
Nursing of Pike-Perch (Sander lucioperca) in Recirculating Aquaculture System (RAS) Provides Growth Advantage in Juvenile Growth Phase

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
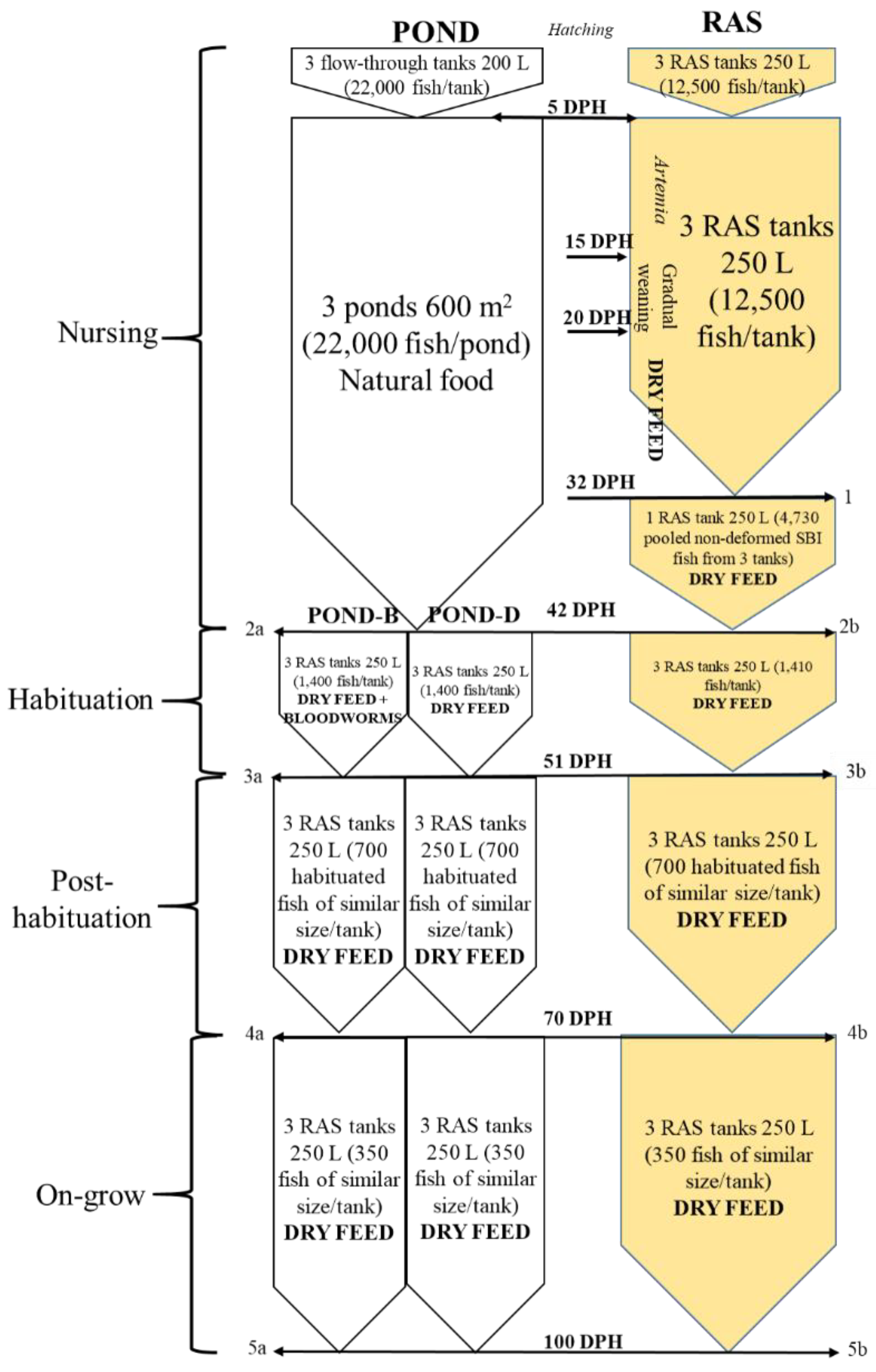
2. Materials and Methods
2.1. Fish Origin
2.2. Nursing Period
2.2.1. Pond
2.2.2. RAS
2.3. Habituation Period—Transport of Pond-Nursed Juveniles to RAS
2.4. Post-Habituation Period—After Transition of Pond-Nursed Juveniles to the RAS
2.5. Juvenile On-Grow Period
2.6. Data Analysis
3. Results
3.1. Larval and Early Post-Larval Rearing
3.2. Habituation Period
3.3. Post-Habituation Period
3.4. On-Growing Period
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bohnes, F.A.; Hauschild, M.Z.; Schlundt, J.; Nielsen, M.; Laurent, A. Environmental sustainability of future aquaculture production: Analysis of Singaporean and Norwegian policies. Aquaculture 2022, 549, 737717. [Google Scholar] [CrossRef]
- Froehlich, H.E.; Koehn, J.Z.; Holsman, K.K.; Halpern, B.S. Emerging trends in science and news of climate change threats to and adaptation of aquaculture. Aquaculture 2022, 549, 737812. [Google Scholar] [CrossRef]
- Martins, C.I.M.; Eding, E.H.; Verdegem, M.C.J.; Heinsbroek, L.T.N.; Schneider, O.; Blancheton, J.P.; Roque d’Orbcastel, E.; Verreth, J.A.J. New developments in recirculating aquaculture systems in Europe: A perspective on environmental sustainability. Aquac. Eng. 2010, 43, 83–93. [Google Scholar] [CrossRef] [Green Version]
- Colchen, T.; Gisbert, E.; Krauss, D.; Ledoré, Y.; Pasquet, A.; Fontaine, P. Improving pikeperch larviculture by combining environmental, feeding and populational factors. Aquac. Rep. 2020, 17, 100337. [Google Scholar] [CrossRef]
- Policar, T.; Křišťan, J.; Blecha, M.; Vaniš, J. Adaptation and Culture of Pikeperch (Sander lucioperca L.) Juveniles in Recirculating Aquaculture System (RAS). In Edition of Methodologies; Kouba, A., Ed.; Faculty of Fisheries and Protection of Waters, University of South Bohemia: České Budějovice, Chech Republic, 2014; ISBN 978-80-7514-044-9. [Google Scholar]
- Kestemont, P.; Xueliang, X.; Hamza, N.; Maboudou, J.; Toko, I.I. Effect of weaning age and diet on pikeperch larviculture. Aquaculture 2007, 264, 197–204. [Google Scholar] [CrossRef]
- Ljubobratović, U.; Fazekas, G.; Koljukaj, A.; Ristović, T.; Vass, V.; Ardó, L.; Stanisavljević, N.; Vukotić, G.; Pešić, M.; Milinčić, D.; et al. Pike-perch larvae growth in response to administration of lactobacilli-enriched inert feed during first feeding. Aquaculture 2021, 542, 736901. [Google Scholar] [CrossRef]
- Szkudlarek, M.; Zakęś, Z. Effect of stocking density on survival and growth performance of pikeperch, Sander lucioperca (L.), larvae under controlled conditions. Aquac. Int. 2007, 15, 67–81. [Google Scholar] [CrossRef]
- Lepič, P.; Buřič, M.; Hajíček, J.; Kozák, P. Adaptation to pelleted feed in pikeperch fingerlings: Learning from the trainer fish over gradual adaptation from natural food. Aquat. Living Resour. 2017, 30, 8. [Google Scholar] [CrossRef] [Green Version]
- Ljubobratović, U.; Péter, G.; Horvath, Z.; Ristović, T.; Rónyai, A. Effect of parental origin on dry feed habituation and intensive on-growing results in pikeperch (Sander lucioperca) Offspring. Turk. J. Fish. Aquat. Sci. 2018, 18, 425–433. [Google Scholar] [CrossRef]
- Policar, T.; Stejskal, V.; Kristan, J.; Podhorec, P.; Svinger, V.; Blaha, M. The effect of fish size and stocking density on the weaning success of pond-cultured pikeperch Sander lucioperca L. juveniles. Aquac. Int. 2013, 21, 869–882. [Google Scholar] [CrossRef]
- Overton, J.L.; Toner, D.; Policar, T.; Kucharczyk, D. Commercial production: Factors for success and limitations in European percid fish culture. In Biology and Culture of Percid Fishes; Springer: Dordrecht, The Netherlands, 2015; pp. 881–890. [Google Scholar]
- Policar, T.; Schaefer, F.J.; Panana, E.; Meyer, S.; Teerlinck, S.; Toner, D.; Żarski, D. Recent progress in European percid fish culture production technology-tackling bottlenecks. Aquac. Int. 2019, 27, 1151–1174. [Google Scholar] [CrossRef]
- Schaefer, F.J.; Overton, J.L.; Krüger, A.; Kloas, W.; Wuertz, S. Influence of batch-specific biochemical egg characteristics on embryogenesis and hatching success in farmed pikeperch. Animal 2018, 12, 2327–2334. [Google Scholar] [CrossRef]
- El Kertaoui, N.; Lund, I.; Assogba, H.; Domínguez, D.; Izquierdo, M.S.; Baekelandt, S.; Cornet, V.; Mandiki, S.N.M.; Montero, D.; Kestemont, P. Key nutritional factors and interactions during larval development of pikeperch (Sander lucioperca). Sci. Rep. 2019, 9, 7074. [Google Scholar] [CrossRef] [Green Version]
- Imentai, A.; Rašković, B.; Steinbach, C.; Rahimnejad, S.; Yanes-Roca, C.; Policar, T. Effects of first feeding regime on growth performance, survival rate and development of digestive system in pikeperch (Sander lucioperca) larvae. Aquaculture 2020, 529, 735636. [Google Scholar] [CrossRef]
- Ljubobratović, U.; Péter, G.; Sándor, Y.; Kugyela, N.; Rónyai, A. The effect of hormonal preparation (gonadotropins vs. gonadoliberins) on pre-seasonally obtained eggs and larvae quality in pikeperch (Sander lucioperca L.). Aquac. Int. 2019, 27, 1009–1024. [Google Scholar] [CrossRef] [Green Version]
- Schäfer, N.; Kaya, Y.; Rebl, H.; Stüeken, M.; Rebl, A.; Nguinkal, J.A.; Franz, G.P.; Brunner, R.M.; Goldammer, T.; Grunow, B.; et al. Insights into early ontogenesis: Characterization of stress and development key genes of pikeperch (Sander lucioperca) in vivo and in vitro. Fish Physiol. Biochem. 2021, 47, 515–532. [Google Scholar] [CrossRef]
- Ljubobratović, U.; Péter, G.; Alvestad, R.; Horváth, Z.; Rónyai, A. Alcalase enzyme treatment affects egg incubation and larval quality in pikeperch (Sander lucioperca). Aquac. Int. 2019, 27, 917–929. [Google Scholar] [CrossRef]
- Ljubobratović, U.; Kucska, B.; Sandor, Z.; Peteri, A.; Rónyai, A. Effects of stocking density, feeding technique and vitamin C supplementation on the habituation on dry feed of pikeperch (Sander lucioperca) pond reared juveniles. Iran. J. Fish. Sci. 2016, 15, 1337–1347. [Google Scholar]
- Horváth, Z.; Németh, S.; Beliczky, G.; Felföldi, Z.; Bercsényi, M. Comparison of efficiencies of using trainer fish and shape or taste modified feed for enchancing direct weaning of pikeperch (Sander lucioperca L.) yearlings on dry feed. Ribar. Croat. J. Fish. 2013, 71, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Arifin, O.Z.; Prakoso, V.A.; Subagja, J.; Kristanto, A.H.; Pouil, S.; Slembrouck, J. Effects of stocking density on survival, food intake and growth of giant gourami (Osphronemus goramy) larvae reared in a recirculating aquaculture system. Aquaculture 2019, 509, 159–166. [Google Scholar] [CrossRef]
- Gubiani, E.A.; Ruaro, R.; Ribeiro, V.R.; de Santa Fé, U.M.G. Relative condition factor: Le Cren’s legacy for fisheries science. Acta Limnol. Bras. 2020, 32, e3. [Google Scholar] [CrossRef]
- IBM SPSS Statistics for Windows, Version 21.0.; IBM Corp: Armonk, NY, USA, 2012.
- Abdel-Aziz, M.F.A.; Abou-Zied, R.M.; Allam, S.M.; Mohammed, R.A. Effect of stocking density and water exchange period on growth performance, feed utilization and body chemical composition on Rabbitfish Siganus rivulatus juvenile under laboratory condition. Egypt. J. Aquat. Biol. Fish. 2016, 20, 15–33. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.F.M. Effects of stocking density and feeding levels on growth and feed efficiency of Nile tilapia (Oreochromis niloticus L.) fry. Aquac. Res. 2002, 33, 621–626. [Google Scholar] [CrossRef]
- Ljubobratović, U.; Kosanovic, D.; Demény, F.Z.; Krajcsovics, A.; Vukotic, G.; Stanisavljevic, N.; Golic, N.; Jeney, G.; Lukic, J. The effect of live and inert feed treatment with lactobacilli on weaning success in intensively reared pike-perch larvae. Aquaculture 2020, 516, 734608. [Google Scholar] [CrossRef]
- Yanes-Roca, C.; Holzer, A.; Mraz, J.; Veselý, L.; Malinovskyi, O.; Policar, T. Improvements on live feed enrichments for pikeperch (Sander lucioperca) larval culture. Animals 2020, 10, 401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, G.P.; Lewerentz, L.; Grunow, B. Observations of growth changes during the embryonic-larval-transition of pikeperch (Sander lucioperca) under near-natural conditions. J. Fish Biol. 2021, 99, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Kupren, K.; Palińska-Żarska, K.; Krejszeff, S.; Żarski, D. Early development and allometric growth in hatchery-reared Eurasian perch, Perca fluviatilis L. Aquac. Res. 2019, 50, 2528–2536. [Google Scholar] [CrossRef]
- Famoofo, O.O.; Abdul, W.O. Biometry, condition factors and length-weight relationships of sixteen fish species in Iwopin fresh-water ecotype of Lekki Lagoon, Ogun State, Southwest Nigeria. Heliyon 2020, 6, E02957. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.-O.; Oh, S.-Y.; Kim, T. Effect of fasting and refeeding on juvenile leopard mandarin fish Siniperca scherzeri. Animals 2022, 12, 889. [Google Scholar] [CrossRef]
- Engrola, S.; Figueira, L.; Conceição, L.E.C.; Gavaia, P.J.; Ribeiro, L.; Dinis, M.T. Co-feeding in Senegalese sole larvae with inert diet from mouth opening promotes growth at weaning. Aquaculture 2009, 288, 264–272. [Google Scholar] [CrossRef]
- Imentai, A.; Gilannejad, N.; Martínez-Rodríguez, G.; López, F.J.M.; Martínez, F.P.; Pěnka, T.; Dzyuba, V.; Dadras, H.; Policar, T. Effects of first feeding regime on gene expression and enzyme activity in pikeperch (Sander lucioperca) larvae. Front. Mar. Sci. 2022, 9, 864536. [Google Scholar] [CrossRef]
- Vestrum, R.I.; Attramadal, K.J.K.; Winge, P.; Li, K.; Olsen, Y.; Bones, A.M.; Vadstein, O.; Bakke, I. Rearing water treatment induces microbial selection influencing the microbiota and pathogen associated transcripts of cod (Gadus morhua) larvae. Front. Microbiol. 2018, 9, 851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chislock, M.F.; Doster, E.; Zitomer, R.A.; Wilson, A.E. Eutrophication: Causes, consequences, and controls in aquatic ecosystems. Nat. Educ. Knowl. 2013, 4, 10. [Google Scholar]
- Attramadal, K.J.K.; Øie, G.; Størseth, T.R.; Alver, M.O.; Vadstein, O.; Olsen, Y. The effects of moderate ozonation or high intensity UV-irradiation on the microbial environment in RAS for marine larvae. Aquaculture 2012, 330–333, 121–129. [Google Scholar] [CrossRef]
- Deng, Y.; Verdegem, M.C.J.; Eding, E.; Kokou, F. Effect of rearing systems and dietary probiotic supplementation on the growth and gut microbiota of Nile tilapia (Oreochromis niloticus) larvae. Aquaculture 2022, 546, 737297. [Google Scholar] [CrossRef]
- Xie, M.; Hao, Q.; Olsen, R.E.; Ringø, E.; Yang, Y.; Zhang, Z.; Ran, C.; Zhou, Z. Growth performance, hepatic enzymes, and gut health status of common carp (Cyprinus carpio) in response to dietary Cetobacterium somerae fermentation product. Aquac. Rep. 2022, 23, 101046. [Google Scholar] [CrossRef]
- Wang, A.; Zhang, Z.; Ding, Q.; Yang, Y.; Bindelle, J.; Ran, C.; Zhou, Z. Intestinal Cetobacterium and acetate modify glucose homeostasis via parasympathetic activation in zebrafish. Gut Microbes 2021, 13, 1–15. [Google Scholar] [CrossRef]
- Blecha, M.; Malinovskyi, O.; Veselý, L.; Křišťan, J.; Policar, T. Swim bladder inflation failure in pikeperch (Sander lucioperca) larvae in pond culture. Aquac. Int. 2019, 27, 983–989. [Google Scholar] [CrossRef]
- Fazekas, G.; Vass, V.; Demény, F.; Tóth, F.; Ljubobratović, U. The effect of different surface-cleaning devices on the success of swim bladder inflation in zander larvae. N. Am. J. Aquac. 2021, 83, 78–82. [Google Scholar] [CrossRef]
- Woolley, L.D.; Qin, J.G. Swimbladder inflation and its implication to the culture of marine finfish larvae. Rev. Aquac. 2010, 2, 181–190. [Google Scholar] [CrossRef]
- Peruzzi, S.; Westgaard, J.I.; Chatain, B. Genetic investigation of swimbladder inflation anomalies in the European sea bass, Dicentrarchus labrax L. Aquaculture 2007, 265, 102–108. [Google Scholar] [CrossRef] [Green Version]
- Policar, T.; Blecha, M.; Křišťan, J.; Mráz, J.; Velíšek, J.; Stará, A.; Stejskal, V.; Malinovskyi, O.; Svačina, P.; Samarin, A.M. Comparison of production efficiency and quality of differently cultured pikeperch (Sander lucioperca L.) juveniles as a valuable product for ongrowing culture. Aquac. Int. 2016, 24, 1607–1626. [Google Scholar] [CrossRef]
- Schwebel, L.N.; Stuart, K.; Lowery, M.S.; Wegner, N.C. Swim bladder inflation failure affects energy allocation, growth, and feed conversion of California Yellowtail (Seriola dorsalis) in aquaculture. Aquaculture 2018, 497, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Barcellos, L.J.G.; Volpato, G.L.; Barreto, R.E.; Coldebella, I.; Ferreira, D. Chemical communication of handling stress in fish. Physiol. Behav. 2011, 103, 372–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamstra, A. Production Costs of Pike-Perch Fingerlings (Sander lucioperca): A Bio-Economic Model. Report Produced in the Framework of the CRAFT Project “LUCIOPERCA: Bio-Economic Feasibility of Intensive Culture of Pike-Perch” 2003. No. C056/03. RIVO. Available online: https://edepot.wur.nl/151171 (accessed on 19 December 2022).
- Schram, E.; Kals, J.; Toner, D.R.; Policar, T. Production Costs of Perch (Perca fluvialitis) Juveniles. Securing Juvenile Production of Eurasian Perch by Improving Reproduction and Larval Rearing. Report/Wageningen IMARES 2006, No. C071/06. IMARES. Available online: https://edepot.wur.nl/148159 (accessed on 19 December 2022).
- Zakes, Z. Pike perch, Sander lucioperca, production in recirculating systems. Bull. VURH Vodnany 2004, 39, 136–140. [Google Scholar]
- Ljubobratović, U.; Kosanovic, D.; Vukotic, G.; Molnar, Z.; Stanisavljevic, N.; Ristovic, T.; Peter, G.; Lukic, J.; Jeney, G. Supplementation of lactobacilli improves growth, regulates microbiota composition and suppresses skeletal anomalies in juvenile pike-perch (Sander lucioperca) reared in recirculating aquaculture system (RAS): A pilot study. Res. Vet. Sci. 2017, 115, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Yanes-Roca, C.; Mráz, J.; Born-Torrijos, A.; Holzer, A.S.; Imentai, A.; Policar, T. Introduction of rotifers (Brachionus plicatilis) during pikeperch first feeding. Aquaculture 2018, 497, 260–268. [Google Scholar] [CrossRef]
- Mes, D.; Palstra, A.P.; Henkel, C.V.; Mayer, I.; Vindas, M.A. Swimming exercise enhances brain plasticity in fish. R. Soc. Open Sci. 2020, 7, 191640. [Google Scholar] [CrossRef] [Green Version]
- Pittman, K.; Yúfera, M.; Pavlidis, M.; Geffen, A.J.; Koven, W.; Ribeiro, L.; Zambonino-Infante, J.L.; Tandler, A. Fantastically plastic: Fish larvae equipped for a new world. Rev. Aquac. 2013, 5, S224–S267. [Google Scholar] [CrossRef] [Green Version]
- Kozłowski, M.; Zakęś, Z.; Szczepkowski, M.; Wunderlich, K.; Piotrowska, I.; Szczepkowska, B. Impact of light intensity on the results of rearing juvenile pikeperch, Sander lucioperca (L.), in recirculating aquaculture systems. Fish. Aquat. Life 2010, 18, 77–84. [Google Scholar] [CrossRef]
- Luchiari, A.C.; De Morais Freire, F.A.; Koskela, J.; Pirhonen, J. Light intensity preference of juvenile pikeperch Sander lucioperca (L.). Aquac. Res. 2006, 37, 1572–1577. [Google Scholar] [CrossRef]


{kind=link}
| Parameters | Pond | RAS |
|---|---|---|
| Final weight (mg) | 469.6 ± 69.4 A | 929.18 ± 250.4 B |
| Final length (mm) | 39.4 ± 2.2 A | 46.1 ± 3.8 B |
| Specific growth rate (%/day) | 16.3 ± 0.4 A | 17.8 ± 0.7 B |
| Allometric coefficient (b) | 2.11 A | 3.01 B |
| Relative condition factor (Kn) | 1.00 ± 0.09 A | 1.01 ± 0.17 A |
| Survival (%) | 67.3 A | 11.3 *B |
| Parameters | Pond-B | Pond-D | RAS |
|---|---|---|---|
| Initial weight (g) | 0.44 ± 0.02 A | 0.43 ± 0.01 A | 0.93 ± 0.08 B |
| Final weight (g) | 1.0 ± 0.0 A | 0.7 ± 0.0 B | 1.8 ± 0.8 C |
| Final length (mm) | 44.5 ± 0.4 A | 40.5 ± 0.6 B | 53.5 ± 1.2 C |
| Specific growth rate (SGR) (%/day) | 7.7 ± 0.1 A | 4.9 ± 0.5 B | 6.1 ± 0.6 A |
| Allometric coefficient (b) | 3.04 ± 0.45 A | 3.02 ± 0.36 A | 2.95 ± 0.46 A |
| Relative condition factor (Kn) | 1.01 ± 0.00 A | 1.01 ± 0.00 A | 1.01 ± 0.00 A |
| Cannibalism (%) | 24.2 ± 2.4 A | 19.1 ± 3.1 A | 5.5 ± 0.6 B |
| Mortality (%) | 5.4 ± 1.6 | 5.4 ± 1.2 | 4.7 ± 0.5 |
| Survival (%) | 70.4 ± 2.9 A | 75.5 ± 2.2 A | 89.8 ± 0.4 B |
| Cumulative survival (%) | 47.3 ± 1.9 A | 50.8 ± 1.5 A | 10.1 ± 0.1 B |
| Parameters | Pond-B | Pond-D | RAS |
|---|---|---|---|
| Initial weight (g) | 1.90 ± 0.03 A | 0.74 ± 0.05 B | 1.80 ± 0.04 C |
| Final weight (g) | 4.7 ± 0.6 A | 4.0 ± 0.1 A | 6.0 ± 0.0 B |
| Final length (mm) | 78.5 ± 2.0 A | 72.4 ± 1.1 B | 81.3 ± 1.0 A |
| Specific growth rate (SGR) (%/day) | 8.5 ± 0.6 A | 9.3 ± 0.5 A | 6.7 ± 0.1 B |
| Feed conversion ratio (FCR) | 0.72 ± 0.11 A | 0.70 ± 0.06 A | 0.79 ± 0.02 A |
| Allometric coefficient (b) | 2.77 ± 0.82 A | 2.93 ± 0.23 A | 3.09 ± 0.27 A |
| Relative condition factor (Kn) | 1.01 ± 0.00 A | 1.00 ± 0.00 A | 1.01 ± 0.00 A |
| Cannibalism (%) | 19.6 ± 2.3 A | 27.2 ± 2.6 B | 0.3 ± 0.3 C |
| Mortality (%) | 3.7 ± 1.1 A | 5.7 ± 1.2 A | 1 ± 0.2 B |
| Survival (%) | 76.7 ± 1.6 A | 67.0 ± 3.8 B | 98.7 ± 0.2 C |
| Cumulative survival (%) | 36.3 ± 1.5 A | 34.0 ± 1.6 A | 10 ± 0.1 B |
| Parameters | Pond-B | Pond-D | RAS |
|---|---|---|---|
| Initial weight (g) | 4.8 ± 0.1 AB | 4.3 ± 0.3 B | 5.4 ± 0.3 A |
| Final weight (g) | 16.3 ± 0.7 A | 14.5 ± 0.3 B | 18.7 ± 0.4 C |
| Final length (mm) | 121.6 ± 1.6 A | 116.5 ± 1.3 B | 121.9 ± 1.5 A |
| Specific growth rate (SGR) (%/day) | * 4.0 ± 0.1 A | * 3.6 ± 0.2 A | * 4.6 ± 0.2 B |
| Feed conversion ratio (FCR) | * 0.72 ± 0.04 A | * 0.86 ± 0.07 A | * 0.54 ± 0.07 B |
| Allometric coefficient (b) | 3.17 ± 0.20 A | 3.13 ± 0.04 A | 2.87 ± 0.14 A |
| Relative condition factor (Kn) | 1.00 ± 0.00 A | 1.00 ± 0.00 A | 1.01 ± 0.00 A |
| Cannibalism (%) | 1.4 ± 0.3 A | 0.8 ± 0.6 A | 1.4 ± 1.8 A |
| Mortality (%) | 0.5 ± 0.3 A | 0.3 ± 0.3 A | 0.7 ± 0.7 A |
| Survival (%) | 98.1 ± 0.6 A | 99.0 ± 0.8 A | 97.9 ± 1.7 A |
| Cumulative survival (%) | 35.6 ± 1.6 A | 33.7 ± 1.3 A | 9.8 ± 0.2 B |
| Cumulative biomass (kg/12,500 larvae) | 72.7 ± 5.0 A | 61.2 ± 2.8 B | 23.0 ± 0.7 C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Péter, G.; Lukić, J.; Alvestad, R.; Horváth, Z.; Nagy, Z.; Rónyai, A.; Bársony, P.; Ljubobratović, U. Nursing of Pike-Perch (Sander lucioperca) in Recirculating Aquaculture System (RAS) Provides Growth Advantage in Juvenile Growth Phase. Animals 2023, 13, 347. https://doi.org/10.3390/ani13030347
Péter G, Lukić J, Alvestad R, Horváth Z, Nagy Z, Rónyai A, Bársony P, Ljubobratović U. Nursing of Pike-Perch (Sander lucioperca) in Recirculating Aquaculture System (RAS) Provides Growth Advantage in Juvenile Growth Phase. Animals. 2023; 13(3):347. https://doi.org/10.3390/ani13030347
Chicago/Turabian StylePéter, Géza, Jovanka Lukić, René Alvestad, Zoltán Horváth, Zoltán Nagy, András Rónyai, Péter Bársony, and Uroš Ljubobratović. 2023. "Nursing of Pike-Perch (Sander lucioperca) in Recirculating Aquaculture System (RAS) Provides Growth Advantage in Juvenile Growth Phase" Animals 13, no. 3: 347. https://doi.org/10.3390/ani13030347