
Using Osmotic Pumps to Induce the Production of Gametes in Male and Female European Eels

 , and
, and 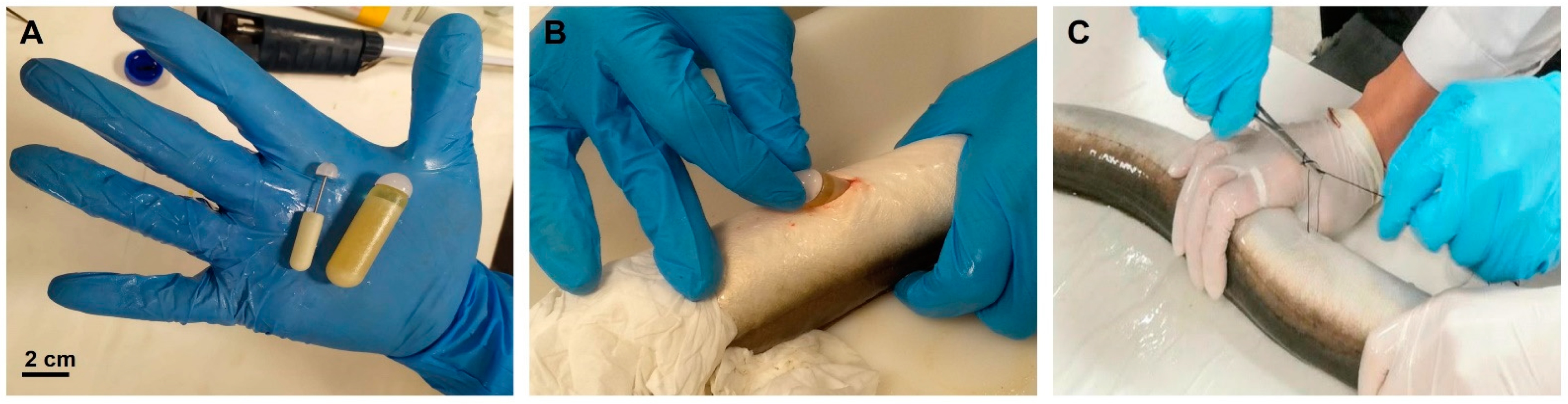{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Handling
2.2. Males
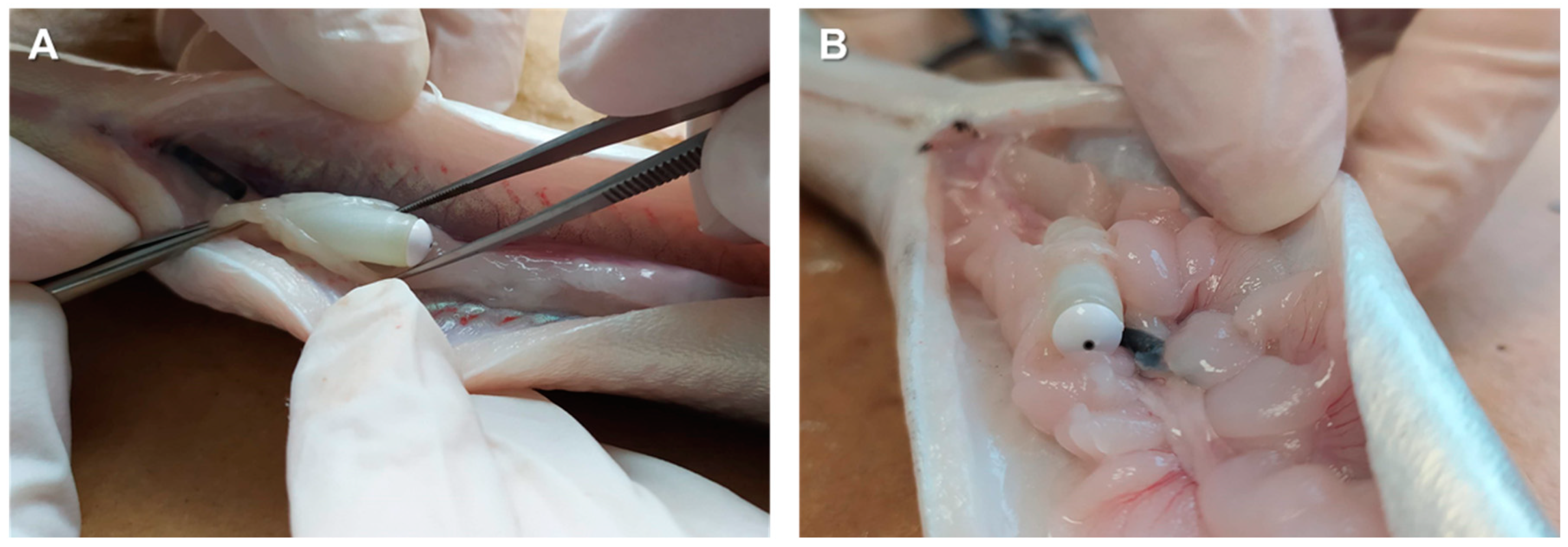
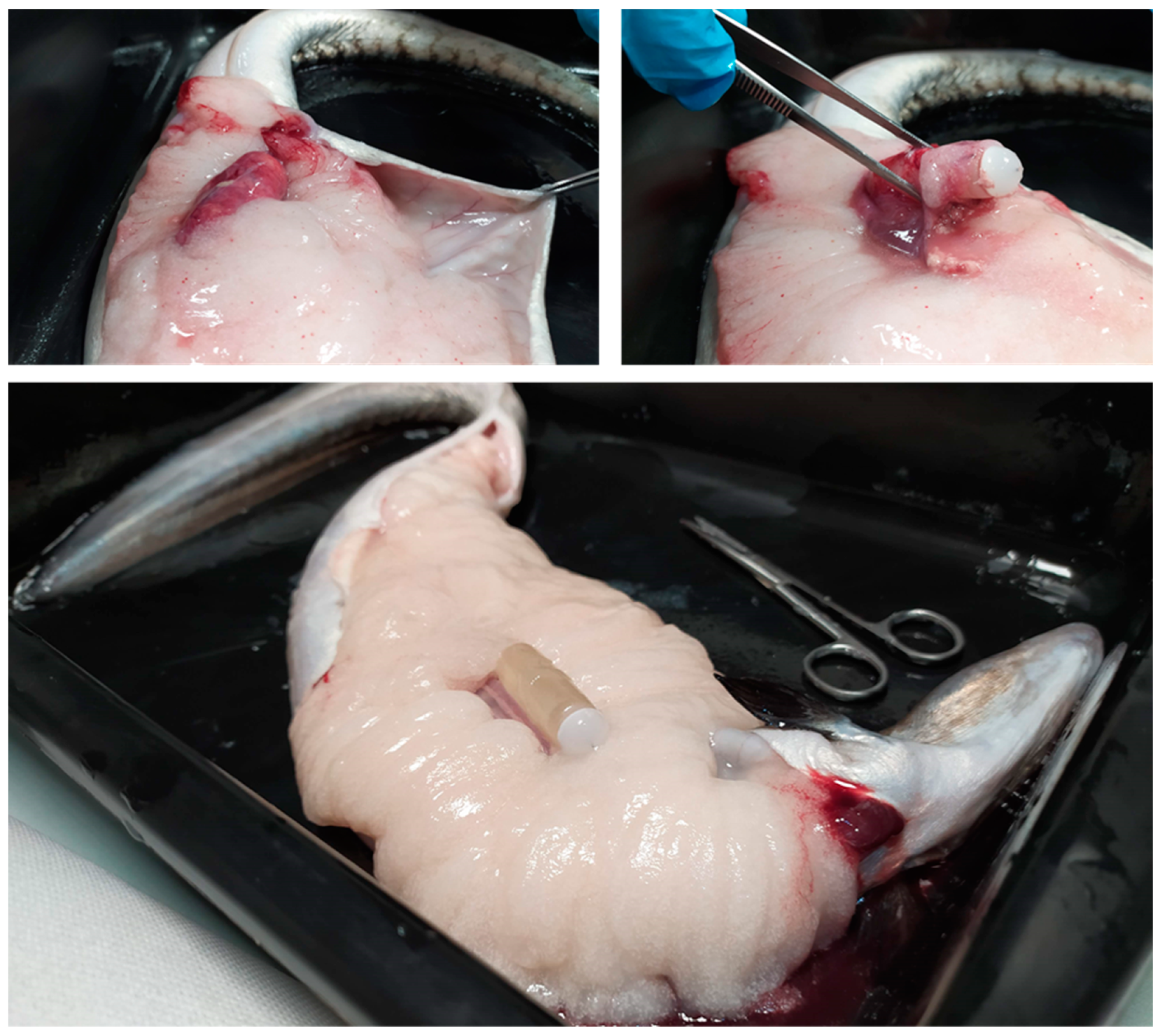
2.2.1. Induction of Sexual Maturation: Experimental Design
2.2.2. Sperm Collection and Sampling
2.2.3. Sperm Motility Assessment
2.2.4. Biometric Parameters
2.3. Females
2.3.1. Induction of Sexual Maturation: Experimental Design
2.3.2. Sexual Maturation Assessment and Spawning Induction
2.3.3. Biometric Parameters: Gonadosomatic Index (GSI)
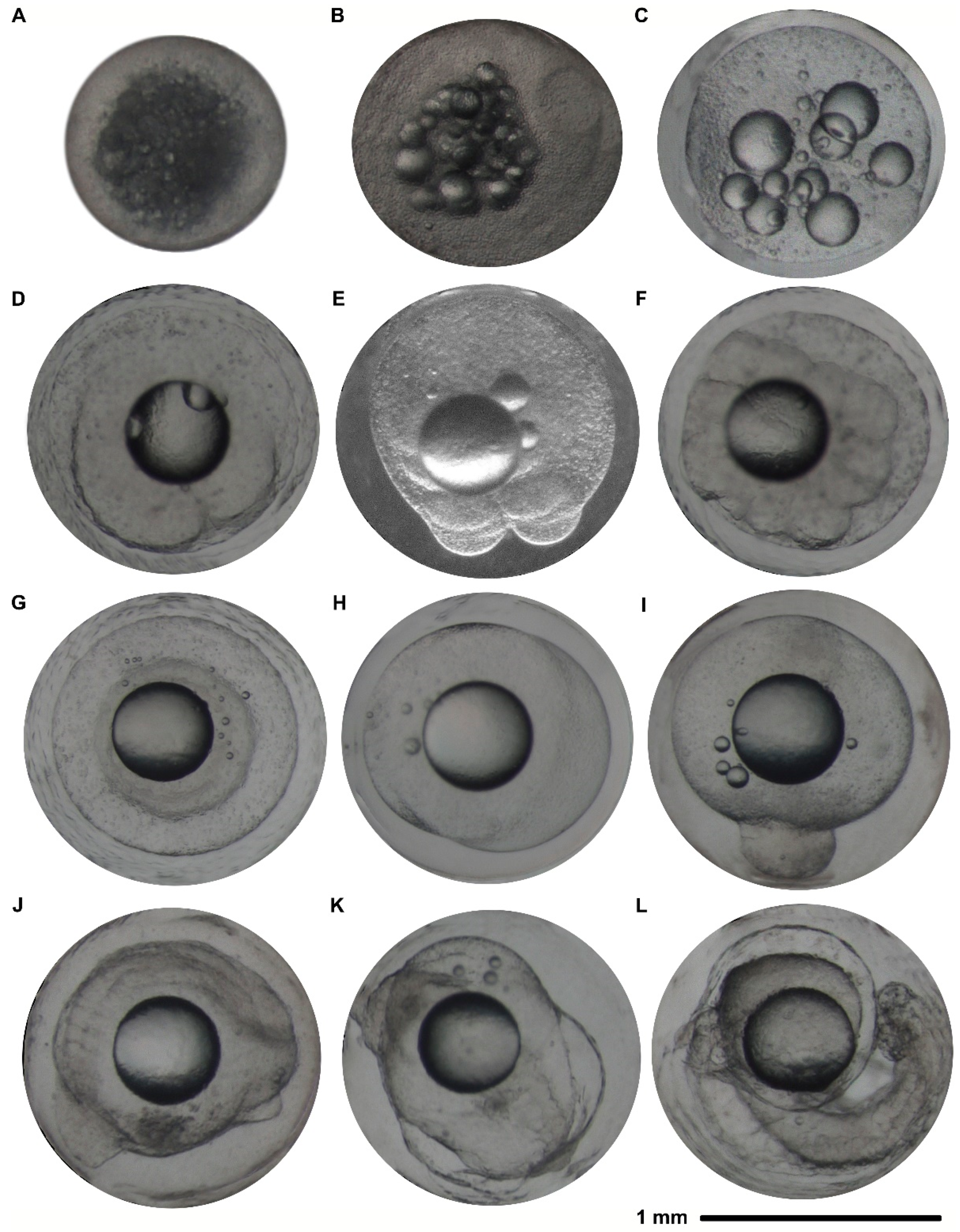
2.3.4. Fertilization, Hatching Rate, and Embryonic Development
2.4. Statistical Analysis
3. Results
3.1. Males
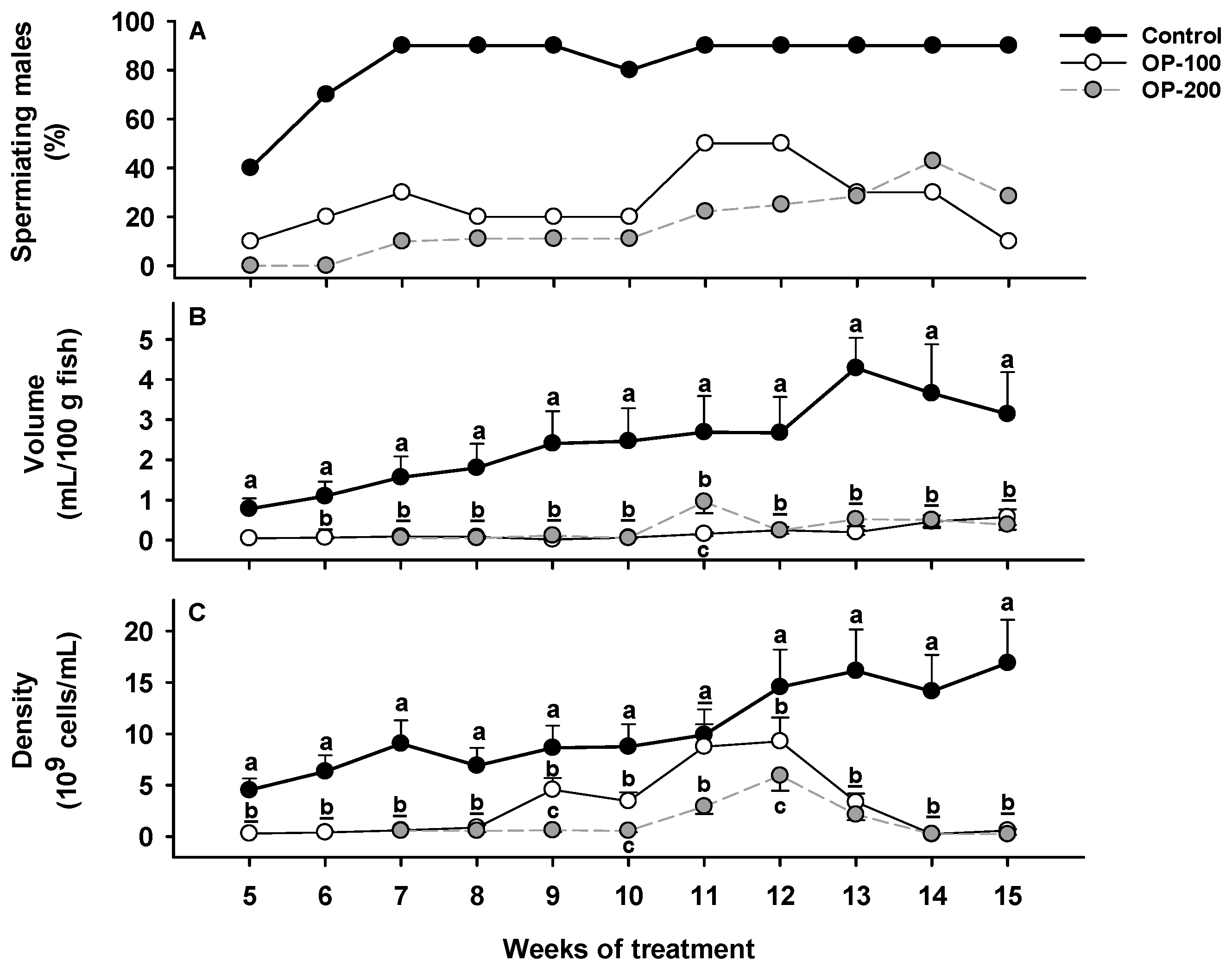
3.1.1. Sperm Production: Spermiating Males, Sperm Volume, and Sperm Density
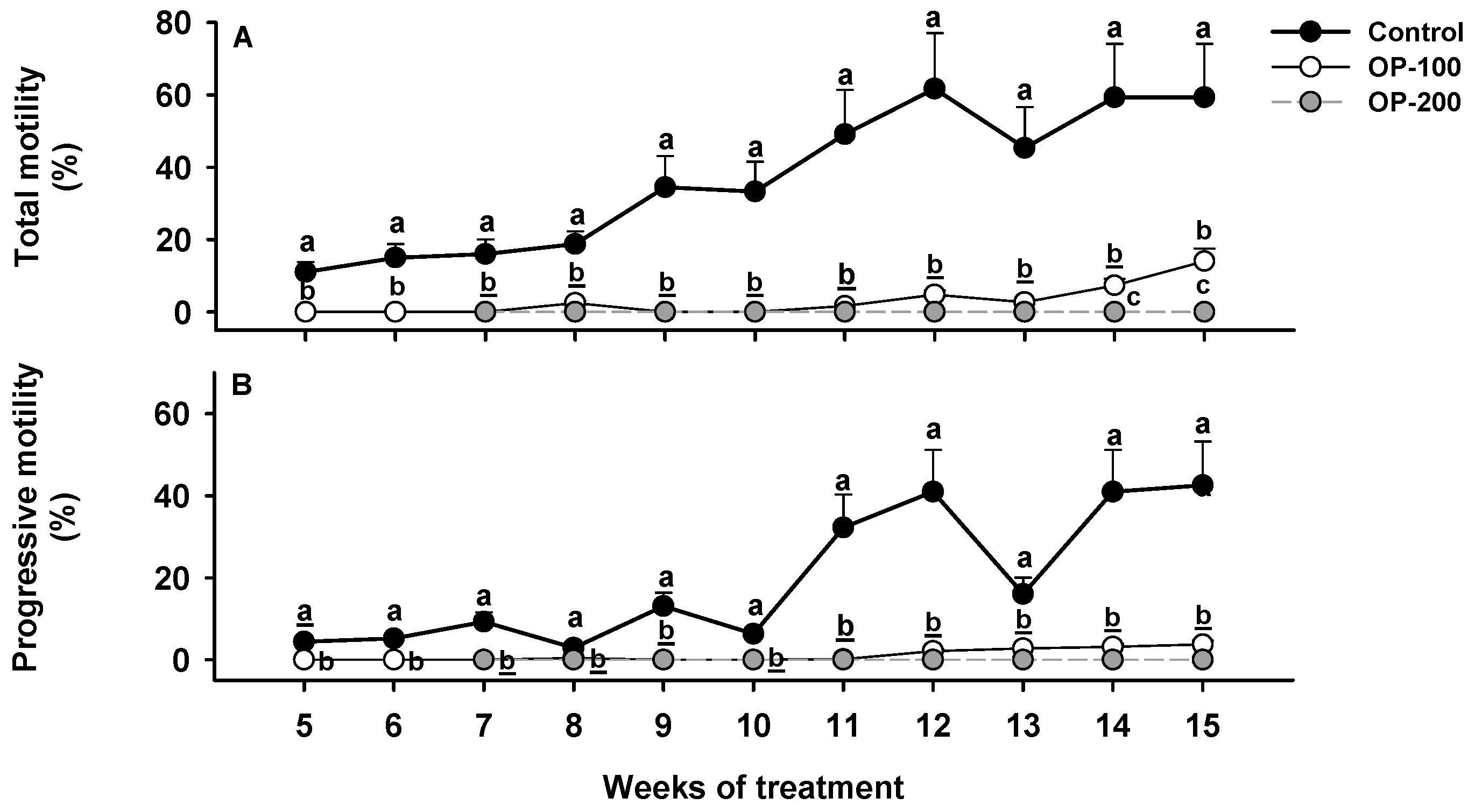
3.1.2. Sperm Quality Assessment: Total Motility and Progressive Motility
3.1.3. Biometric Parameters
3.2. Females
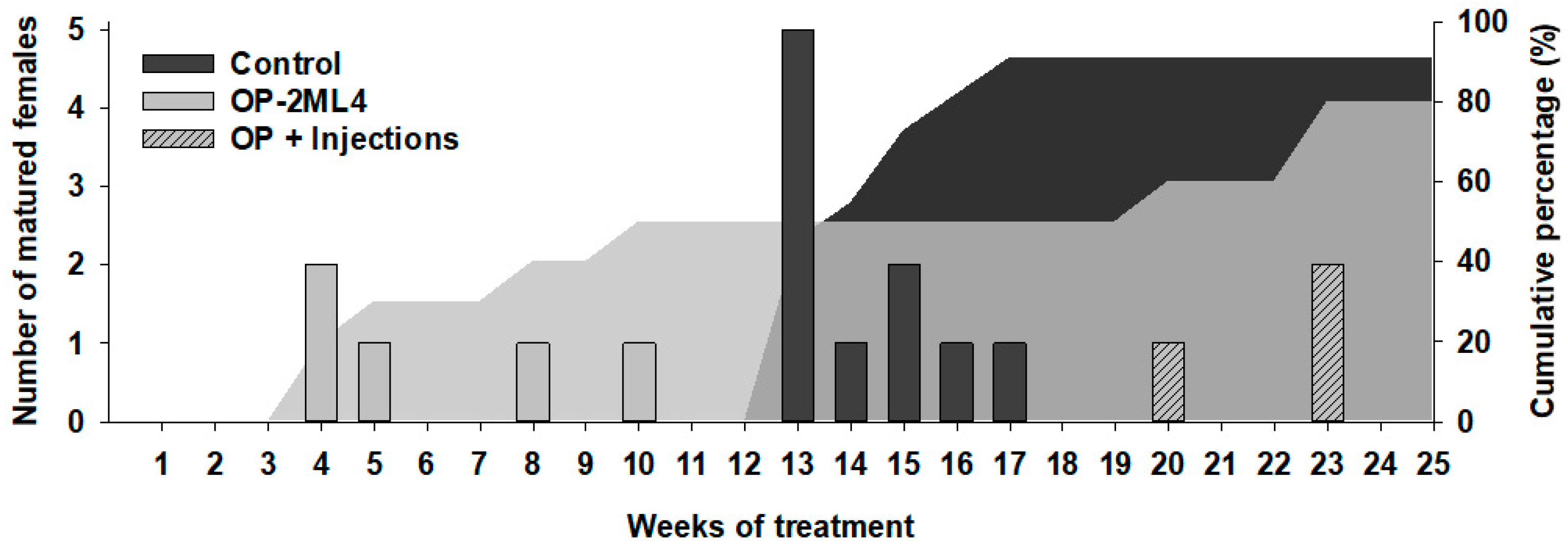
3.2.1. Sexual Maturation Evaluation
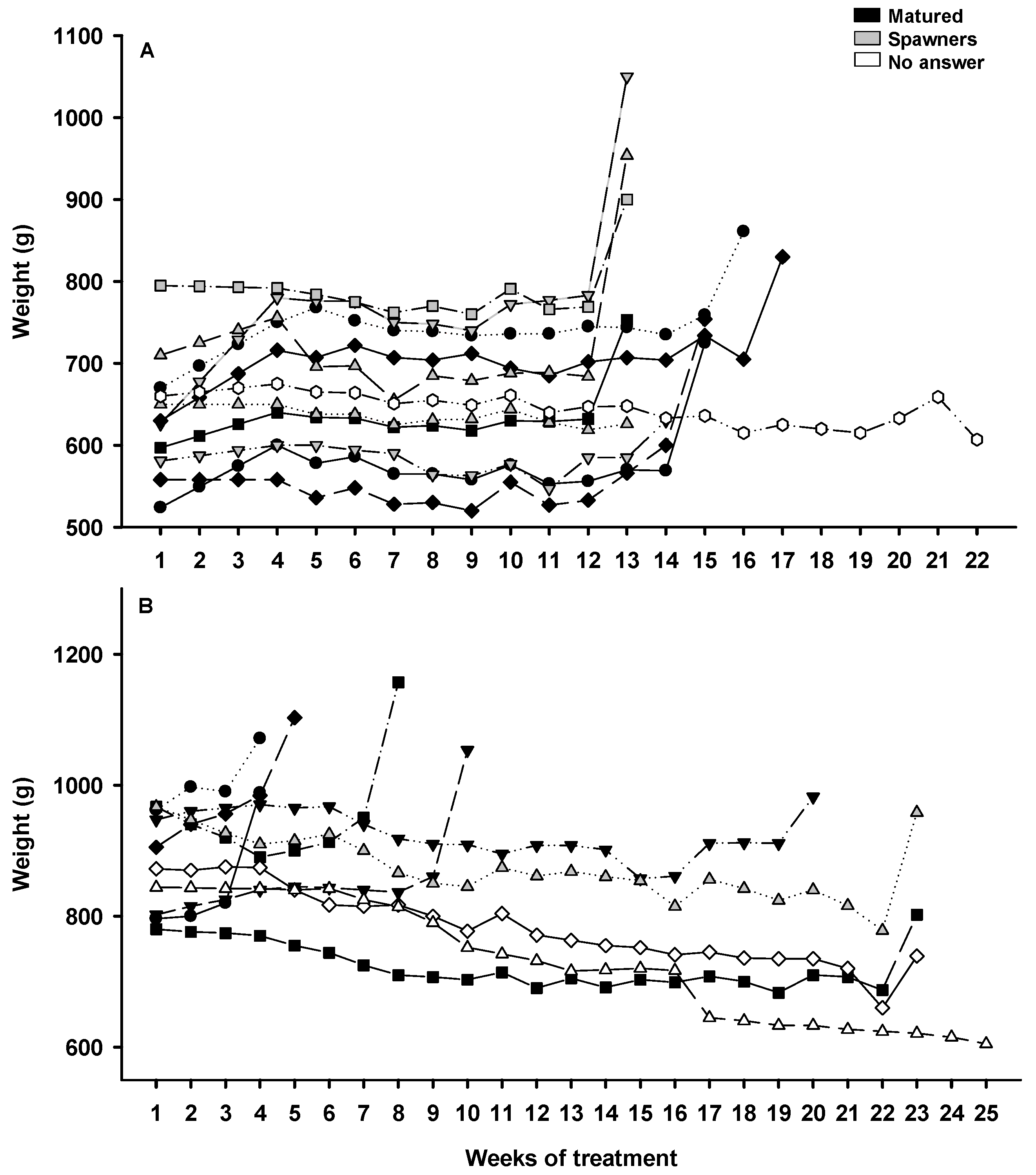
3.2.2. Weight Evolution during Sexual Maturation
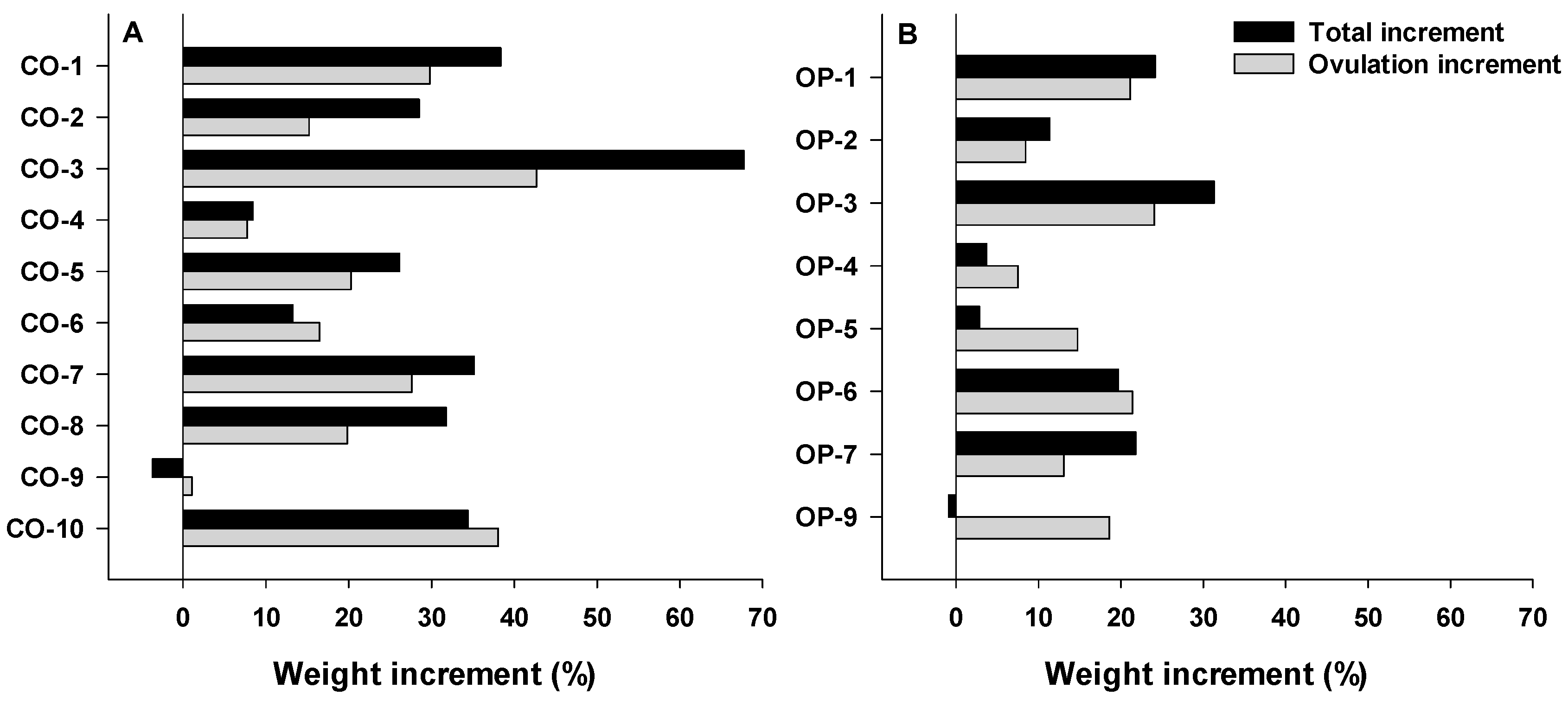
3.2.3. Biometric Parameter: Gonadosomatic Index (GSI)
3.2.4. Fertilization, Hatching Rate, and Embryonic Development
4. Discussion
4.1. Males
4.2. Females
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- ICES. European eel (Anguilla anguilla) throughout its natural range. In ICES Advice of Fishing Opportunities, Catch, and Effort Ecoregions in the Northeast Atlantic; ICES: Copenhagen, Denmark, 2017. [Google Scholar] [CrossRef]
- van den Thillart, G.E.E.J.M.; Dufour, S. How to estimate the reproductive success of European silver eels. In Spawning Migration of the European eel: Reproduction Index, a Useful Tool for Conservation Management; van den Thillart, G.E.E.J.M., Dufour, S., Rankin, J.C., Eds.; Springer: New York, NY, USA, 2009; Volume 30, pp. 3–12. [Google Scholar]
- Pike, C.; Crook, V.; Gollock, M. Anguilla anguilla. The IUCN Red List of Threatened Species. 2020. e.T60344A152845178. Available online: https://www.iucnredlist.org/species/60344/152845178 (accessed on 7 December 2021). [CrossRef]
- Asturiano, J.F. Improvements on the reproductive control of the European eel. In Reproduction in Aquatic Animals: From Basic Biology to Aquaculture Technology; Yoshida, M., Asturiano, J.F., Eds.; Springer: New York, NY, USA, 2020; pp. 293–320. [Google Scholar] [CrossRef]
- Pedersen, B.H. Fertilisation of eggs, rate of embryonic development and hatching following induced maturation of the European eel Anguilla anguilla. Aquaculture 2004, 237, 461–473. [Google Scholar] [CrossRef]
- Tomkiewicz, J.; Kofoed, T.M.N.; Pedersen, J.S. Assessment of testis development during induced spermatogenesis in the European eel Anguilla anguilla. Mar. Coast. Fish. 2011, 3, 106–118. [Google Scholar] [CrossRef] [Green Version]
- Asturiano, J.F.; Pérez, L.; Garzón, D.L.; Peñaranda, D.S.; Marco-Jiménez, F.; Martínez-Llorens, S.; Tomás, A.; Jover, M. Effect of different methods for the induction of spermiation on semen quality in European eel. Aquac. Res. 2005, 36, 1480–1487. [Google Scholar] [CrossRef]
- Koumpiadis, P.; Sganga, D.E.; Politis, S.N.; Gallego, V.; Butts, I.A.E.; Asturiano, J.F.; Batjakas, I.E.; Tomkiewicz, J. Sperm production and quality in European eel (Anguilla anguilla) in relation to hormonal treatment. Reprod. Domest. Anim. 2021, 56, 1497–1505. [Google Scholar] [CrossRef]
- Gallego, V.; Mazzeo, I.; Vílchez, M.C.; Peñaranda, D.S.; Carneiro, P.C.F.; Pérez, L.; Asturiano, J.F. Study of the effects of thermal regime and alternative hormonal treatments on the reproductive performance of European eel males (Anguilla anguilla) during induced sexual maturation. Aquaculture 2012, 354–355, 7–16. [Google Scholar] [CrossRef]
- Herranz-Jusdado, J.G.; Rozenfeld, C.; Morini, M.; Pérez, L.; Asturiano, J.F.; Gallego, V. Recombinant vs purified mammal gonadotropins as maturation hormonal treatments of European eel males. Aquaculture 2019, 501, 527–536. [Google Scholar] [CrossRef]
- Palstra, A.P.; Cohen, E.G.H.; Niemantsverdriet, P.R.W.; van Ginneken, V.J.T.; van den Thillart, G.E.E.J.M. Artificial maturation and reproduction of European silver eel: Development of oocytes during final maturation. Aquaculture 2005, 249, 533–547. [Google Scholar] [CrossRef]
- Butts, I.A.E.; Sørensen, S.R.; Politis, S.N.; Pitcher, T.E.; Tomkiewicz, J. Standardization of fertilization protocols for the European eel, Anguilla anguilla. Aquaculture 2014, 426–427, 9–13. [Google Scholar] [CrossRef]
- Pérez, L.; Peñaranda, D.S.; Dufour, S.; Baloche, S.; Palstra, A.P.; van den Thillart, G.E.E.J.M.; Asturiano, J.F. Influence of temperature regime on endocrine parameters and vitellogenesis during experimental maturation of European eel (Anguilla anguilla) females. Gen. Comp. Endocrinol. 2011, 174, 51–59. [Google Scholar] [CrossRef]
- Mordenti, O.; Di Biase, A.; Bastone, G.; Sirri, R.; Zaccaroni, A.; Parmeggiani, A. Controlled reproduction in the wild European eel (Anguilla anguilla): Two populations compared. Aquac. Int. 2013, 21, 1045–1063. [Google Scholar] [CrossRef]
- Brydges, N.M.; Boulcott, P.; Ellis, T.; Braithwaite, V.A. Quantifying stress responses induced by different handling methods in three species of fish. Appl. Anim. Behav. Sci. 2009, 116, 295–301. [Google Scholar] [CrossRef]
- Schreck, C.B.; Contreras-Sanchez, W.; Fitzpatrick, M.S. Effects of stress on fish reproduction, gamete quality, and progeny. Aquaculture 2001, 197, 3–24. [Google Scholar] [CrossRef]
- Mazzeo, I.; Peñaranda, D.S.; Gallego, V.; Baloche, S.; Nourizadeh-Lillabadi, R.; Tveiten, H.; Dufour, S.; Asturiano, J.F.; Weltzien, F.-A.; Pérez, L. Temperature modulates the progression of vitellogenesis in the European eel. Aquaculture 2014, 434, 38–47. [Google Scholar] [CrossRef]
- Burgerhout, E.; Lokman, P.M.; van den Thillart, G.E.E.J.M.; Dirks, R.P. The time-keeping hormone melatonin: A possible key cue for puberty in freshwater eels (Anguilla spp.). Rev. Fish Biol. Fisheries 2019, 29, 1–21. [Google Scholar] [CrossRef]
- Mylonas, C.C.; Duncan, N.J.; Asturiano, J.F. Hormonal manipulations for the enhancement of sperm production in cultured fish and evaluation of sperm quality. Aquaculture 2017, 472, 21–44. [Google Scholar] [CrossRef]
- Matejkova, J.; Podhorec, P. Sustained drug delivery system in fish and the potential for use of PLGA microparticles: A review. Vet. Med. 2019, 64, 287–293. [Google Scholar] [CrossRef]
- Marte, C.L.; Sherwood, N.M.; Crim, L.W.; Harvey, B. Induced spawning of maturing milkfish (Chanos chanos Forsskal) with gonadotropin-releasing hormone (GnRH) analogues administered in various ways. Aquaculture 1987, 60, 303–310. [Google Scholar] [CrossRef]
- Almendras, J.M.; Dueñas, C.; Nacario, J.; Sherwood, N.M.; Crim, L. Sustained hormone release. III. Use of gonadotropin releasing hormone analogues to induce multiple spawnings in sea bass, Lates calcarifer. Aquaculture 1988, 74, 97–111. [Google Scholar] [CrossRef]
- Murugananthkumar, R.; Prathibha, Y.; Senthilkumaran, B.; Rajakumar, A.; Kagawa, H. In vivo induction of human chorionic gonadotropin by osmotic pump advances sexual maturation during pre-spawning phase in adult catfish. Gen. Comp. Endocrinol. 2017, 251, 74–84. [Google Scholar] [CrossRef]
- Kagawa, H.; Kasuga, Y.; Adachi, J.; Nishi, A.; Hashimoto, H.; Imaizumi, H.; Kaji, S. Effects of continuous administration of human chorionic gonadotropin, salmon pituitary extract, and gonadotropin-releasing hormone using osmotic pumps on induction of sexual maturation in male Japanese eel, Anguilla japonica. Aquaculture 2009, 296, 117–122. [Google Scholar] [CrossRef]
- Kagawa, H.; Fujie, N.; Imaizumi, H.; Masuda, Y.; Oda, K.; Adachi, J.; Nishi, A.; Hashimoto, H.; Teruya, K.; Kaji, S. Using osmotic pumps to deliver hormones to induce sexual maturation of female Japanese eels, Anguilla japonica. Aquaculture 2013, 388–391, 30–34. [Google Scholar] [CrossRef]
- Irtamar—Irta. Available online: https://irtamar.irta.cat/ (accessed on 22 December 2021).
- ALZET® Osmotic Pumps. Available online: https://www.alzet.com/ (accessed on 22 December 2021).
- Pérez, L.; Asturiano, J.F.; Tomás, A.; Zegrari, S.; Barrera, R.; Espinós, F.J.; Navarro, J.C.; Jover, M. Induction of maturation and spermiation in the male European eel: Assessment of sperm quality throughout treatment. J. Fish Biol. 2000, 57, 1488–1504. [Google Scholar] [CrossRef]
- Peñaranda, D.S.; Pérez, L.; Gallego, V.; Barrera, R.; Jover, M.; Asturiano, J.F. European eel sperm diluent for short-term storage. Reprod. Domest. Anim. 2010, 45, 407–415. [Google Scholar] [CrossRef]
- Peñaranda, D.S.; Gallego, V.; Rozenfeld, C.; Herranz-Jusdado, J.G.; Pérez, L.; Gómez, A.; Giménez, I.; Asturiano, J.F. Using specific recombinant gonadotropins to induce spermatogenesis and spermiation in the European eel (Anguilla anguilla). Theriogenology 2018, 107, 6–20. [Google Scholar] [CrossRef]
- Okamura, A.; Yamada, Y.; Yokouchi, K.; Horie, N.; Mikawa, N.; Utoh, T.; Tanaka, S.; Tsukamoto, K. A silvering index for the Japanese eel Anguilla japonica. Environ. Biol. Fish 2007, 80, 77–89. [Google Scholar] [CrossRef]
- Kleppe, L.; Karlsen, O.; Edvarsen, R.B.; Norberg, B.; Andersson, E.; Taranger, G.L.; Wargelius, A. Cortisol treatment of prespawning female cod affects cytogenesis related factors in eggs and embryos. Gen. Comp. Endocrinol. 2013, 189, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Khatri, M.; Sudhakumari, C.; Kagawa, H.; Dutta-Gupta, A.; Senthilkumaran, B. Controlled reléase of sex steroids through osmotic pump alters brain GnRH1 and catecholaminergic system dimorphically in the catfish, Clarias gariepinus. Brain Res. Bull. 2020, 164, 325–333. [Google Scholar] [CrossRef]
- Gallego, V.; Asturiano, J.F. Sperm motility in fish: Technical applications and perspectives through CASA-Mot systems. Reprod. Fertil. Dev. 2018, 30, 820–832. [Google Scholar] [CrossRef]
- Rolfe, B.; Mooney, J.; Zhang, B.; Jahnke, S.; Le, S.-J.; Chau, Y.-Q.; Huang, Q.; Wang, H.; Campbell, G.; Campbell, J. The fibrotic response to implanted biomaterials: Implications for tissue engineering. In Regenerative Medicine and Tissue Engineering: Cells and Biomaterials; Eberli, D., Ed.; InTech: Rijeka, Croatia, 2011; pp. 551–569. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, B.H. Induced sexual maturation of the European eel Anguilla anguilla and fertilisation of the eggs. Aquaculture 2003, 224, 323–338. [Google Scholar] [CrossRef]
- Kottmann, J.S.; Jørgensen, M.G.P.; Bertolini, F.; Loh, A.; Tomkiewicz, J. Differential impacts of carp and salmon pituitary extracts on induced oogenesis, egg quality, molecular ontogeny and embryonic development competence in European eel. PLoS ONE 2020, 15, e0235617. [Google Scholar] [CrossRef]
- Mcbride, R.S.; Somarakis, S.; Fitzhugh, G.R.; Albert, A.; Yaragina, N.A.; Wuenschel, M.J.; Alonso-Fernández, A.; Basilone, G. Energy acquisition and allocation to egg production in relation to fish reproductive strategies. Fish Fish. 2015, 16, 23–57. [Google Scholar] [CrossRef] [Green Version]
- Durif, C.M.F.; Dufour, S.; Elie, P. Impact of silvering stage, age, body size and condition on reproductive potential of the European eel. Mar. Ecol. Prog. Ser. 2006, 327, 171–181. [Google Scholar] [CrossRef] [Green Version]
- Mordenti, O.; Di Biase, A.; Sirri, R.; Modugno, S.; Tasselli, A. Induction of sexual maturation in wild female European eels (Anguilla anguilla) in darkness and light. Isr. J. Aquac. 2012, 64, 726–735. [Google Scholar] [CrossRef]
- Ohta, H.; Kagawa, H.; Tanaka, H.; Okuzawa, K.; Hirose, K. Changes in fertilization and hatching rates with time after ovulation induced by 17, 20β-dihydroxy-4-pregnen-3-one in the Japanese eel, Anguilla japonica. Aquaculture 1996, 139, 291–301. [Google Scholar] [CrossRef]
- Tanaka, H.; Kagawa, H.; Ohta, H.; Unuma, T.; Nomura, K. The first production of glass eel in captivity: Fish reproductive physiology facilitates great progress in aquaculture. Fish Physiol. Biochem. 2003, 28, 493–497. [Google Scholar] [CrossRef]
- Tomkiewicz, J.; Støttrup, J.G.; Corraze, G.; Kausik, S.; Holst, L.; Mcevoy, F.; Dufour, S.; Lafont, A.G.; Asturiano, J.F.; Sørensen, S.; et al. Reproduction of European eel and larval culture: State of the art. Commun. Agric. Appl. Biol. Sci. 2013, 78, 455–456. [Google Scholar]
- Mordenti, O.; Casalini, A.; Mandelli, M.; Di Biase, A. A closed recirculating aquaculture system for artificial seed production of the European eel (Anguilla anguilla): Technology development for spontaneous spawning and eggs incubation. Aquac. Eng. 2014, 58, 88–94. [Google Scholar] [CrossRef]
- Jéhannet, P.; Palstra, A.P.; Heinsbroek, L.T.N.; Krujit, L.; Dirks, R.P.; Swinkels, W.; Komen, H. What goes wrong during early development of artificially reproduced European eel Anguilla anguilla? Clues from the larval transcriptome and gene expression patterns. Animals 2021, 11, 1710. [Google Scholar] [CrossRef]
- Politis, S.N.; Butts, I.A.E.; Tomkiewicz, J. Light impacts embryonic and early larval development of the European eel, Anguilla anguilla. J. Exp. Mar. Biol. Ecol. 2014, 461, 407–415. [Google Scholar] [CrossRef]
- Sørensen, S.; Skov, P.; Lauesen, P.; Tomkiewicz, J.; Bossier, P.; de Schryver, P. Microbial interference and potential control in culture of European eel (Anguilla anguilla) embryos and larvae. Aquaculture 2014, 426–427, 1–8. [Google Scholar] [CrossRef]
- Sørensen, S.R.; Tomkiewicz, J.; Munk, P.; Butts, I.A.E.; Nielsen, A.; Lauesen, P.; Graver, C. Ontogeny and growth of early life stages of captive-bred European eel. Aquaculture 2016, 456, 50–61. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanes-García, M.; García-Salinas, P.; Morini, M.; Pérez, L.; Asturiano, J.F.; Gallego, V. Using Osmotic Pumps to Induce the Production of Gametes in Male and Female European Eels. Animals 2022, 12, 387. https://doi.org/10.3390/ani12030387
Blanes-García M, García-Salinas P, Morini M, Pérez L, Asturiano JF, Gallego V. Using Osmotic Pumps to Induce the Production of Gametes in Male and Female European Eels. Animals. 2022; 12(3):387. https://doi.org/10.3390/ani12030387
Chicago/Turabian StyleBlanes-García, Marta, Pablo García-Salinas, Marina Morini, Luz Pérez, Juan F. Asturiano, and Victor Gallego. 2022. "Using Osmotic Pumps to Induce the Production of Gametes in Male and Female European Eels" Animals 12, no. 3: 387. https://doi.org/10.3390/ani12030387