
Effects of Bee Pollen on Growth Performance, Intestinal Microbiota and Histomorphometry in African Catfish
 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Research Material and Studied Animals
2.2. Feed Preparation
2.3. Experimental Design, Fish Management, and Sampling Procedures
2.4. Rearing Indicators and Equations
2.5. Origin of Bee Pollen
2.6. The Chemical Composition of BP used in the Present Study
2.6.1. Extract BP Analysis
2.6.2. Determination of the Total Antioxidant Capacity (TAC)
2.6.3. Determination of Total Flavonoid Content (TFC)
2.6.4. Determination of Total Phenolic Content (TPC)
2.7. Chemical Analysis of Elements found in the Fish Body
2.8. Intestinal Microbiota
2.9. Histological Analysis
2.10. Statistical Analysis
3. Results
3.1. Effects of BP on the Fish Rearing Indicators
3.2. Analysis of TAC, TFC and TPC of BP
3.3. The Elemental Composition of the Fish Muscles
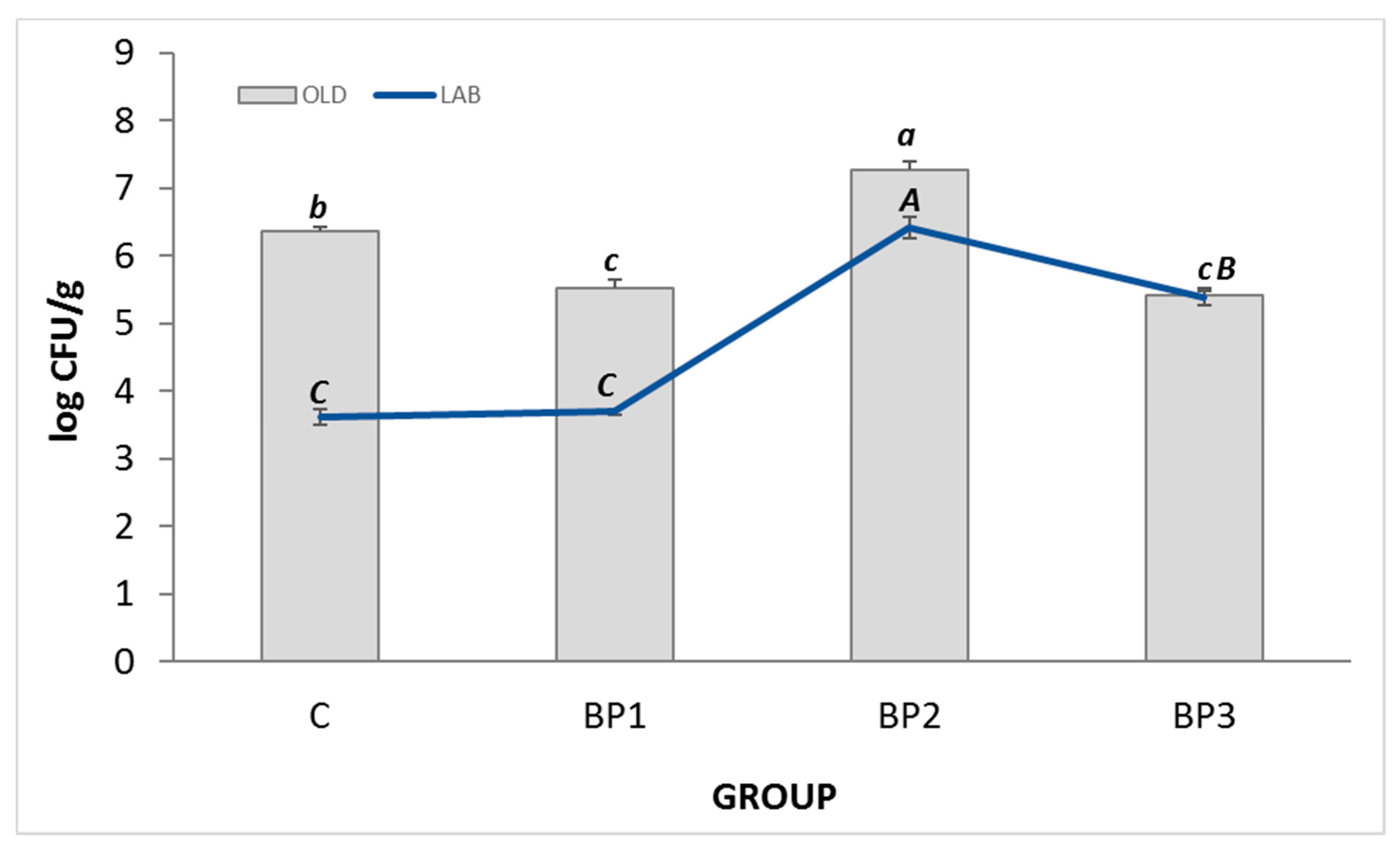
3.4. Intestinal Microflora
3.5. Histological Analysis
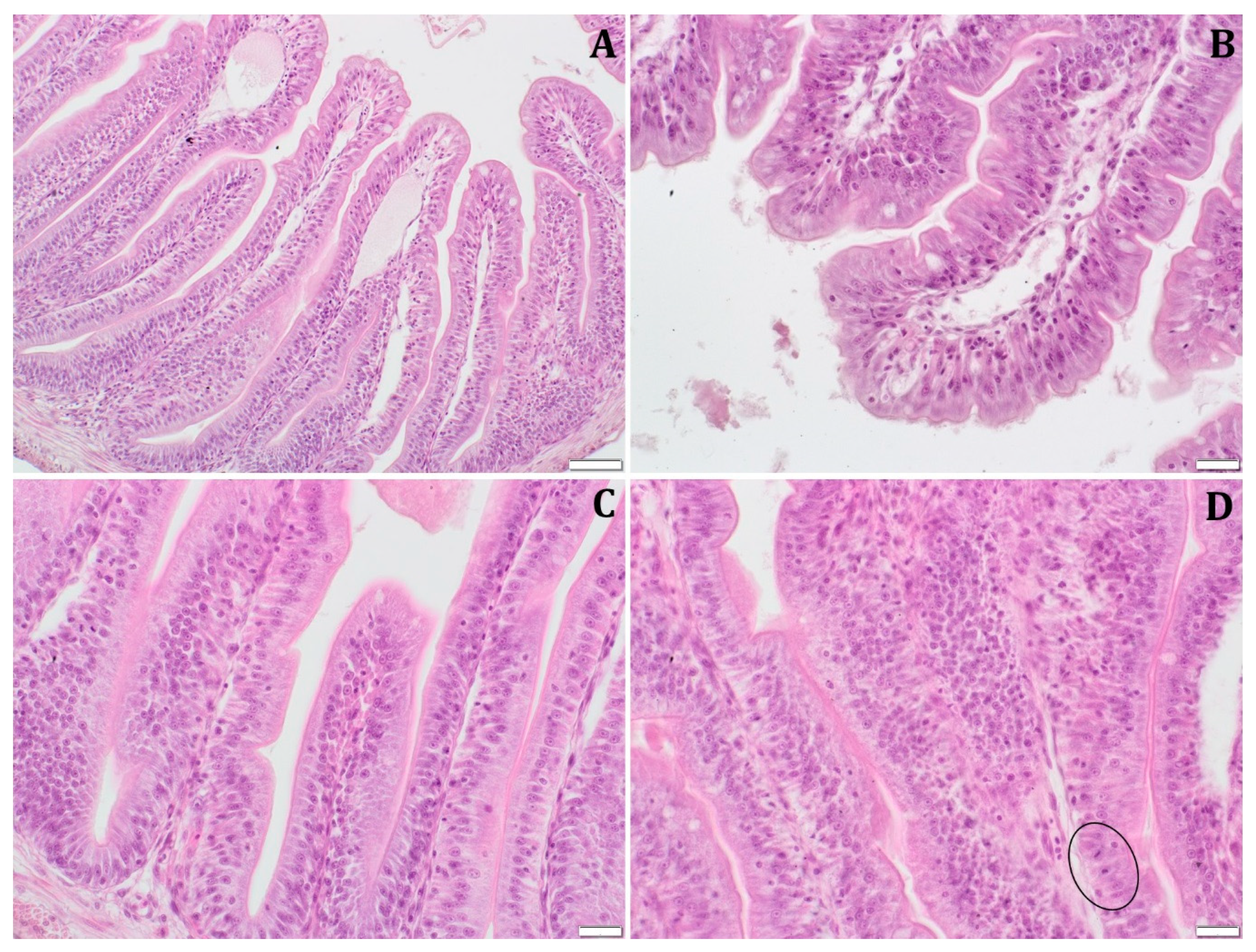
3.5.1. Intestinal Histomorphology and Morphometry
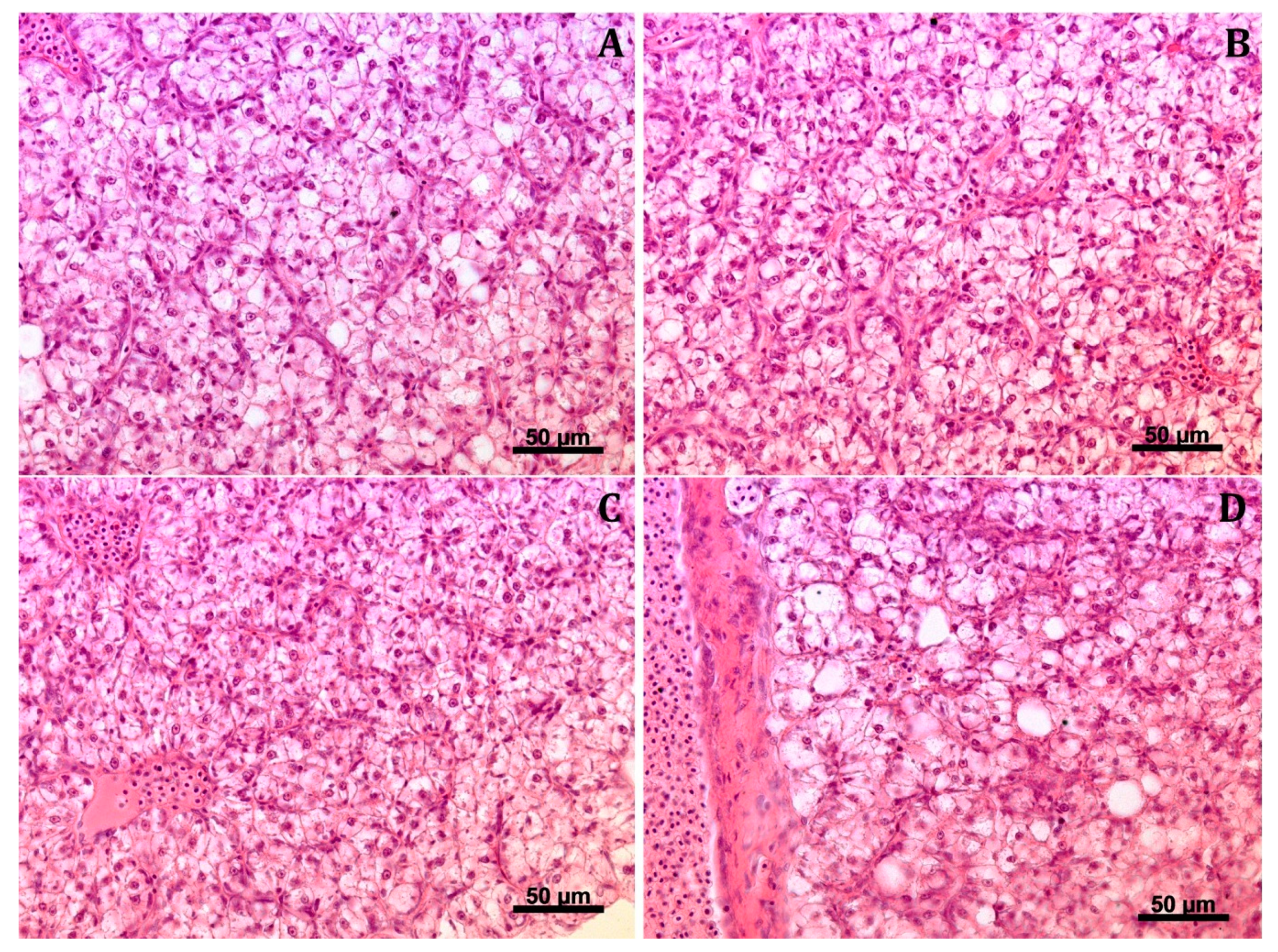
3.5.2. Hepatic Histoarchitecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Buschmann, A.H.; Bush, S.R.; Cao, L.; Klinger, D.H.; Little, D.C.; Lubchenco, J.; Shumway, S.E.; Troell, M. A 20-year retrospective review of global aquaculture. Nature 2021, 591, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, D.; Kucharczyk, D.J.; Nowosad, J.; Omirzhanova, N. Optimization of artificial insemination outcomes of African catfish (Clarias gariepinus) with differing hatchery conditions. Anim. Reprod. Sci. 2019, 211, 106222. [Google Scholar] [CrossRef] [PubMed]
- Dauda, A.B.; Natrah, I.; Karim, M.; Kamarudin, M.S.; Bichi, A. African catfish aquaculture in Malaysia and Nigeria: Status, trends and prospects. Fish. Aquac. J. 2018, 9, 1–5. [Google Scholar] [CrossRef]
- Onwuteaka, J.; Prince, C.O. Cannibalism of juvenile catfish (Clarias gariepinus Burchell). J. Enviromental Sci. Toxicol. Food Technol. 2015, 9, 46–49. [Google Scholar]
- Rosa, R.; Bandarra, N.M.; Nunes, M.L. Nutritional quality of African catfish Clarias gariepinus (Burchell 1822): A positive criterion for the future development of the European production of Siluroidei. Int. J. Food Sci. Technol. 2007, 42, 342–351. [Google Scholar] [CrossRef]
- Paleckaitis, M.; Buckiūnienė, V.; Trepenaitienė, R.; Klementaviciūtė, J.; Racevičiūtė-Stupelienė, A.; Šašytė, V.; Kudlinskienė, I. Effect of different extruded compound feed on African catfish (Clarias gariepinus) productivity, meat chemical and technological parameters. Vet. Zootech. 2018, 76, 66–70. [Google Scholar]
- Belão, T.C.; Leite, C.A.C.; Florindo, L.H.; Kalinin, A.L.; Rantin, F.T. Cardiorespiratory responses to hypoxia in the African catfish, Clarias gariepinus (Burchell 1822), an air-breathing fish. J. Comp. Physiol. B 2011, 181, 905–916. [Google Scholar] [CrossRef]
- Păpuc, T.; Petrescu-Mag, I.V.; Gavriloaie, C.; Botha, M.; Kovacs, E.; Coroian, C.O. Swimming in the mud-a short review of environmental parameter ranges tolerated by Clarias gariepinus. Extrem. Life Biospeology Astrobiol. 2019, 11, 9–17. [Google Scholar]
- van de Nieuwegiessen, P.G.; Olwo, J.; Khong, S.; Verreth, J.A.J.; Schrama, J.W. Effects of age and stocking density on the welfare of African catfish, Clarias gariepinus Burchell. Aquaculture 2009, 288, 69–75. [Google Scholar] [CrossRef]
- Abraham, T.J.; Mallick, P.K.; Paul, P. African catfish Clarias gariepinus farming practices in North and South 24 Parganas districts of West Bengal, India. J. Fish. 2018, 6, 579–586. [Google Scholar] [CrossRef]
- Besson, M.; Komen, H.; Aubin, J.; de Boer, I.J.M.; Poelman, M.; Quillet, E.; Vancoillie, C.; Vandeputte, M.; van Arendonk, J.A.M. Economic values of growth and feed efficiency for fish farming in recirculating aquaculture system with density and nitrogen output limitations: A case study with African catfish (Clarias gariepinus). J. Anim. Sci. 2014, 92, 5394–5405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rola, W.R.; Hasan, M.R. Economics of aquaculture feeding practices: A synthesis of case studies undertaken in six Asian countries. FAO Fish. Tech. Pap. 2007, 505, 1. [Google Scholar]
- Van Weerd, J.H. Nutrition and growth in Clarias species-a review. Aquat. Living Resour. 1995, 8, 395–401. [Google Scholar] [CrossRef]
- Ajiboye, O.; Yakubu, A.; Adams, T. A perspective on the ingestion and nutritional effects of feed additives in farmed fish species. World J. Fish Mar. Sci. 2012, 4, 87–101. [Google Scholar]
- Al-Hafedh, Y.S.; Ali, S.A. Effects of feeding on survival, cannibalism, growth and feed conversion of African catfish, Clarias gariepinus (Burchell) in concrete tanks. J. Appl. Ichthyol. 2004, 20, 225–227. [Google Scholar] [CrossRef]
- Indriastuti, C.E.; Junior, M.Z.; Suprayudi, M.A.; Supriyono, E.; Alimuddin, A. Cannibalism, survival and growth performance of juvenile African catfish Clarias gariepinus in relation to photoperiod and 17β-Oestradiol treatment. Aquac. Res. 2022, 53, 4437–4448. [Google Scholar] [CrossRef]
- Ukwe, O.I.K.; Edun, O.M.; Akinrotimi, O.A. Growth and Microbial Indices in African Catfish (Clarias gariepinus) Larvae Fed Formulated and Commercial Diets. J. Fish. Sci. 2018, 12, 1–8. [Google Scholar] [CrossRef]
- Kawamura, G.; Lim, J.X.; Ching, F.F.; Mustafa, S.; Lim, L.-S. Possible sensory control of cannibalism in the African catfish (Clarias gariepinus) larvae by electrical ablation of electroreceptors. Aquaculture 2021, 542, 73687. [Google Scholar] [CrossRef]
- Demir, O.; Türker, A.; Acar, U.; Kesbiç, S.O. Effects of Dietary Fish Oil Replacement by Unrefined Peanut Oil on the Growth, Serum Biochemical and Hematological Parameters of Mozambique Tilapia Juveniles (Oreochromis mossambicus). Turk. J. Fish. Aquat. Sci. 2014, 14, 887–892. [Google Scholar] [CrossRef]
- Yılmaz, S.; Acar, U.; Kesbiç, S.O.; Gültepe, N.; Ergün, S. Effects of dietary allspice, Pimenta dioica powder on physiological responses of Oreochromis mossambicus under low pH stress. Springer Plus 2015, 4, 719. [Google Scholar] [CrossRef] [Green Version]
- Farag, M.R.; Abdelnour, S.A.; Patra, A.K.; Dhama, K.; Dawood, M.A.O.; Elnesr, S.S. AlagawanyPropolis: Properties and composition, health benefits and applications in fish nutrition. Fish Shellfish Immunol. 2021, 115, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Di Chiacchio, I.M.; Gómez-Abenza, E.; Paiva, I.M.; de Abreu, D.J.M.; Rodríguez-Vidal, J.F.; Carvalho, E.E.N.; Carvalho, S.M.; Solis-Murgas, L.D.; Mulero, V. Bee pollen in zebrafish diet affects intestinal microbiota composition and skin cutaneous melanoma development. Sci. Rep. 2022, 12, 9998. [Google Scholar] [CrossRef] [PubMed]
- Messina, C.M.; Panettieri, V.; Arena, R.; Renda, G.; Espinosa Ruiz, C.; Morghese, M.; Piccolo, G.; Santulli, A.; Bovera, F. The Inclusion of a Supercritical Fluid Extract, Obtained From Honey Bee Pollen, in the Diet of Gilthead Sea Bream (Sparus aurata), Improves Fish Immune Response by Enhancing Anti-oxidant, and Anti-bacterial Activities. Front. Vet. Sci. 2020, 7, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Chiacchio, I.M.; Paiva, I.M.; de Abreu, D.J.M.; Carvalho, E.E.N.; Martínez, P.J.; Carvalho, S.M.; Mulero, V.; Murgas, L.D.S. Bee pollen as a dietary supplement for fish: Effect on the reproductive performance of zebrafish and the immunological response of their offspring. Fish Shellfish Immunol. 2021, 119, 300–307. [Google Scholar] [CrossRef]
- Acar, Ü.; Kesbiç, O.S.; Yılmaz, S.; Inanan, B.E.; Zemheri-Navruz, F.; Terzi, F.; Fazio, F.; Parrino, V. Effects of Essential Oil Derived from the Bitter Orange (Citrus aurantium) on Growth Performance, Histology and Gene Expression Levels in Common Carp Juveniles (Cyprinus carpio). Animals 2021, 11, 1431. [Google Scholar] [CrossRef]
- Arena, R.; de Medeiros, A.C.L.; Secci, G.; Mancini, S.; Manuguerra, S.; Bovera, F.; Santulli, A.; Parisi, G.; Messina, C.M.; Piccolo, G. Effects of Dietary Supplementation with Honeybee Pollen and Its Supercritical Fluid Extract on Immune Response and Fillet’s Quality of Farmed Gilthead Seabream (Sparus aurata). Animals 2022, 12, 675. [Google Scholar] [CrossRef]
- Thorp, R.W. The collection of pollen by bees. In Pollen and Pollination; Dafni, A., Hesse, M., Pacini, E., Eds.; Springer: Vienna, Austria, 2000; pp. 211–223. [Google Scholar]
- Ahmadifar, E.; Yousefi, M.; Karimi, M.; Fadaei Raieni, R.; Dadar, M.; Yilmaz, S.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Benefits of Dietary Polyphenols and Polyphenol-Rich Additives to Aquatic Animal Health: An Overview. Rev. Fish. Sci. Aquac. 2021, 29, 478–511. [Google Scholar] [CrossRef]
- Campos, M.G.; Webby, R.F.; Markham, K.R.; Mitchell, K.A.; da Cunha, A.P. Age-Induced Diminution of Free Radical Scavenging Capacity in Bee Pollens and the Contribution of Constituent Flavonoids. J. Agric. Food Chem. 2003, 51, 742–745. [Google Scholar] [CrossRef] [Green Version]
- Campos, M.G.R.; Bogdanov, S.; de Almeida-Muradian, L.B.; Szczesna, T.; Mancebo, Y.; Frigerio, C.; Ferreira, F. Pollen composition and standardisation of analytical methods. J. Apic. Res. 2008, 47, 154–161. [Google Scholar] [CrossRef]
- Thakur, M.; Nanda, V. Composition and functionality of bee pollen: A review. Trends Food Sci. Technol. 2020, 98, 82–106. [Google Scholar] [CrossRef]
- Yang, K.; Wu, D.; Ye, X.; Liu, D.; Chen, J.; Sun, P. Characterization of Chemical Composition of Bee Pollen in China. J. Agric. Food Chem. 2013, 61, 708–718. [Google Scholar] [CrossRef] [PubMed]
- Abbass, A.A.; El-Asely, A.M.; Kandiel, M.M. Effects of dietary propolis and pollen on growth performance, fecundity and some hematological parameters of Oreochromis niloticus. Turk. J. Fish. Aquat. Sci. 2012, 12, 851–859. [Google Scholar]
- Abdel-Hamid, T.M.; El-Tarabany, M.S. Effect of bee pollen on growth performance, carcass traits, blood parameters, and the levels of metabolic hormones in New Zealand White and Rex rabbits. Trop. Anim. Health Prod. 2019, 51, 2421–2429. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Abd El-Hack, M.E.; Alagawany, M.; Farag, M.R.; Elnesr, S.S. Beneficial impacts of bee pollen in animal production, reproduction and health. J. Anim. Physiol. Anim. Nutr. 2019, 103, 477–484. [Google Scholar] [CrossRef]
- El-Asely, A.M.; Abbass, A.A.; Austin, B. Honey bee pollen improves growth, immunity and protection of Nile tilapia (Oreochromis niloticus) against infection with Aeromonas hydrophila. Fish Shellfish Immunol. 2014, 40, 500–506. [Google Scholar] [CrossRef]
- Choobkar, N.; Kakoolaki, S.; Mohammadi, F.; Rezaeimanesh, M. The effect of dietary propolis and pollen extracts on growth performance and haematological responses of rainbow trout (Onchorhynchus mykiss). Iran. J. Aquat. Anim. Health 2017, 3, 16–25. [Google Scholar] [CrossRef] [Green Version]
- Asgari, M.; Kenari, A.A.; Esmaeili, M.; Rombenso, R. Effects of hydroalcoholic extract of honeybee pollen on growth performance, flesh quality, and immune and stress response response of rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2020, 26, 1505–1519. [Google Scholar] [CrossRef]
- Panettieri, V.; Chatzifotis, S.; Messina, C.M.; Olivotto, I.; Manuguerra, S.; Randazzo, B.; Ariano, A.; Bovera, F.; Santulli, A.; Severino, L.; et al. Honey Bee Pollen in Meagre (Argyrosomus regius) Juvenile Diets: Effects on Growth, Diet Digestibility, Intestinal Traits, and Biochemical Markers Related to Health and Stress. Animals 2020, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Latif, H.M.R.; Shukry, M.; Saad, M.F.; Mohamed, N.A.; Nowosad, J.; Kucharczyk, D. Effects of GnRHa and hCG with or without dopamine receptor antagonists on the spawning efficiency of African catfish (Clarias gariepinus) reared in hatchery conditions. Anim. Reprod. Sci. 2021, 231, 106798. [Google Scholar] [CrossRef] [PubMed]
- Kucska, B.; Quyen, N.N.; Szabó, T.; Gebremichael, A.; Alebachew, G.W.; Bógó, B.; Horváth, L.; Csorbai, B.; Urbányi, B.; Kucharczyk, D.; et al. The effects of different hormone administration methods on propagation successes in African catfish (Clarias gariepinus). Aquaculture Reports 2022, 26, 101311. [Google Scholar] [CrossRef]
- Sikora, M.; Nowosad, J.; Biegaj, M.; Kucharczyk, D.; Dębowski, M. The possibility of application of agglomerate elastomers (EPP) as media for biological bed in aquaculture. Aquac. Res. 2018, 49, 2988–2994. [Google Scholar] [CrossRef]
- Kesbic, O.S.; Acar, U.; Yigit, M.; Bulut, M.; Gultepe, N.; Yilmaz, S. Unrefined Peanut Oil as a Lipid Source in Diets for Juveniles of Two-banded Seabream Diplodus vulgaris. N. Am. J. Aquac. 2016, 78, 64–71. [Google Scholar] [CrossRef]
- Lawag, I.L.; Yoo, O.; Lim, L.Y.; Hammer, K.; Locher, C. Optimisation of Bee Pollen Extraction to Maximise Extractable Antioxidant Constituents. Antioxidants 2021, 10, 1113. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, D.; Turemko, M. Electroactive Phenolic Contributors and Antioxidant Capacity of Flesh and Peel of 11 Apple Cultivars Measured by Cyclic Voltammetry and HPLC–DAD–MS/MS. Antioxidants 2020, 9, 1054. [Google Scholar] [CrossRef] [PubMed]
- Kühlwein, H.; Emery, M.J.; Rawling, M.D.; Harper, G.M.; Merrifield, D.L.; Davies, S.J. Effects of a dietary β-(1,3)(1,6)-D-glucan supplementation on intestinal microbial communities and intestinal ultrastructure of mirror carp (Cyprinus carpio L.). J. Appl. Microbiol. 2013, 115, 1091–1106. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. The hematoxylin and eosin. In Theory and Practice of Histological Techniques, 7th ed.; Suvarna, S., Layton, C., Bancroft, J., Eds.; Churchill Livingstone: Edinburgh, UK; New York, NY, USA, 2013; pp. 179–220. [Google Scholar]
- Abdel-Latif, H.M.R.; Abdel-Daim, M.M.; Shukry, M.; Nowosad, J.; Kucharczyk, D. Benefits and applications of Moringa oleifera as a plant protein source in Aquafeed: A review. Aquaculture 2022, 547, 737369. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Shukry, M.; Noreldin, A.E.; Ahmed, H.A.; El-Bahrawy, A.; Ghetas, H.A.; Khalifa, E. Milk thistle (Silybum marianum) extract improves growth, immunity, serum biochemical indices, antioxidant state, hepatic histoarchitecture, and intestinal histomorphometry of striped catfish, Pangasianodon hypophthalmus. Aquaculture 2023, 562, 738761. [Google Scholar] [CrossRef]
- Picoli, F.; Lopes, D.L.d.A.; Zampar, A.; Serafini, S.; Freccia, A.; Veronezi, L.O.; Kowalski, M.W.; Ghizzo, J.B.; Emerenciano, M.G.C. Dietary bee pollen affects hepatic–intestinal histomorphometry of Nile tilapia fingerlings. Aquac. Res. 2019, 50, 3295–3304. [Google Scholar] [CrossRef]
- Prusińska, M.; Nowosad, J.; Jarmołowicz, S.; Mikiewicz, M.; Duda, A.; Wiszniewski, G.; Sikora, M.; Biegaj, M.; Samselska, A.; Arciuch-Rutkowska, M.; et al. Effect of feeding barbel larvae (Barbus barbus (L, 1758)) Artemia sp. nauplii enriched with PUFAs on their growth and survival rate, blood composition, alimentary tract histological structure and body chemical composition. Aquac. Rep. 2020, 18, 100492. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 737951. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Yilmaz, E.; Dawood, M.A.O.; Ringø, E.; Ahmadifar, E.; Yilmaz, S. Shrimp vibriosis and possible control measures using probiotics, postbiotics, prebiotics, and synbiotics: A review. Aquaculture 2022, 551, 737951. [Google Scholar] [CrossRef]
- Oluyemi, K.G. Histometry and Growth Performance of African Catfish, Clarias gariepinus (Burchell, 1822) Fed Probiotics Supplemented Diets. Asian J. Res. Zool. 2019, 2, 1–10. [Google Scholar] [CrossRef]
- Yilmaz, S.; Yilmaz, E.; Dawood, M.A.O.; Ringø, E.; Ahmadifar, E.; Abdel-Latif, H.M.R. Probiotics, prebiotics, and synbiotics used to control vibriosis in fish: A review. Aquaculture 2022, 547, 737514. [Google Scholar] [CrossRef]
- Denisow, B.; Denisow-Pietrzyk, M. Biological and therapeutic properties of bee pollen: A review. J. Sci. Food Agric. 2016, 96, 4303–4309. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Wang, Q.; Xin, B.; Wang, H. Trophic effect of bee pollen on small intestine in broiler chickens. Journal of Medicinal Food 2007, 10, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Hajková, Z.; Toman, R.; Hluchý, S.; Gálik, B.; Bíro, D.; Martiniaková, M.; Omelka, R.; Boboňová, I. The effect of pollen on the structure of the small intestine in rats after an experimental addition in diet. Anim. Sci. and Bio 2013, 46, 232–237. [Google Scholar]
- Attia, Y.A.; Al-Hanoun, A.; Tag El- Din, A.E.; Bovera, F.; Shewika, Y.E. Effect of bee pollen levels on productive, reproductive and blood traits of NZW rabbits. J. Anim. Physiol. Anim. Nutr. 2011, 95, 294–303. [Google Scholar] [CrossRef]
- Baker, R.T.M.; Davies, S.J. Modulation of tissue α-tocopherol in African catfish, Clarias gariepinus (Burchell), fed oxidized oils, and the compensatory effect of supplemental dietary vitamin E. Aquac. Nutr. 1997, 3, 91–97. [Google Scholar] [CrossRef]
- Habryka, C.; Socha, R.; Juszczak, L. Effect of Bee Pollen Addition on the Polyphenol Content, Antioxidant Activity, and Quality Parameters of Honey. Antioxidants 2021, 10, 810. [Google Scholar] [CrossRef]
- Okhionkpamwonyi, O.; Edema, C. Effects of supplemental vitamin C (Ascorbic Acid) on the growth and health of African catfish Clarias gariepinus. J. Appl. Sci. Environ. Manag. 2017, 21, 177–183. [Google Scholar] [CrossRef] [Green Version]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant Potential of Propolis, Bee Pollen, and Royal Jelly: Possible Medical Application. Oxidative Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef]
- Galeotti, F.; Barile, E.; Curir, P.; Dolci, M.; Lanzotti, V. Flavonoids from carnation (Dianthus caryophyllus) and their antifungal activity. Phytochem. Lett. 2008, 1, 44–48. [Google Scholar] [CrossRef]
- Rzepecka-Stojko, A.; Stojko, J.; Kurek-Górecka, A.; Górecki, M.; Kabała-Dzik, A.; Kubina, R.; Moździerz, A.; Buszman, E. Polyphenols from Bee Pollen: Structure, Absorption, Metabolism and Biological Activity. Molecules 2015, 20, 21732–21749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouda, Z.; Zerdani, I.; Kalalou, I.; Faid, M.; Ahami, M.T. The Antibacterial Activity of Moroccan Bee Bread and Bee-Pollen (Fresh and Dried) against Pathogenic Bacteria. Res. J. Microbiol. 2011, 6, 376–384. [Google Scholar]
- Ramos, O.Y.; Basualdo, M.; Libonatti, C.; Vega, M.F. Current status and application of lactic acid bacteria in animal production systems with a focus on bacteria from honey bee colonies. J. Appl. Microbiol. 2020, 128, 1248–1260. [Google Scholar] [CrossRef] [Green Version]
- Reda, R.M.; El-Hady, M.A.; Selim, K.M.; El-Sayed, H.M. Comparative study of three predominant gut Bacillus strains and a commercial B. amyloliquefaciens as probiotics on the performance of Clarias gariepinus. Fish Shellfish Immunol. 2018, 80, 416–425. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.O.; Dhama, K.; Abdel-Latif, H.M.R. The functionality of probiotics in aquaculture: An overview. Fish Shellfish Immunol. 2021, 117, 36–52. [Google Scholar] [CrossRef] [PubMed]
- Kačániová, M.; Rovná, K.; Arpášová, H.; Hleba, L.; Petrová, J.; Haščík, P.; Čuboň, J.; Pavelková, A.; Chlebo, R.; Bobková, A.; et al. The effects of bee pollen extracts on the broiler chicken’s gastrointestinal microflora. Res. Vet. Sci. 2013, 95, 34–37. [Google Scholar] [CrossRef]
- Kupren, K.; Nowosad, J.; Żarski, D.; Targońska, K.; Hakuć-Błażowska, A.; Kucharczyk, D. Early development and allometric growth in laboratory-reared European chub Leuciscus cephalus (Linnaeus, 1758). Turk. J. Fish. Aquat. Sci. 2015, 15, 391–398. [Google Scholar] [CrossRef]
- Kujawa, R.; Lach, M.; Pol, P.; Ptaszkowski, M.; Mamcarz, A.; Nowosad, J.; Furgała-Selezniow, G.; Kucharczyk, D. Influence of water salinity on the survival of embryos and growth of the sichel larvae Pelecus cultratus (L.) under controlled conditions. Aquac. Res. 2017, 48, 1302–1314. [Google Scholar] [CrossRef]
- Sebesta, R.; Kucharczyk, D.; Nowosad, J.; Sikora, M.; Stejskal, V. Effect of temperature on growth and survival of maraena whitefish Coregonus maraena (Bloch 1779) larvae in controlled condition. Aquac. Res. 2018, 49, 2988–2994. [Google Scholar] [CrossRef]
- Nowosad, J.; Kupren, K.; Biegaj, M.; Kucharczyk, D. Allometric and ontogenetic larval development of common barbel during rearing under optimal conditions. Animal 2021, 15, 100107. [Google Scholar] [CrossRef] [PubMed]
- Stejskal, V.; Gebauer, T.; Sebesta, R.; Nowosad, J.; Sikora, M.; Biegaj, M.; Kucharczyk, D. Effect of feeding strategy on survival, growth, intestine development, and liver status of maraena whitefish Coregonus maraena larvae. J. World Aquac. Soc. 2021, 52, 829–842. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Dawood, M.A.O.; Alagawany, M.; Faggio, C.; Nowosad, J.; Kucharczyk, D. Health benefits and potential applications of fucoidan (FCD) extracted from brown seaweeds in aquaculture: An updated review. Fish Shellfish Immunol. 2022, 122, 115–130. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Measured Parameters | Experimental Groups | |||
|---|---|---|---|---|
| C | BP1 | BP2 | BP3 | |
| Initial weight (g) | 0.48 ± 0.17 | 0.48 ± 0.17 | 0.48 ± 0.17 | 0.48 ± 0.17 |
| Initial length (mm) | 38.9 ± 4.5 | 38.9 ± 4.5 | 38.9 ± 4.5 | 38.9 ± 4.5 |
| Final weight (g) | 5.01 ± 2.05 b | 7.38 ± 2.92 a | 6.63 ± 1.84 a | 7.45 ± 3.17 a |
| Final length (mm) | 84.7 ± 9.7 b | 95.9 ± 12.1 a | 93.8 ± 8.4 a | 95.2 ± 12.8 a |
| Growth (g/d) | 0.23 ± 0.02 b | 0.34 ± 0.05 a | 0.31 ± 0.03 a | 0.35 ± 0.05 a |
| Specific growth rate (%/d) | 11.70 ± 0.45 b | 13.63 ± 0.64 a | 13.11 ± 0.30 a | 13.68 ± 0.65 a |
| Condition factor | 0.82 ± 0.04 | 0.83 ± 0.02 | 0.81 ± 0.03 | 0.86 ± 0.06 |
| Relative growth rate (%) | 941.33 ± 91.69 b | 1436.76 ± 196.29 a | 1277.30 ± 82.60 a | 1451.96 ±193.72 a |
| Cumulative natural mortality (%) | 11.3 ± 3.1 | 6.0 ± 5.3 | 5.3 ± 2.3 | 6.0 ± 3.5 |
| Cannibalism factor (%) | 10.0 ± 2.0 | 6.0 ± 2.0 | 6.7 ± 1.2 | 6.7 ± 4.6 |
| Total survival rate (%) | 78.7 ± 4.6 | 88.0 ± 3.5 | 88.0 ± 2.0 | 87.3 ± 4.2 |
| TAC | TFC | TPC | ||
|---|---|---|---|---|
| mM TE/g | mg TE/g | IC50 mg/mL | mg QE/g | mg GAE/ g |
| 40.29 ± 1.24 | 10.27 ± 0.33 | 1.87 ± 0.07 | 19.64 ± 0.58 | 28.14 ± 0.23 |
| Analyzed Elements (mg/kg) | Experimental Groups | |||
|---|---|---|---|---|
| C | BP1 | BP2 | BP3 | |
| Ca | 77.13 ± 64.56 c | 146.80 ± 30.98 ab | 177.68 ± 43.6 a | 102.30 ± 19.99 bc |
| Zn | 12.3 ± 2.60 | 12.42 ± 2.28 | 19.40 ± 14.32 | 11.87 ± 1.37 |
| Mn | 0.15 ± 0.15 | 0.19 ± 0.16 | 0.19 ± 0.11 | 0.29 ± 0.16 |
| Fe | 2.86 ± 0.60 | 2.73 ± 0.76 | 3.06 ± 1.08 | 3.57 ± 1.93 |
| K | 2871.75 ± 145.46 | 3010.25 ± 52.90 | 2952.25 ± 102.59 | 2980.25 ± 130.23 |
| Na | 498.58 ± 55.32 | 464.50 ± 65.60 | 508.35 ± 45.59 | 507.20 ± 22.72 |
| Mg | 293.48 ± 22.16 | 305.55 ± 7.06 | 288.60 ± 18.11 | 317.00 ± 10.17 |
| Parameters (µm) | Experimental Groups | |||
|---|---|---|---|---|
| C | BP1 | BP2 | BP3 | |
| Villi height | 387.46 ± 83.17 b | 471.50 ± 63.90 a | 532.48 ± 67.34 a | 492.73 ± 110.22 a |
| Villi width | 71.88 ± 11.69 | 76.61 ± 9.18 | 73.29 ± 10.48 | 72.04 ± 12.29 |
| Enterocytes height | 28.94 ± 3.28 b | 34.85 ± 6.33 a | 31.83 ± 6.06 b | 30.20 ± 9.84 b |
| Width of intestinal wall | 67.51 ± 20.95 b | 76.85 ± 12.80 ab | 83.38 ± 24.67 a | 83.48 ± 19.78 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowosad, J.; Jasiński, S.; Arciuch-Rutkowska, M.; Abdel-Latif, H.M.R.; Wróbel, M.; Mikiewicz, M.; Zielonka, Ł.; Kotsyumbas, I.Y.; Muzyka, V.P.; Brezvyn, O.M.; et al. Effects of Bee Pollen on Growth Performance, Intestinal Microbiota and Histomorphometry in African Catfish. Animals 2023, 13, 132. https://doi.org/10.3390/ani13010132
Nowosad J, Jasiński S, Arciuch-Rutkowska M, Abdel-Latif HMR, Wróbel M, Mikiewicz M, Zielonka Ł, Kotsyumbas IY, Muzyka VP, Brezvyn OM, et al. Effects of Bee Pollen on Growth Performance, Intestinal Microbiota and Histomorphometry in African Catfish. Animals. 2023; 13(1):132. https://doi.org/10.3390/ani13010132
Chicago/Turabian StyleNowosad, Joanna, Szymon Jasiński, Martyna Arciuch-Rutkowska, Hany M. R. Abdel-Latif, Marcin Wróbel, Mateusz Mikiewicz, Łukasz Zielonka, Ihor Y. Kotsyumbas, Viktor P. Muzyka, Oksana M. Brezvyn, and et al. 2023. "Effects of Bee Pollen on Growth Performance, Intestinal Microbiota and Histomorphometry in African Catfish" Animals 13, no. 1: 132. https://doi.org/10.3390/ani13010132