
Effect of Application of Different Activation Media on Fertilization and Embryo Survival of Northern Pike, (Esox lucius) under Hatchery Conditions


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Content
2.2. Experimental Design: General Content
2.2.1. Design of Experiment No. 1
2.2.2. Design of Experiment No. 2
2.2.3. Mass Fertilization in Field Conditions
2.3. Incubation Conditions
2.4. Statistical Analysis
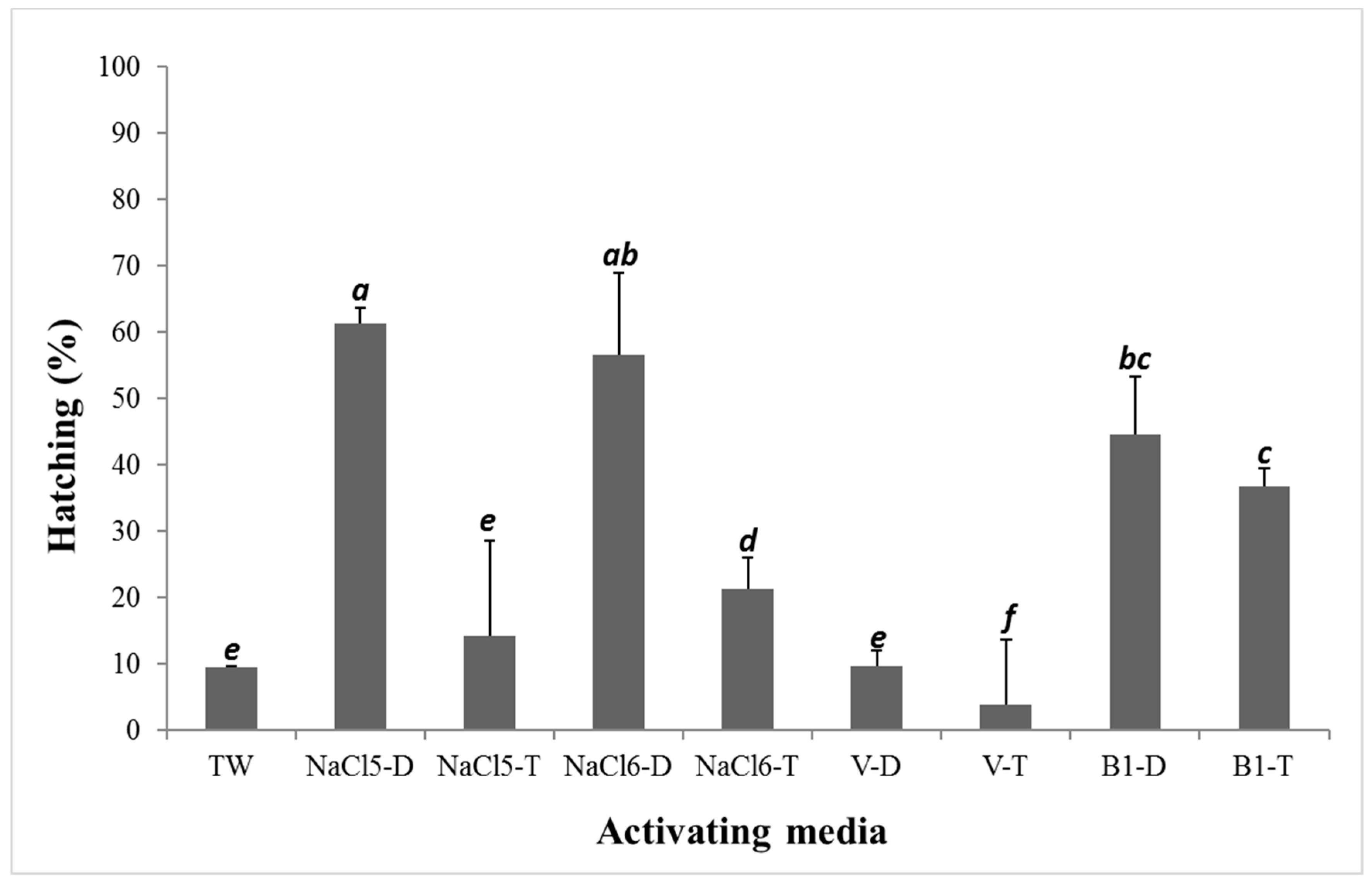
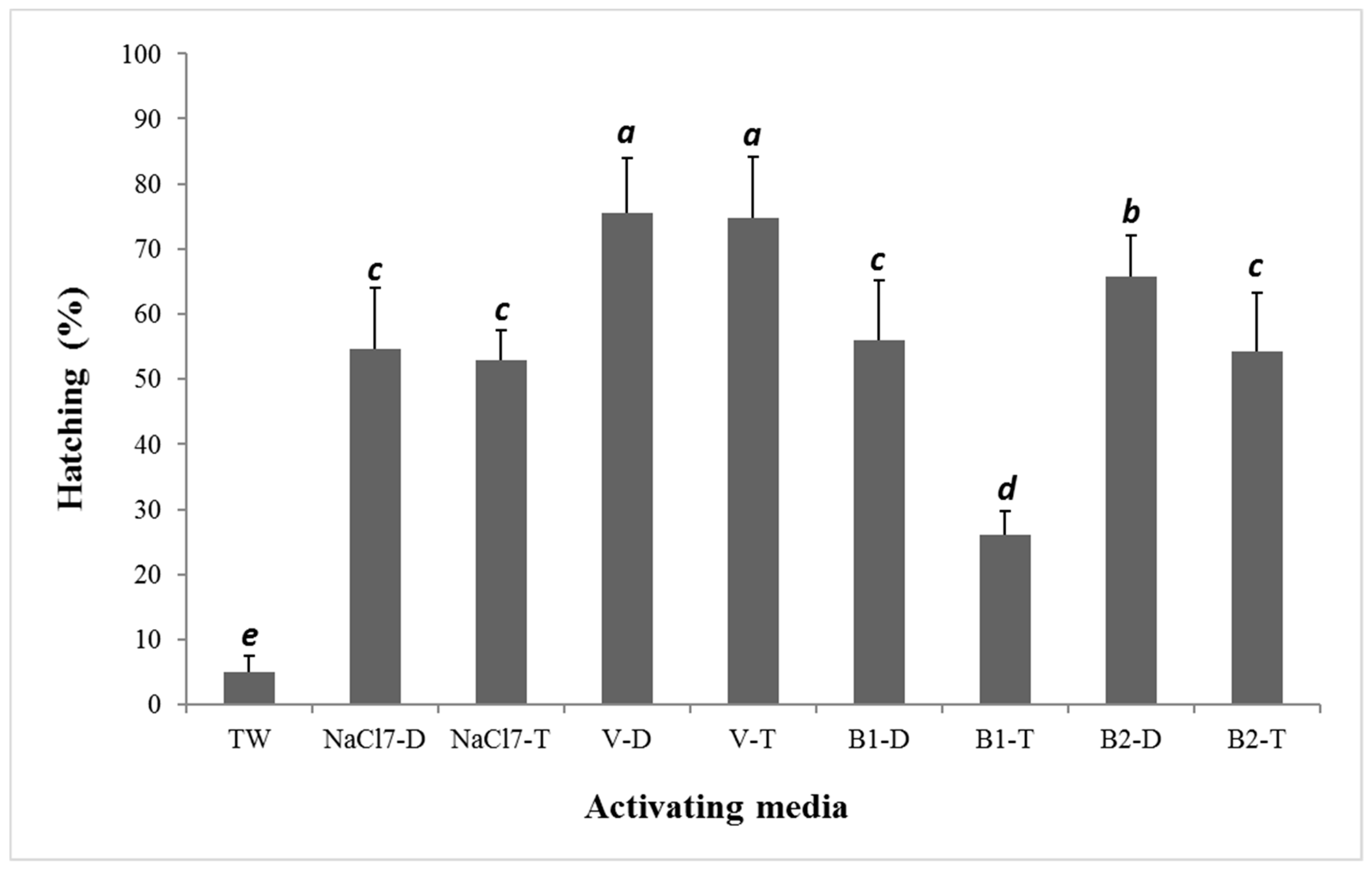
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2018–Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Kujawa, R.; Mamcarz, A.; Wyszomirska, E. Induced spawning in rudd (Scardinius erythrophthalmus L.). Pol. Arch. Hydrobiol. 1997, 44, 209–213. [Google Scholar]
- Kucharczyk, D.; Kujawa, R.; Mamcarz, A.; Wyszomirska, E. Artificial spawning in bream (Abramis brama L.). Pol. Arch. Hydrobiol. 1997, 44, 201–205. [Google Scholar]
- Szczerbowski, A.; Kucharczyk, D.; Mamcarz, A.; Łuczyński, M.; Targońska, K.; Kujawa, R. Artificial off-season spawning of Eurasian perch Perca fluviatilis L. Arch. Pol. Fish. 2009, 17, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Nowosad, J.; Targońska, K.; Chwaluczyk, R.; Kaszubowski, R.; Kucharczyk, D. Effect of temperature on the effectiveness of artificial reproduction of dace [Cyprinidae (Leuciscus leuciscus (L.))] under laboratory and field conditions. J. Therm. Biol. 2014, 45, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, J.; Kucharczyk, D.; Targońska, K.; Wyszomirska, E.; Chwaluczyk, R.; Kupren, K. The synergistic effect of temperature and hormonal stimulation on spawning efficiency of common barbel, Barbus barbus L. Turk. J. Fish. Aquat. Sci. 2016, 16, 517–524. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Nowosad, J.; Łuczyński, M.J.; Targońska, K. New technique for fertilizing eggs of burbot, asp and ide under hatchery conditions. Anim. Reprod. Sci. 2016, 172, 143–147. [Google Scholar] [CrossRef]
- Kristan, J.; Żarski, D.; Blecha, M.; Policar, T.; Malinovskyi, O.; Samarin, A.M.; Palińska-Żarska, K.; Nowosad, J.; Krejszeff, S.; Kucharczyk, D. Fertilizing ability of gametes at different post-activation times and the sperm-oocyte ratio in the artificial reproduction of pikeperch Sander lucioperca. Aquac. Res. 2018, 49, 1383–1388. [Google Scholar] [CrossRef]
- Müller, T.; Horváth, L.; Szabó, T.; Ittzés, I.; Bognár, A.; Faidt, P.; Ittzés, Á.; Urbányi, B.; Kucska, B. Novel method for induced propagation of fish: Sperm injection in oviducts and ovary/ovarian lavage with sperm. Aquaculture 2018, 482, 124–129. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Kucharczyk, D.J.; Nowosad, J.; Omirzhanova, N. Optimization of artificial insemination outcomes of African catfish (Clarias gariepinus) with differing hatchery conditions. Anim. Reprod. Sci. 2019, 211, 106222. [Google Scholar] [CrossRef]
- Fopp-Bayat, D.; Szadkowska, J.; Szczepkowski, M.; Szczepkowska, B.; Naumowicz, K. Gonadal analysis in the F1 progeny of a gynogenetic Siberian sturgeon Acipenser baerii female. Anim. Reprod. Sci. 2020, 221, 106548. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Żarski, D.; Targońska, K.; Łuczyński, M.J.; Szczerbowski, A.; Nowosad, J.; Kujawa, R.; Mamcarz, A. Induced Artificial Androgenesis in Common Tench, Tinca tinca (L.), Using Common Carp and Common Bream Eggs. Ital. J. Anim. Sci. 2014, 13, 2890. [Google Scholar] [CrossRef]
- Nowosad, J.; Kucharczyk, D.; Liszewski, T.; Targońska, K.; Kujawa, R. Comparison of temperature shock timing to induced artificial mitotic gynogenesis and androgenesis in common tench. Aquac. Int. 2015, 23, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.; Xin, M.; Gela, D.; Rodina, M.; Tučková, V.; Kašpar, V.; Siddique, M.A.M.; Shelton, W.L.; Linhart, O. Optimization of sterlet (Acipenser ruthenus) egg incubation. Anim. Reprod. Sci. 2020, 215, 106334. [Google Scholar] [CrossRef]
- Kujawa, R.; Mamcarz, A.; Kucharczyk, D. Effect of temperature on embryonic development of asp (Aspius aspius L.). Pol. Arch. Hydrobiol. 1997, 44, 139–143. [Google Scholar]
- Kupren, K.; Mamcarz, A.; Kucharczyk, D. Effect of variable and constant thermal conditions on embryonic and early larval development of fish from the genus Leuciscus (Cyprinidae, Teleostei). Czech J. Anim. Sci. 2011, 56, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Nowosad, J.; Sikora, M.; Kucharczyk, D. Survival rates and the occurrence of larval malformations, including Siamese twins, following fertilization of post-ovulatory aged oocytes in ide Leuciscus idus. Dis. Aquat. Org. 2018, 127, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Szabó, T. Ovulation induction in northern pike Esox lucius L. using different GnRH analogues, Ovaprim, Dagin and carp pituitary. Aquac. Res. 2003, 34, 479–486. [Google Scholar] [CrossRef]
- Szabó, T. Use of Carbopol resin for carp pituitary administration improves the fertilization percentage of northern pike (Esox lucius Linnaeus) eggs in commercial hatcheries. Hydrobiologia 2008, 601, 91–97. [Google Scholar] [CrossRef]
- Cejko, B.I.; Krejszeff, S.; Żarski, D.; Judycka, S.; Targońska, K.; Kucharczyk, D. Effect of carp pituitary homogenate (CPH) and sGnRHa (Ovaprim) on northern pike (Esox lucius) spermiation stimulation and its effect on quantity and quality of sperm. Anim. Reprod. Sci. 2018, 193, 217–225. [Google Scholar] [CrossRef]
- Cejko, B.I.; Sarosiek, B.; Dryl, K.; Judycka, S.; Szczepkowska, B.; Szczepkowski, M.; Kowalski, R.K. The effect of cryopreservation extender on sperm motility and hatch success in northern pike (Esox lucius). Aquaculture 2020, 514, 734482. [Google Scholar] [CrossRef]
- Cejko, B.I.; Krejszeff, S.; Judycka, S.; Targońska, K.; Kucharczyk, D. Effect of different treatment agents and post-treatment latency times on spermiation stimulation of northern pike (Esox lucius) under controlled conditions. Theriogenology 2020, 142, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Hakuć-Błażowska, A.; Kupren, K.; Turkowski, K.; Targońska, K.; Jamróz, M.; Krejszeff, S.; Kwiatkowski, M.; Żarski, D.; Kucharczyk, D. Comparison of Economic Effectiveness of Applying Different Hormonal Preparations for Reophile Cyprinid Fish Reproduction Stimulation Based on the Example of ASP Aspius aspius (L.) and Ide Leuciscus idus (L.). Pol. J. Nat. Sci. 2009, 24, 224–234. [Google Scholar] [CrossRef] [Green Version]
- Kowalski, R.K.; Cejko, B.I. Sperm quality in fish: Determinants and affecting factors. Theriogenology 2019, 135, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Billard, R. Reproduction in rainbow trout: Sex differentiation, dynamics of gametogenesis, biology and preservation of gametes. Aquaculture 1992, 100, 263–298. [Google Scholar] [CrossRef]
- Bobe, J.; Labbé, C. Egg and sperm quality in fish. Gen. Comp. Endocrinol. 2010, 165, 535–548. [Google Scholar] [CrossRef]
- Nowosad, J.; Dryl, K.; Kupren, K.; Kucharczyk, D. Inhibiting the influence of ovarian fluid on spermatozoa activation and spermatozoa kinetic characteristics in the common barbel Barbus barbus. Theriogenology 2020, 158, 250–257. [Google Scholar] [CrossRef]
- Nowosad, J.; Kucharczyk, D.; Sikora, M.; Kupren, K. Optimization of barbel (Barbus barbus L.) fertilization and effects of ovarian fluid when there are controlled conditions for gamete activations. Anim. Reprod. Sci. 2021, 224, 106652. [Google Scholar] [CrossRef]
- Babiak, I.; Glogowski, J.; Łuczyński, M.J.; Kucharczyk, D. Cryopreservation of the milt of the northern pike. J. Fish Biol. 1995, 46, 819–828. [Google Scholar] [CrossRef]
- De Montalembert, G.; Marcel, J.; Billard, R. La spermiation chez le brochet. 1—Evolution de la quantité de sperme récolté au cours de la saison de reproduction. Bull. Fr. Pêche Piscic. 1980, 276, 90–103. [Google Scholar] [CrossRef] [Green Version]
- Alavi, H.M.S.; Cosson, J. Sperm motility in fishes. (II) Effects of ions and osmolality: A review. Cell Biol. Int. 2006, 30, 1–14. [Google Scholar] [CrossRef]
- Alavi, S.H.; Rodina, M.; Viveiros, A.T.; Cosson, J.; Gela, D.; Boryshpolets, S.; Linhart, O. Effects of osmolality on sperm morphology, motility and flagellar wave parameters in Northern pike (Esox lucius L.). Theriogenology 2009, 72, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Cejko, B.I.; Targońska, K.; Kowalski, R.K.; Żarski, D.; Sarosiek, B.; Kucharczyk, D.; Glogowski, J. The effectiveness of hormonal preparations (Ovopel, Ovaprim, LHRHa, hCG and CPE) in stimulating spermiation in dace Leuciscus leuciscus (L.). J. Appl. Ichthyol. 2012, 28, 873–877. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Nowosad, J.; Kucharczyk, D.J.; Kupren, K.; Targońska, K.; Wyszomirska, E.; Kujawa, R. Out-of-season artificial reproduction of common dace (Leuciscus leuciscus L.) under controlled conditions. Anim. Reprod. Sci. 2019, 202, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, D.; Nowosad, J.; Wyszomirska, E.; Cejko, B.I.; Arciuch-Rutkowska, M.; Juchno, D.; Boroń, A. Comparison of artificial spawning effectiveness of hCG, CPH and GnRHa in combination with dopamine inhibitors in a wild strain of ide Leuciscus idus (L.) in hatchery conditions. Anim. Reprod. Sci. 2020, 221, 106543. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Szabó, T.; Kollár, T.; Csorbai, B.; Marinović, Z.; Horváth, L.; Kucska, B.; Bodnár, Á.; Urbányi, B.; Horváth, Á. Artificial insemination of African catfish (Clarias gariepinus) using cryopreserved sperm. Theriogenology 2019, 123, 145–150. [Google Scholar] [CrossRef]
- Dietrich, G.J.; Wojtczak, M.; Słowińska, M.; Dobosz, S.; Kuźmiński, H.; Ciereszko, A. Broken eggs decrease pH of rainbow trout (Oncorhynchus mykiss) ovarian fluid. Aquaculture 2007, 273, 748–751. [Google Scholar] [CrossRef]
- Wojtczak, M.; Dietrich, G.J.; Słowińska, M.; Dobosz, S.; Kuźmiński, H.; Ciereszko, A. Ovarian fluid pH enhances motility parameters of rainbow trout (Oncorhynchus mykiss) spermatozoa. Aquaculture 2007, 270, 259–264. [Google Scholar] [CrossRef]
- Bondarenko, V.; Drozd, B.; Policar, T. Effect of water temperature on egg incubation time and quality of newly hatched larvae of northern pike (Esox lucius L., 1758). J. Appl. Ichthyol. 2015, 31 (Suppl. S2), 45–50. [Google Scholar] [CrossRef]
- Babiak, I.; Glogowski, J.; Luczynski, M.J.; Luczynski, M. Effect of individual male variability on cryopreservation of northern pike, Esox lucius L., sperm. Aquac. Res. 1997, 28, 191–197. [Google Scholar] [CrossRef]
- Cejko, B.I.; Sarosiek, B.; Krejszeff, S.; Judycka, S.; Szczepkowski, M.; Szczepkowska, B.; Kowalski, R. Effects of different stripping methods of female and activation medium on fertilization success in northern pike (Esox lucius). Czech J. Anim. Sci. 2016, 61, 481–486. [Google Scholar] [CrossRef] [Green Version]
- Kujawa, R.; Kucharczyk, D.; Mamcarz, A. A model system for keeping spawners of wild and domestic fish before artificial spawning. Aquac. Eng. 1999, 20, 85–89. [Google Scholar] [CrossRef]
- Luczynski, M.; Dabrowski, K.; Kucharczyk, D.; Glogowski, J.; Luczynski, M.; Szczerbowski, A.; Babiak, I. Gynogenesis in northern pike: UV-inactivation of spermatozoa and the heat shock inducing meiotic diploidization. Environ. Biotechnol. 2007, 3, 39–43. [Google Scholar]
- Billard, R. Utilisation d’un système tris-glycocolle pour tamponner le dilueur d’insémination pour truite. Bull. Fr. Pêche Piscic. 1977, 264, 102–112. [Google Scholar] [CrossRef]
- Woynarovich, E.; Woynarovich, A. Modified technology for elimination of stickness of common carp Cyprius carpio eggs. Aquacult. Hun. (Szarvas) 1980, 2, 19–21. [Google Scholar]
- Billard, R. Spermatogenesis and spermatology of some teleost fish species. Reprod. Nutr. Dev. 1986, 26, 877–920. [Google Scholar] [CrossRef]
- Luczynski, M.J.; Glogowski, J.; Kucharczyk, D.; Luczynski, M.; Demska-Zakes, K. Gynogenesis in northern pike (Esox lucius L.) induced by heat shock–preliminary data. Pol. Arch. Hydrobiol. 1997, 44, 25–32. [Google Scholar]
- Nowosad, J.; Żarski, D.; Biłas, M.; Dryl, K.; Krejszeff, S.; Kucharczyk, D. Dynamics of ammonia excretion in juvenile common tench, Tinca tinca (L.), during intensive rearing under controlled conditions. Aquac. Int. 2013, 21, 629–637. [Google Scholar] [CrossRef]
- Billard, R. Artificial insemination of the trout, Salmo gairdneri Richardson. IV. Effects of K+ and Na+ ions on gamete fertilizing ability. Bull. Fr. Piscic. 1974, 256, 88–100. [Google Scholar]
- Billard, R.; Debruille, M.; Gerard, J.-P.; De Montalembert, G. L’insémination artificielle du brochet. Bull. Fr. Pêche Piscic. 1976, 262, 30–34. [Google Scholar] [CrossRef] [Green Version]
- Reniets, J.P.; Millard, J.L. Use of saline solutions to improve fertilization of Northern pike (Esox lucius) eggs. Progress.-Fish-Cult. 1985, 49, 117–119. [Google Scholar] [CrossRef]
- Żarski, D.; Horváth, Á.; Kotrik, L.; Targońska, K.; Palińska, K.; Krejszeff, S.; Bokor, Z.; Urbányi, B.; Kucharczyk, D. Effect of different activating solutions on the fertilization ability of Eurasian perch, Perca fluviatilis L., eggs. J. Appl. Ichthyol. 2012, 28, 967–972. [Google Scholar] [CrossRef]
- Żarski, D.; Horváth, A.; Held, J.A.; Kucharczyk, D. Artificial Reproduction of Percid Fishes. In Biology and Culture of Percid Fishes; Kestemont, P., Dabrowski, K., Summerfelt, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 123–161. [Google Scholar] [CrossRef]
- Szabó, T.; Urbányi, B.; Müller, T.; Szabó, R.; Horváth, L. Assessment of induced breeding of major Chinese carps at a large-scale hatchery in Hungary. Aquac. Rep. 2019, 14, 100193. [Google Scholar] [CrossRef]
- Brzuska, E. Reproduction effectiveness of carp (Cyprinus carpio L.) from the Hungarian W breeding line after stimulating ovulation with spawning inducing agents of natural (CPH, hCG, PMSG) and/or synthetic origin (Ovopel, Dagin, Ovaprim, mGnRH-a). Aquaculture 2021, 532, 736023. [Google Scholar] [CrossRef]
- Cejko, B.I.; Brzuska, E. Differences in the effectiveness of reproduction among ten breeding strains of the common carp Cyprinus carpio L. after stimulation of ovulation with carp pituitary homogenate or [(D-Ala6, Pro9NEt) mGnRH-a + metoclopramide] (Ovopel). Aquac. Res. 2017, 48, 2221–2230. [Google Scholar] [CrossRef]
- Luczynski, M.J.; Szczerbowski, A.; Kucharczyk, D.; Babiak, I. Preliminary observations on inducing homozygous gynogenesis in northern pike (Esox lucius) using pressure shock. Arch. Pol. Fish. 2004, 12, 87–91. [Google Scholar]
- Samarin, A.M.; Blecha, M.; Uzhytchak, M.; Bytyutskyy, D.; Zarski, D.; Flajshans, M.; Policar, T. Post-ovulatory and post-stripping oocyte ageing in northern pike, Esox lucius (Linnaeus, 1758), and its effect on egg viability rates and the occurrence of larval malformations and ploidy anomalies. Aquaculture 2016, 450, 431–438. [Google Scholar] [CrossRef]
- Żarski, D.; Bokor, Z.; Kotrik, L.; Urbanyi, B.; Horváth, A.; Targońska, K.; Krejszeff, S.; Palińska, K.; Kucharczyk, D. A new classification of a preovulatory oocyte maturation stage suitable for the synchronization of ovulation in controlled reproduction of Eurasian perch, Perca fluviatilis L. Reprod. Biol. 2011, 11, 194–209. [Google Scholar] [CrossRef]
- Kristan, J.; Samarin, A.M.; Malinovskyi, O.; Policar, T. Gamete management for artificial reproduction of northern pike Esox lucius (Linnaeus, 1758). Aquaculture 2020, 528, 735575. [Google Scholar] [CrossRef]
- Wojtczak, M.; Kowalski, R.; Dobosz, S.; Goryczko, K.; Kuźminski, H.; Glogowski, J.; Ciereszko, A. Assessment of water turbidity for evaluation of rainbow trout (Oncorhynchus mykiss) egg quality. Aquaculture 2004, 242, 617–624. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Medium | Composition of Diluent | pH | Author |
|---|---|---|---|
| TW | Tap water (T) | 7.35 | |
| DW | Deionized water (D) | 6.37 | |
| NaCl5-D | 86 mM NaCl, water D | 6.05 | |
| NaCl5-T | 86 mM NaCl, water T | 7.30 | |
| NaCl6-D | 103 mM NaCl, water D | 6.07 | |
| NaCl6-T | 103 mM NaCl, water T | 7.27 | |
| B1-D | 94 mM NaCl, 20 mM Tris, 50 mM glicine, water D | 8.73 | Billard (1977) [44] |
| B1-T | 94 mM NaCl, 20 mM Tris, 50 mM glicine, water T | 8.62 | |
| V-D | 68 mM NaCl, 50 mM urea, water D | 5.55 | Woynarowich and Woynarowich (1980) [45] |
| V-T | 68 mM NaCl, 50 urea, water T | 7.00 |
| Medium | Composition of Diluent | pH | Author |
|---|---|---|---|
| TW | Tap water (T) | 7.35 | |
| DW | Deionized water (D) | 6.37 | |
| NaCl7-D | 120 mM NaCl, water D | 5.40 | Babiak et al. (1997) [40] |
| NaCl7-T | 120 mM NaCl, water T | 7.00 | |
| B2-D | 125 mM NaCl, 20 mM TRIS, 30 mM glicine, 1 mM CaCl2, water D | 8.86 | Billard (1986) [46] |
| B2-T | 125 mM NaCl, 20 mM TRIS, 30 mM glicine, 1 mM CaCl2, water T | 8.73 | |
| B1-D | 94 mM NaCl, 20 mM Tris, 50 mM glicine, water D | 8.73 | Billard (1977) [44] |
| B1-T | 94 mM NaCl, 20 mM Tris, 50 mM glicine, water T | 8.62 | |
| V-D | 68 mM NaCl, 50 mM urea, water D | 5.55 | Woynarowich and Woynarowich (1980) [45] |
| V-T | 68 mM NaCl, 50 mM urea, water T | 7.00 |
| Hatchery/Group | Pasłęk | Pasym | Sterławki |
|---|---|---|---|
| HW | 84.2 ± 2.2 a | 57.8 ± 3.3 b | 60.3 ± 2.9 b |
| V | 0.0 | 26.3 ± 3.6 c | 73.2 ± 2.2 a |
| B1 | 84.5 ± 1.9 a | 72.9 ± 2.9 a | 64.1 ± 3.5 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łuczyński, M.J.; Nowosad, J.; Łuczyńska, J.; Kucharczyk, D. Effect of Application of Different Activation Media on Fertilization and Embryo Survival of Northern Pike, (Esox lucius) under Hatchery Conditions. Animals 2022, 12, 1022. https://doi.org/10.3390/ani12081022
Łuczyński MJ, Nowosad J, Łuczyńska J, Kucharczyk D. Effect of Application of Different Activation Media on Fertilization and Embryo Survival of Northern Pike, (Esox lucius) under Hatchery Conditions. Animals. 2022; 12(8):1022. https://doi.org/10.3390/ani12081022
Chicago/Turabian StyleŁuczyński, Marek J., Joanna Nowosad, Joanna Łuczyńska, and Dariusz Kucharczyk. 2022. "Effect of Application of Different Activation Media on Fertilization and Embryo Survival of Northern Pike, (Esox lucius) under Hatchery Conditions" Animals 12, no. 8: 1022. https://doi.org/10.3390/ani12081022