

Effects of Melatonin and Silymarin on Reactive Oxygen Species, Nitric Oxide Production, and Sperm Viability and Motility during Sperm Freezing in Pigs

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal
2.2. Chemicals
2.3. Preparation of Semen
2.4. Freezing and Thawing of Semen
2.5. Evaluation of Sperm Motility
2.6. Sperm Viability Assay
2.7. Reactive Oxygen Species Determination
2.8. Production of Nitric Oxide
2.9. Statistical Analysis
3. Results
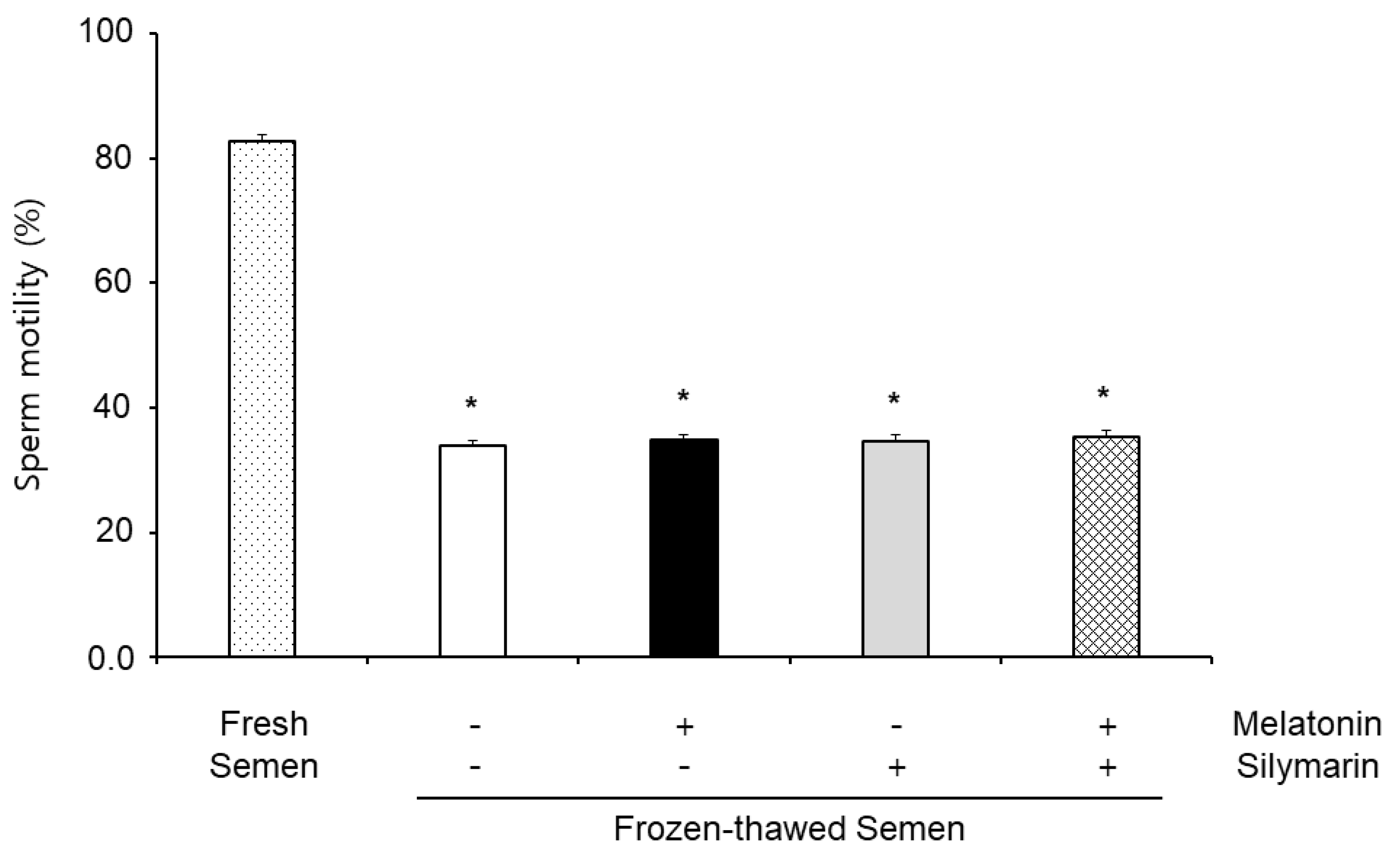
3.1. Effects of Melatonin and Silymarin on Sperm Motility
3.2. Dose-Dependent Effects of Melatonin and Silymarin on Sperm Viability
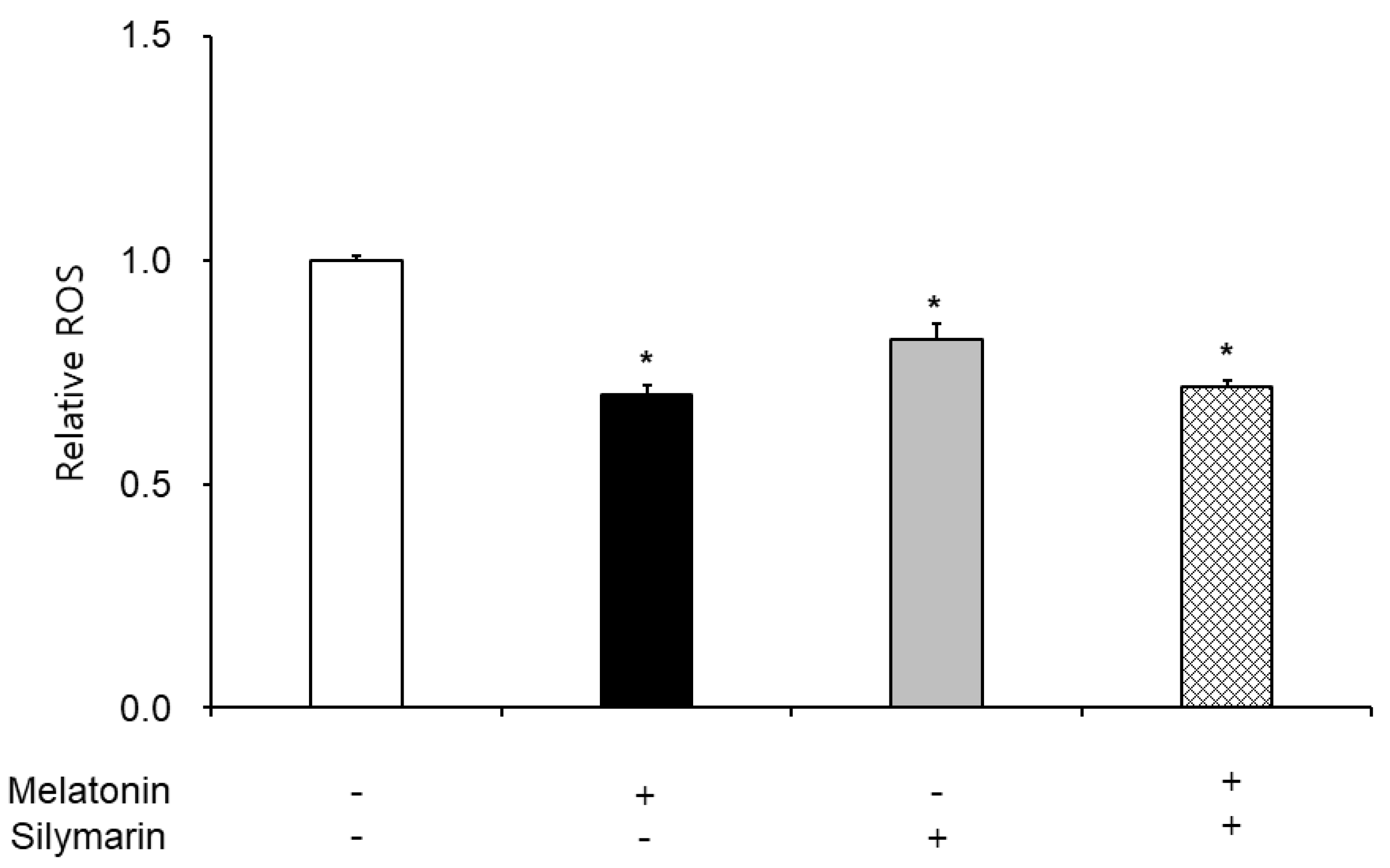
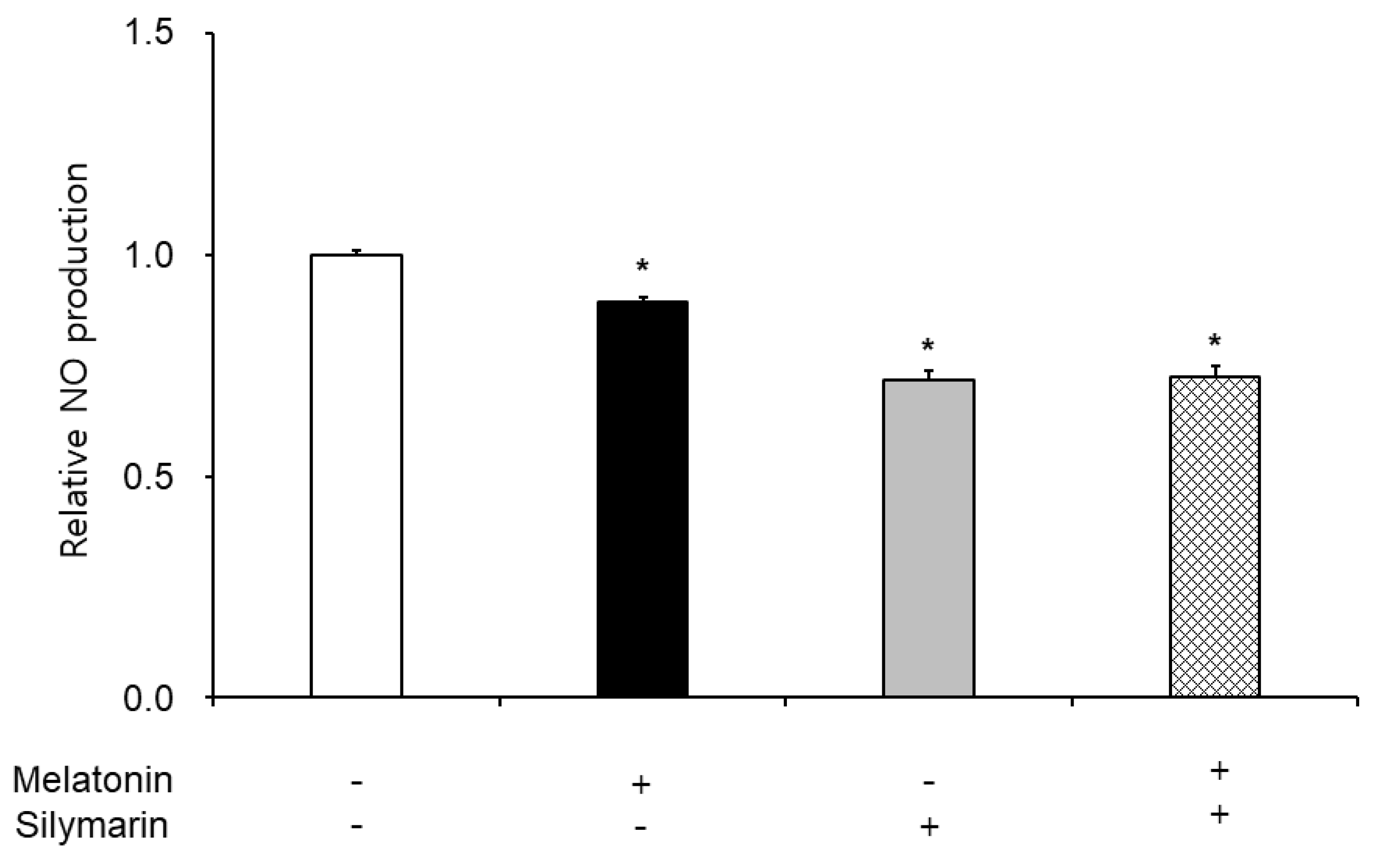
3.3. Effects of Melatonin and Silymarin on ROS and NO Production and Sperm Viability
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Agarwal, A.; Nallella, K.P.; Allamaneni, S.S.R.; Said, T.M. Role of antioxidants in treatment of male infertility: An overview of the literature. Reprod. Biomed. Online 2004, 8, 616–627. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2012, 10, 49. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Bucak, M.N.; Tuncer, P.B.; Sariozkan, S.; Baspinar, N.; Taspinar, M.; Coyan, K.; Bilgili, A.; Akalin, P.P.; Buyukleblebici, S.; Aydos, S.; et al. Effects of antioxidants on post-thawed bovine sperm and oxidative stress parameters: Antioxidants protect DNA integrity against cryodamage. Cryobiology 2010, 61, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E.; Mohey-Elsaeed, O.M. Evaluation of bull spermatozoa during and after cryopreservation: Structural and ultrastructural insights. Int. J. Vet. Sci. Med. 2018, 6, 49–56. [Google Scholar] [CrossRef]
- Agarwal, A.; Rosas, I.M.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative stress and assisted reproduction: A comprehensive review of its pathophysiological role and strategies for optimizing embryo culture environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef]
- Ford, W.C.L. Reactive oxygen species and sperm. Hum. Fertil. 2001, 4, 77–78. [Google Scholar] [CrossRef]
- Maiorino, M.; Ursini, F. Oxidative stress, Spermatogenesis and Fertility. Biol. Chem. 2002, 383, 591–597. [Google Scholar] [CrossRef]
- Yeste, M. Sperm cryopreservation update: Cryodamage, markers, and factors affecting the sperm freezability in pigs. Theriogenology 2017, 96, 246–256. [Google Scholar] [CrossRef]
- Koppers, A.J.; de Iuliis, G.N.; Finnie, J.M.; McLaughlin, E.A.; Aitken, R.J. Significance of mitochondrial reactive oxygen species in the generation of oxidative stress in spermatozoa. J. Clim. Endocrinol. Metab. 2008, 93, 3199–3207. [Google Scholar] [CrossRef]
- de Paz, P.; Muino-Blanco, T.; Alvarez, M.; Herrraez, P. Nitric oxide in mammalian spermatozoa: Its function in the regulation of sperm motility, capacitation, and acrosome reaction. Anim. Reprod. Sci. 2017, 176, 42–55. [Google Scholar]
- Kumar, S.; Kadirvel, G.; Kumaresan, A.; Anand, M.; Baithalu, R.K. Comparison of different antioxidants in semen extenders on post-thaw quality and oxidative status of buffalo spermatozoa. Reprod. Domest. Anim. 2020, 55, 293–302. [Google Scholar]
- Sieme, H.; Oldenhof, H.; Wolkers, W.F. Mode of action of cryoprotectants for sperm preservation. Anim. Reprod. Sci. 2016, 169, 2–5. [Google Scholar] [CrossRef]
- Ball, B.A.; Vo, A. Osmotic tolerance of equine spermatozoa and the effects of soluble cryoprotectants on equine sperm motility, viability, and mitochondrial membrane potential. J. Androl. 2001, 22, 1061–1069. [Google Scholar] [CrossRef]
- Mahfouz, R.Z.; du Plessis, S.S.; Aziz, N.; Sharma, R.; Sabanegh, E.; Agarwal, A. Sperm viability, apoptosis, and intracellular reactive oxygen species levels in human spermatozoa before and after induction of oxidative stress. Fertil. Steril. 2010, 93, 814–821. [Google Scholar] [CrossRef]
- Roca, J.; Rodríguez, M.J.; Gil, M.A.; Carvajal, G.; Garcia, E.M.; Cuello, C.; Vazquez, J.M.; Martinez, E.A. Survival and in vitro fertility of boar spermatozoa frozen in the presence of superoxide dismutase and/or catalase. J. Androl. 2005, 26, 15–24. [Google Scholar]
- Martin, G.; Sabido, O.; Durand, P.; Levy, R. Cryopreservation induces an apoptosis-like mechanism in bull sperm. Biol. Reprod. 2004, 71, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Ortega Ferrusola, C.; Gonzalez Fernandez, L.; Macias Garcia, B.; Salazar-Sandoval, C.; Morillo Rodriguez, A.; Rodríguez Martinez, H.; Tapia, J.; Pena, F. Effect of cryopreservation on nitric oxide production by stallion spermatozoa. Biol. Reprod. 2009, 81, 1106–1111. [Google Scholar] [CrossRef]
- Karimfar, M.; Niazvand, F.; Haghani, K.; Ghafourian, S.; Shirazi, R.; Bakhtiyari, S. The protective effects of melatonin against cryopreservation-induced oxidative stress in human sperm. Int. J. Immunopathol. Pharmacol. 2015, 28, 69–76. [Google Scholar] [CrossRef]
- Du Plessis, S.; Hagenaar, K.; Lampiao, F. The in vitro effects of melatonin on human sperm function and its scavenging activities on NO and ROS. Andrologia 2010, 42, 112–116. [Google Scholar] [CrossRef]
- Surai, P.F. Silymarin as a natural antioxidant: An overview of the current evidence and perspectives. Antioxidants 2015, 4, 204–247. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Tian, X.Z.; Kang, J.T.; Liang, X.W. Effects of oxidative stress on semen quality and fertilizing capacity of boar spermatozoa. J. Anim. Sci. Biotechnol. 2016, 7, 52. [Google Scholar]
- Tvrda, E.; Lukac, N. Oxidative stress, sperm damage and antioxidant strategies in the context of breeding management of boar. Anim. Reprod. Sci. 2021, 232, 106737. [Google Scholar]
- Pourmasumi, S.; Jafari-Joozani, R.; Talebi, A.R.; Deldar, H. Melatonin supplementation improves boar sperm viability, motility and DNA integrity during liquid storage at 17 °C. Theriogenology 2019, 130, 80–86. [Google Scholar]
- Yoon, S.J.; Rahman, M.S.; Kwon, W.S.; Park, Y.J.; Ryu, B.Y.; Pang, M.G.; Lee, W.C. Melatonin maintains mitochondrial membrane potential and decreases excessive intracellular reactive oxygen species levels in boar spermatozoa. J. Pineal Res. 2019, 67, e12590. [Google Scholar]
- Amini, A.; Gilanpour, H.; Zare-Shahneh, A.; Zeinoaldini, S. Melatonin improves the quality of cryopreserved boar semen by reducing oxidative stress. Theriogenology 2020, 144, 110–117. [Google Scholar]
- Wang, X.; Liu, X.; Zhang, L.; Xu, L.; Zhang, Y.; Hu, J. Protective effects of silymarin on boar sperm quality and fertility after freezing-thawing. Cryobiology 2018, 80, 89–95. [Google Scholar]
- Tuncer, P.B.; Kilic, N.; Oktem, F.; Ozguner, F. The effects of silymarin on oxidative stress and DNA damage in boar semen after cryopreservation. Andrologia 2019, 51, e13338. [Google Scholar]
- Gadea, J. Semen extenders used in the artificial insemination of swine. Span. J. Agric. Res. 2003, 1, 17–27. [Google Scholar] [CrossRef]
- Kim, Y.J.; Lee, S.H.; Lee, Y.J.; Oh, H.I.; Cheong, H.T.; Yang, B.K.; Lee, S.; Park, C.K. Effect of nicotinic acid on sperm characteristic and oocyte development after in vitro fertilization using cryopreserved boar semen. J. Emb. Trans. 2015, 30, 7–15. [Google Scholar] [CrossRef]
- Lee, Y.S.; Lee, S.; Lee, S.H.; Yang, B.K.; Park, C.K. Effect of cholesterol-loaded-cyclodextrin on sperm viability and acrosome reaction in boar semen cryopreservation. Anim. Reprod. Sci. 2015, 159, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.J.; Jeong, Y.J.; Kim, J.; Jung, E.; Jun, J.H. Soybean peptides induce apoptosis in HeLa cells by increasing oxidative stress. Biomed. Sci. Lett. 2015, 21, 77–83. [Google Scholar] [CrossRef]
- Kojima, H.; Nakatsubo, N.; Kikuchi, K.; Kawahara, S.; Kirino, Y.; Nagoshi, H.; Hirata, Y.; Nagano, T. Detection and imaging of nitric oxide with novel fluorescent indicators: Diaminofluoresceins. Anal. Chem. 1998, 70, 2446–2453. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin protects ram spermatozoa from cryopreservation injuries in a dose-dependent manner. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Melchiorri, D.; Sewerynek, E.; Poeggeler, B.; Barlow-Walden, L.; Chuang, J.; Ortiz, G.G.; Acuña Castroviejo, D. A review of the evidence supporting melatonin’s role as an antioxidant. J. Pineal Res. 1995, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhao, X.; Zhang, H.; Li, Q.; Lu, G.; Zhao, X. Modulatory effect of silymarin on pulmonary vascular dysfunction through HIF-1α-iNOS following rat lung ischemia-reperfusion injury. Exp. Ther. Med. 2016, 12, 1135–1140. [Google Scholar] [CrossRef]
- Katiyar, S.K. Silymarin and skin cancer prevention: Anti-inflammatory, antioxidant and immunomodulatory effects. Int. J. Oncol. 2005, 26, 169–176. [Google Scholar] [CrossRef]
- Zhao, J.; Lahiri-Chatterjee, M.; Sharma, Y.; Agarwal, R. Inhibitory effect of a flavonoid antioxidant silymarin on benzoyl peroxide-induced tumor promotion, oxidative stress and inflammatory responses in SENCAR mouse skin. Carcinogenesis 2000, 21, 811–816. [Google Scholar] [CrossRef]
- Etemadi, T.; Momeni, H.R.; Ghafarizadeh, A.A. Impact of silymarin on cadmium-induced apoptosis in human spermatozoa. Andrologia 2020, 52, e13795. [Google Scholar] [CrossRef]
- Ognjanović, B.I.; Marković, S.D.; Ðorđević, N.Z.; Trbojević, I.S.; Štajn, A.Š.; Saičić, Z.S. Cadmium-induced lipid peroxidation and changes in antioxidant defense system in the rat testes: Protective role of coenzyme Q10 and Vitamin E. Reprod. Toxicol. 2010, 29, 191–197. [Google Scholar] [CrossRef]
- Abedi, H.; Jahromi, H.K.; Hashemi, S.M.A.; Jashni, H.K.; Jahromi, Z.K.; Pourahmadi, M. The effect of silymarin on spermatogenesis process in rats. Int. J. Med. Res. Health Sci. 2016, 5, 146–150. [Google Scholar]
- Jang, H.; Park, I.; Yuh, I.; Cheong, H.; Kim, J.; Park, C.; Yang, B. Beneficial effects of silymarin against nitric oxide-induced oxidative stress on cell characteristics of bovine oviduct epithelial cell and developmental ability of bovine IVF embryos. J. Appl. Anim. Res. 2014, 42, 166–176. [Google Scholar] [CrossRef]
- Stuehr, D.J. Mammalian nitric oxide synthases. Biochim. Biophys. Acta-Bioenerg. 1999, 1411, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.S.; Kim, Y.M.; Wang, Q.; Shapiro, R.A.; Billiar, T.R.; Geller, D.A. Nitric oxide down-regulates hepatocyte–inducible nitric oxide synthase gene expression. Arch. Surg. 1997, 132, 1177–1183. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.J.; Kwon, Y.G.; Chung, H.T.; Lee, S.K.; Simmons, R.L.; Billiar, T.R.; Kim, Y.M. Antioxidant enzymes suppress nitric oxide production through the inhibition of NF-κB activation: Role of H2O2 and nitric oxide in inducible nitric oxide synthase expression in macrophages. Nitric Oxide-Biol.Chem. 2001, 5, 504–513. [Google Scholar] [CrossRef]
- Salerno, L.; Modica, M.N.; Romeo, G.; Pittalà, V.; Siracusa, M.A.; Amato, M.E.; Acquaviva, R.; Di Giacomo, C.; Sorrenti, V. Novel inhibitors of nitric oxide synthase with antioxidant properties. Eur. J. Med. Chem. 2012, 49, 118–126. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Company | Cat. # | Concentration (g/L) | |
|---|---|---|---|---|
| Modena 1 | Glucose | Sigma | G8270 | 25.0 |
| EDTA | E5134 | 2.25 | ||
| Sodium citrate | S4641 | 6.90 | ||
| Sodium bicarbonate | S5761 | 1.00 | ||
| Tris | T6066 | 5.65 | ||
| Citrate | C1909 | 2.00 | ||
| Cysteine | 7880 | 0.05 | ||
| BSA 2 | A4503 | 3.00 | ||
| Gentamicin sulfate | Sigma | G3632 | 0.30 | |
| pH | - | - | 7.00 |
| Melatonin (mM) | ||||
|---|---|---|---|---|
| Viability | 0 | 0.01 | 0.1 | 1.0 |
| 41.71 ± 0.84 | 42.72 ± 0.67 | 45.52 ± 1.07 * | 39.17 ± 0.45 | |
| Silymarin (mM) | ||||
|---|---|---|---|---|
| Viability | 0 | 0.001 | 0.01 | 0.1 |
| 39.31 ± 0.60 | 41.33 ± 1.04 | 45.86 ± 1.24 * | 37.75 ± 1.85 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Lee, S. Effects of Melatonin and Silymarin on Reactive Oxygen Species, Nitric Oxide Production, and Sperm Viability and Motility during Sperm Freezing in Pigs. Animals 2023, 13, 1705. https://doi.org/10.3390/ani13101705
Lee S-H, Lee S. Effects of Melatonin and Silymarin on Reactive Oxygen Species, Nitric Oxide Production, and Sperm Viability and Motility during Sperm Freezing in Pigs. Animals. 2023; 13(10):1705. https://doi.org/10.3390/ani13101705
Chicago/Turabian StyleLee, Sang-Hee, and Seunghyung Lee. 2023. "Effects of Melatonin and Silymarin on Reactive Oxygen Species, Nitric Oxide Production, and Sperm Viability and Motility during Sperm Freezing in Pigs" Animals 13, no. 10: 1705. https://doi.org/10.3390/ani13101705