S.; Udomkit, A. Silencing of gonad-inhibiting hormone gene expression in Penaeus monodon by feeding with GIH dsRNA-enriched Artemia. Aquaculture 2013, 404, 116–121. [Google Scholar] [CrossRef]
- Jin, S.B.; Wang, N.; Qiao, H.; Fu, H.T.; Wu, Y.; Gong, Y.S.; Jiang, S.F.; Xiong, Y.W. Molecular cloning and expression of a full-length cDNA encoding crustacean hyperglycemic hormone (CHH) in oriental river pawn (Macrobrachium nipponense). J. Fish. China. 2013, 20, 82–92. [Google Scholar] [CrossRef]
- Qiao, H.; Xiong, Y.W.; Zhang, W.Y.; Fu, H.T.; Jiang, S.F.; Sun, S.M.; Bai, H.K.; Jin, S.B.; Gong, Y.S. Characterization, expression, and function analysis of gonad-inhibiting hormone in Oriental River prawn, Macrobrachium nipponense and its induced expression by temperature. Comp. Biochem. Physiol. A 2015, 185, 1–8. [Google Scholar] [CrossRef]
- Jin, S.B.; Zhang, W.Y.; Wang, P.C.; Jiang, S.F.; Qiao, H.; Gong, Y.; Wu, Y.; Xiong, Y.; Fu, H. Identification of potential functions of polo-like kinase 1 in male reproductive development of the oriental river prawn (Macrobrachium nipponense) by RNA interference analysis. Front. Endocrinol. 2022, 13, 1084802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Y.; Xiong, Y.W.; Wang, P.C.; Chen, T.Y.; Jiang, S.F.; Qiao, H.; Gong, Y.; Wu, Y.; Jin, S.B.; Fu, H.T. RNA interference analysis of potential functions of cyclin A in the reproductive development of male oriental river prawns (Macrobrachium nipponense). Front. Genet. 2022, 13, 1053826. [Google Scholar] [CrossRef]
- Zhang, W.Y.; Wang, P.C.; Xiong, Y.W.; Chen, T.Y.; Jiang, S.F.; Qiao, H.; Gong, Y.S.; Wu, Y.; Jin, S.B.; Fu, H.T. RNA interference analysis of the functions of Cyclin B in male reproductive development of the oriental river prawn (Macrobrachium nipponense). Genes 2022, 13, 2079. [Google Scholar] [CrossRef]
- Banerjee, S.K.; Weston, A.P.; Zoubine, M.N.; Campbell, D.R.; Cherian, R. Expression of cdc2 and cyclin B1 in Helicobacter pylori-associated gastric MALT and MALT lymphoma: Relationship to cell death, proliferation, and transformation. Am. J. Pathol. 2000, 156, 217–225. [Google Scholar] [CrossRef]
- Murray, A.; Hunt, T. The Cell Cycle: An Introduction; Oxford University Press: Oxford, UK, 1993. [Google Scholar]
- Bolsover, S.R.; Hyams, J.S.; Shephard, E.A.; White, H.A.; Wiedemann, C.G. Cell Biology: A Short Course, 2nd ed.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2004; pp. 408–415. [Google Scholar]
- Nurse, P. Universal control mechanism regulating onset of M-phase. Nature 1990, 344, 503–508. [Google Scholar] [CrossRef]
- Pines, J. Four-dimensional control of the cell cycle. Nat. Cell Biol. 1999, 1, E73–E79. [Google Scholar] [CrossRef]
- Murray, A.W.; Kirschner, M.W. Cyclin synthesis drives the early embryonic cell cycle. Nature 1989, 339, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Gallant, P.; Nigg, E.A. Identification of a novel vertebrate cyclin: Cyclin B3 shares properties with both A- and B-type cyclins. EMBO J. 1994, 13, 595–605. [Google Scholar] [CrossRef]
- Jacobs, H.W.; Knoblich, J.A.; Lehner, C.F. Drosophila cyclin B3 is required for female fertility and is dispensable for mitosis like cyclin B. Genes Dev. 1998, 12, 3741–3751. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.L. The Cloning and Functional Analysis of Cyclin B3 in Bombyx mori; Southwest University: Chongqing, China, 2009. [Google Scholar]
- Wang, T.; Li, L.; Kan, H.Y.; Zhang, G.F. Molecular cloning and characterization of the key regulator of cell cycle cyclin B3 in Pacific Oyster (Crassostrea gigas), and its role in gonad development. Mar. Sci. 2011, 12, 1–9. [Google Scholar]
- Karasu, M.E.; Keeney, S. Cyclin B3 is dispensable for mouse spermatogenesis. Chromosoma 2019, 128, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.B.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Sun, S.M.; Zhang, W.Y.; Gong, Y.S.; Fu, H.T. Molecular cloning of two tropomyosin family genes and expression analysis during development in oriental river prawn, Macrobrachium nipponense. Gene 2014, 546, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.B.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Sun, S.M.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Molecular cloning, expression, and in situ hybridization analysis of forkhead box protein L2 during development in Macrobrachium nipponense. J. World Aquacul. Soc. 2018, 49, 429–440. [Google Scholar] [CrossRef]
- Hu, Y.N.; Fu, H.T.; Qiao, H.; Sun, S.M.; Zhang, W.Y.; Jin, S.B.; Jiang, S.F.; Gong, Y.S.; Xiong, Y.W.; Wu, Y. Validation and evaluation of reference genes for Quantitative real-time PCR in Macrobrachium nipponense. Int. J. Mol. Sci. 2018, 19, 2258. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhang, S.B.; Jiang, P.; Wang, Z.Q.; Long, S.R.; Liu, R.D.; Zhang, X.; Yang, W.; Ren, H.J.; Cui, J. Dsrna-Mediated Silencing of Nudix Hydrolase in Trichinella Spiralis Inhibits the Larval Invasion and Survival in Mice. Exp. Parasitol. 2016, 162, 35–42. [Google Scholar] [CrossRef]
- Li, F.; Qiao, H.; Fu, H.T.; Sun, S.M.; Zhang, W.Y.; Jin, S.B.; Jiang, S.F.; Gong, Y.S.; Xiong, Y.W.; Wu, Y.; et al. Identification and characterization of opsin gene and its role in ovarian maturation in the oriental river prawn Macrobrachium nipponense. Comp. Biochem. Physiol. B 2018, 218, 1–12. [Google Scholar] [CrossRef]
- Jiang, F.W.; Fu, H.T.; Qiao, H.; Zhang, W.Y.; Jiang, S.F.; Xiong, Y.W.; Sun, S.M.; Gong, Y.S.; Jin, S.B. The RNA Interference Regularity of Transformer-2 Gene of Oriental River Prawn Macrobrachium nipponense. Chin. Agricul. Sci. Bul. 2014, 30, 32–37. [Google Scholar]
- Ma, X.K.; Liu, X.Z.; Wen, H.S.; Xu, Y.J.; Zhang, L.J. Histological observation on gonadal sex differentiation in Cynoglossus semilaevis Günther. Mar. Fish. Res. 2006, 27, 55–61. [Google Scholar]
- ShangGuan, B.M.; Liu, Z.Z.; Li, S.Q. Histological Studies on Ovarian Development in Scylla serrata. J. Fish. Sci. China 1991, 15, 96–103. [Google Scholar]
- Sigrist, S.; Jacobs, H.; Stratmann, R.; Lehner, C.F. Exit from mitosis is regulated by Drosophila fizzy and the sequential destruction of cyclins A, B and B3. EMBO J. 1995, 14, 4827–4838. [Google Scholar] [CrossRef] [PubMed]
- Lozano, J.C.; Perret, E.; Schatt, P.; Arnould, C.; Peaucellier, G.; Picard, A. Molecular Cloning, Gene Localization, and Structure of Human Cyclin B3. Biochem. Biophys. Res. Commun. 2002, 291, 406–413. [Google Scholar] [CrossRef]
- Kajiura-Kobayashi, H.; Kobayashi, T.; Nagahama, Y. The cloning of cyclin B3 and its gene expression during hormonally induced spermatogenesis in the teleost, Anguilla japonica. Biochem. Biophys. Res. Commun. 2004, 323, 288–292. [Google Scholar] [CrossRef]
- Jin, S.B.; Hu, Y.N.; Fu, H.T.; Sun, S.M.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Analysis of testis metabolome and transcriptome from the oriental river prawn (Macrobrachium nipponense) in response to different temperatures and illumination times. Comp. Biochem. Physiol. D 2020, 34, 100662. [Google Scholar] [CrossRef]
- Jin, S.B.; Zhang, W.Y.; Xiong, Y.W.; Jiang, S.F.; Qiao, H.; Gong, Y.S.; Wu, Y.; Fu, H.T. Genetic regulation of male sexual development in the oriental river prawn Macrobrachium nipponense during reproductive vs. non-reproductive season. Aquac. Int. 2022, 30, 2059–2079. [Google Scholar] [CrossRef]
- Jin, S.B.; Hu, Y.N.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. Identification and Characterization of the Pyruvate Dehydrogenase E1 Gene in the Oriental River Prawn, Macrobrachium nipponense. Front. Endocrinol. 2021, 12, 752501. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.B.; Fu, H.T.; Jiang, S.F.; Xiong, Y.W.; Qiao, H.; Zhang, W.Y.; Gong, Y.S.; Wu, Y. RNA Interference Analysis Reveals the Positive Regulatory Role of Ferritin in Testis Development in the Oriental River Prawn, Macrobrachium nipponense. Front. Physiol. 2022, 13, 805861. [Google Scholar] [CrossRef]
- Jin, S.B.; Zhang, W.Y.; Xiong, Y.W.; Fu, H.T. Recent progress of male sexual differentiation and development in the oriental river prawn (Macrobrachium nipponense): A review. Rev. Aquac. 2023, 15, 305–317. [Google Scholar] [CrossRef]
- Atsuro, O.; Yuriko, H.; Makoto, N.; Tsuyoshi, O.; Rinkei, K.; Masaaki, K.; Shogo, M.; Hiromichi, N. Preparation of an active recombinant peptide of crustacean androgenic gland hormone. Peptides 2002, 3, 567–572. [Google Scholar]
- Morakot, S.; Charoonroj, C.; Michael, J.S.; Nantawan, S.; Napamanee, K.; Ittipon, P.; Peter, J.H.; Prasert, S. Bilateral eyestalk ablation of the blue swimmer crab, Portunus pelagicus, produces hypertrophy of the androgenic gland and an increase of cells producing insulin-like androgenic gland hormone. Tissue Cell 2010, 5, 293–300. [Google Scholar]
- Sagi, A.; Cohen, D.; Milner, Y. Effect of androgenic gland ablation on morphotypic differentiation and sexual characteristics of male freshwater prawns Macrobrachium rosenbergii. Gen. Comp. Endocrinol. 1990, 77, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Sagi, A.; Cohen, D.; Wax, Y. Production of Macrobrachium rosenbetgii in momosex population: Yield characteristes under intensive monoculture conditions in cages. Aquaculture 1986, 51, 265–275. [Google Scholar] [CrossRef]
- Li, S.H.; Li, F.H.; Sun, Z.; Xiang, J.H. Two spliced variants of insulin-like androgenic gland hormone gene in the Chinese shrimp, Fenneropenaeus chinensis. Gen. Comp. Endocrinol. 2012, 177, 246–255. [Google Scholar] [CrossRef]
- Huang, X.S.; Ye, H.H.; Huang, H.Y.; Yang, Y.N.; Gong, J. An insulin-like androgenic gland hormone gene in the mud crab, Scylla paramamosain, extensively expressed and involved in the processes of growth and female reproduction. Gen. Comp. Endocrinol. 2014, 204, 229–238. [Google Scholar] [CrossRef]
- Liu, F.; Shi, W.; Ye, H.; Liu, A.; Zhu, Z. RNAi Reveals Role of Insulin-Like Androgenic Gland Hormone 2 (IAG2) in Sexual Differentiation and Growth in Hermaphrodite Shrimp. Front. Mar. Sci. 2021, 8, 666763. [Google Scholar] [CrossRef]
- Zhou, T.T.; Wang, W.; Wang, C.G.; Sun, C.B.; Shi, L.L.; Chan, S.F. Insulin-like Androgenic Gland Hormone from the Shrimp Fenneropenaeus merguiensis: Expression, Gene organization and Transcript variants. Gene 2021, 782, 145529. [Google Scholar] [CrossRef]
- Ma, K.Y.; Li, J.L.; Qiu, G.F. Identification of putative regulatory region of insulin-like androgenic gland hormone gene (IAG) in the prawn Macrobrachium nipponense and proteins that interact with IAG by using yeast two-hybrid system. Gen. Comp. Endocrinol. 2016, 229, 112–118. [Google Scholar] [CrossRef]
- Ventura, T.; Manor, R.; Aflalo, E.D.; Weil, S.; Rosen, O.; Sagi, A. Timing sexual differentiation: Full functional sex reversal achieved through silencing of a single insulin-like gene in the prawn, Macrobrachium rosenbergii. Biol. Reprod. 2012, 86, 90. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Nucleotide Sequence (5′→3′) | Purpose |
|---|---|---|
| CycB3-3GSP1 | ATGCCGACGACTTTCTTTATAT | FWD first primer for CycB3 3′ RACE |
| CycB3-3GSP2 | GCTGCAGCTGCTCTTTTCTTTA | FWD second primer for CycB3 3′ RACE |
| CycB3-5GSP1 | CTGTCGCACCTGAAGCTCAACC | RVS first primer for CycB3 5′ RACE |
| CycB3-5GSP2 | GTGAGGTCACCAAAAGCAGATC | RVS second primer for CycB3 5′ RACE |
| 3′RACE OUT | TACCGTCGTTCCACTAGTGATTT | RVS first primer for 3′ RACE |
| 3′RACE IN | CGCGGATCCTCCACTAGTGATTTCACTATAGG | RVS second primer for 3′ RACE |
| 5′RACE OUT | CATGGCTACATGCTGACAGCCTA | FWD first primer for 5′ RACE |
| 5′RACE IN | CGCGGATCCACAGCCTACTGATGATCAGTCGATG | FWD second primer for 5′ RACE |
| CycB3-RTF | GAAGGCGTTGACGATTATGACAG | FWD primer for CycB3 expression |
| CycB3-RTR | CTCATGCTTTTGGACACTTCAGG | RVS primer for CycB3 expression |
| IAG-RTF | CTGACCACACCTACTGAAGACAA | FWD primer for IAG expression |
| IAG-RTR | CGTTTTCGATAAGAGGTCAAGCC | RVS primer for IAG expression |
| EIF-F | CATGGATGTACCTGTGGTGAAAC | FWD primer for EIF expression |
| EIF-R | CTGTCAGCAGAAGGTCCTCATTA | RVS primer for EIF expression |
| CycB3 RNAi-F | TAATACGACTCACTATAGGGTCCGAGACACAACCAACAAA | FWD primer for RNAi analysis |
| CycB3 RNAi-R | TAATACGACTCACTATAGGGAGGAGGCTCTCACAAAACGA | RVS primer for RNAi analysis |
| Species | Accession Number |
|---|---|
| Macrobrachium nipponense | |
| Penaeus monodon | XP_037786045.1 |
| Penaeus japonicus | XP_042878469.1 |
| Procambarus clarkii | XP_045600532.1 |
| Homarus americanus | XP_042217784.1 |
| Penaeus vannamei | XP_027238877.1 |
| Portunus trituberculatus | XP_045137868.1 |
| Callinectes arcuatus | QPO25106.1 |
| Chionoecetes opilio | KAG0693500.1 |
| Hyalella azteca | XP_018006502.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jin, S.; Zhou, Z.; Zhang, W.; Xiong, Y.; Qiao, H.; Gong, Y.; Wu, Y.; Jiang, S.; Fu, H. RNAi Analysis of Potential Functions of Cyclin B3 in Reproduction of Male Oriental River Prawns (Macrobrachium nipponense). Animals 2023, 13, 1703. https://doi.org/10.3390/ani13101703
Jin S, Zhou Z, Zhang W, Xiong Y, Qiao H, Gong Y, Wu Y, Jiang S, Fu H. RNAi Analysis of Potential Functions of Cyclin B3 in Reproduction of Male Oriental River Prawns (Macrobrachium nipponense). Animals. 2023; 13(10):1703. https://doi.org/10.3390/ani13101703
Chicago/Turabian StyleJin, Shubo, Zhenyu Zhou, Wenyi Zhang, Yiwei Xiong, Hui Qiao, Yongsheng Gong, Yan Wu, Sufei Jiang, and Hongtuo Fu. 2023. "RNAi Analysis of Potential Functions of Cyclin B3 in Reproduction of Male Oriental River Prawns (Macrobrachium nipponense)" Animals 13, no. 10: 1703. https://doi.org/10.3390/ani13101703