
Preference for Shelters at Different Developmental Stages of Chinese Mitten Crab (Eriocheir sinensis)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
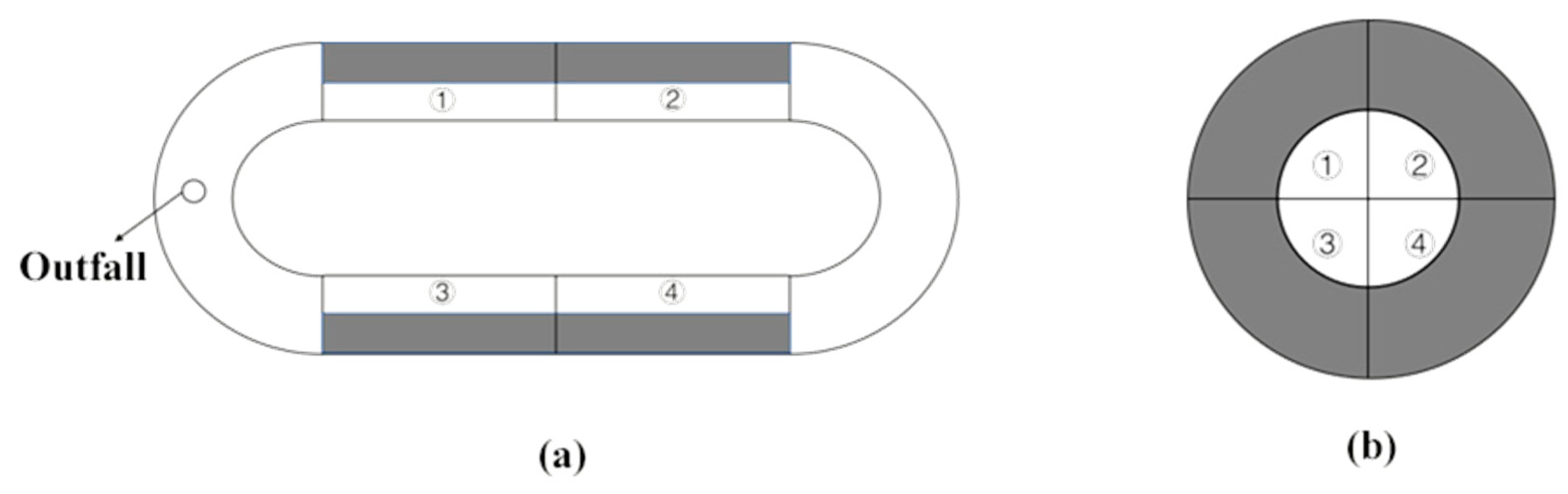
2.1. Animal Collection and Source of Shelters
2.2. Experimental Design and Procedure
2.3. Data Acquisition and Statistical Analysis
3. Results
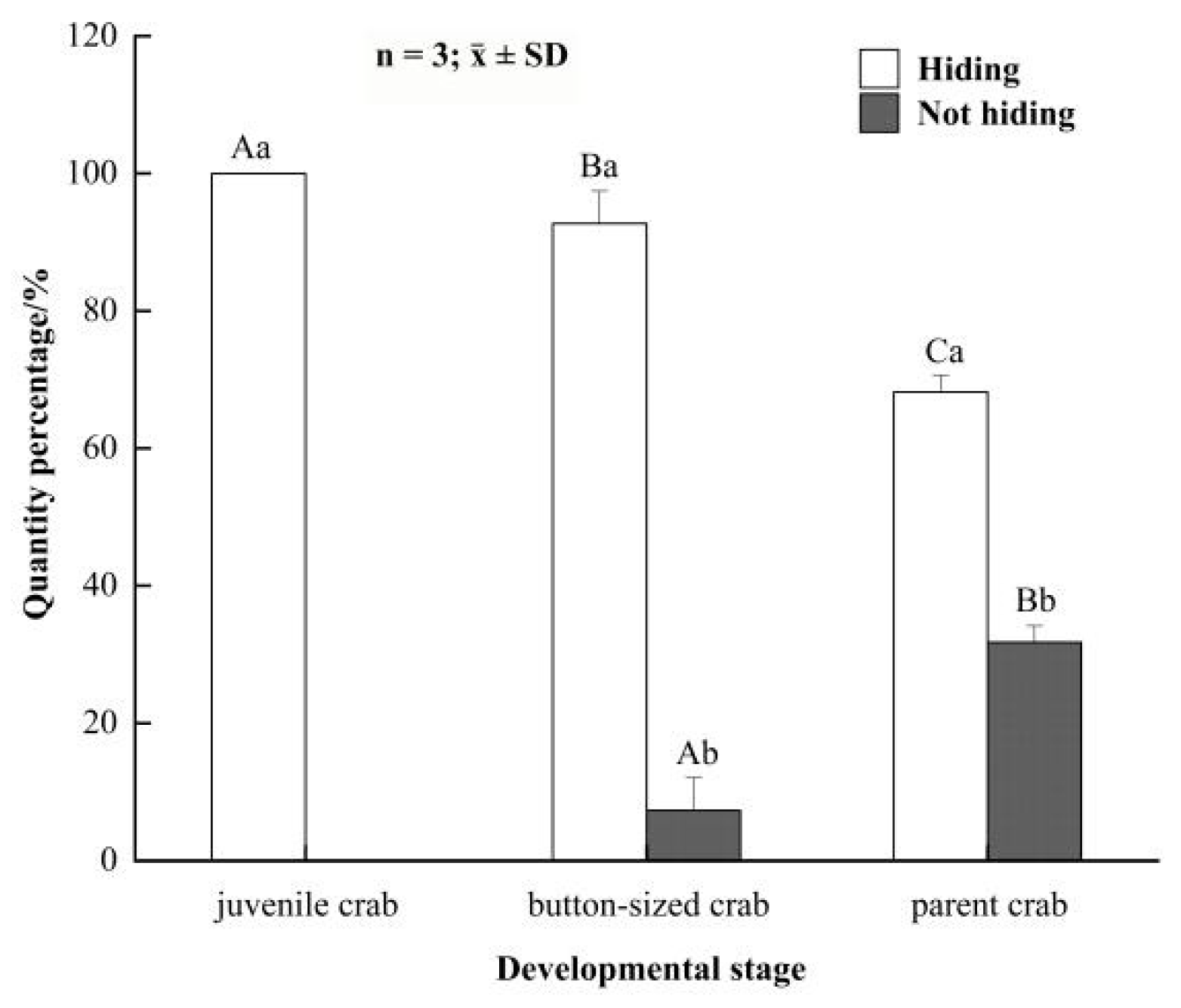
3.1. Concealing Behavior of E. sinensis in Different Developmental Stages
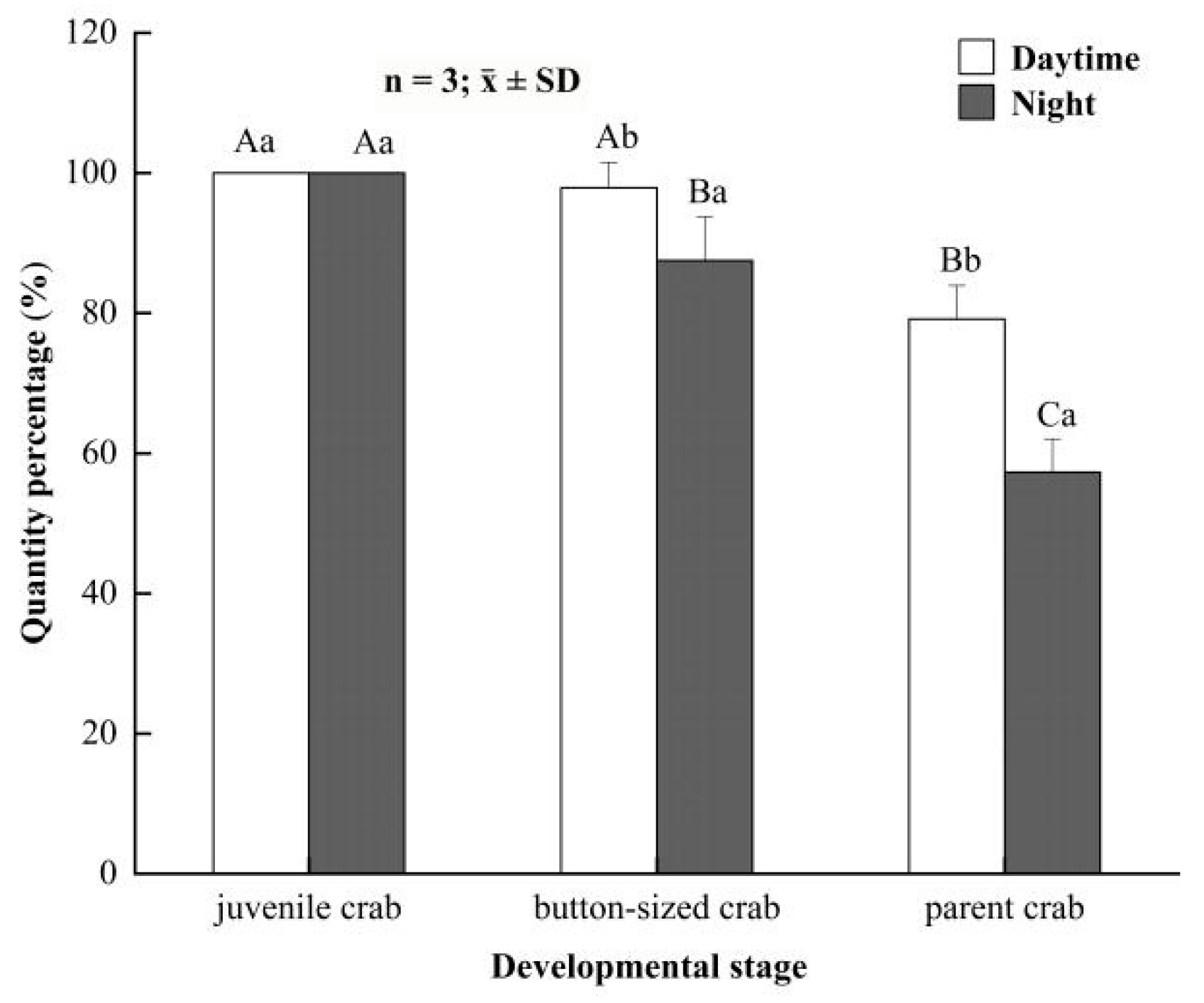
3.2. Differences in Concealing Behavior of E. sinensis between Daytime and Night
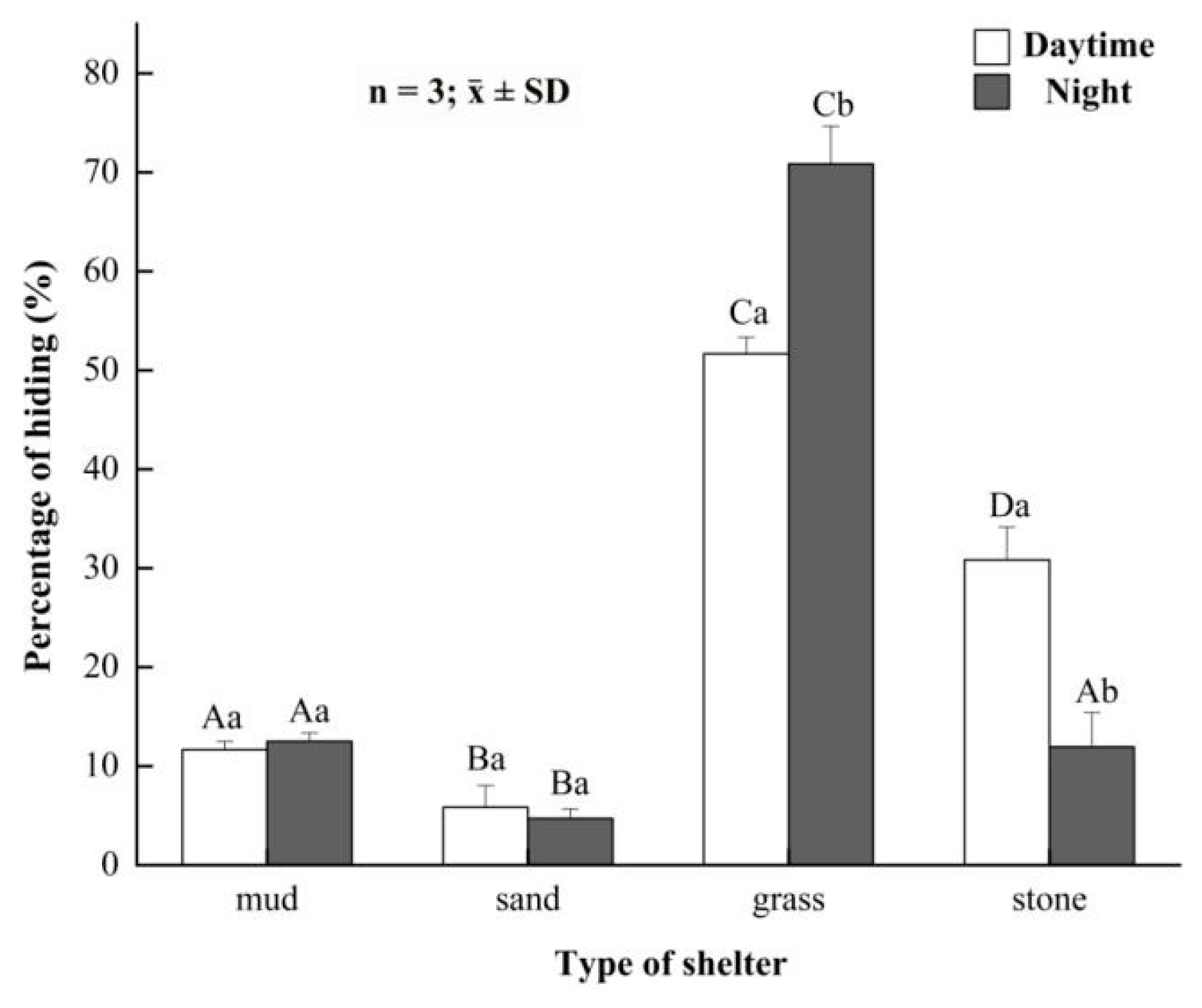
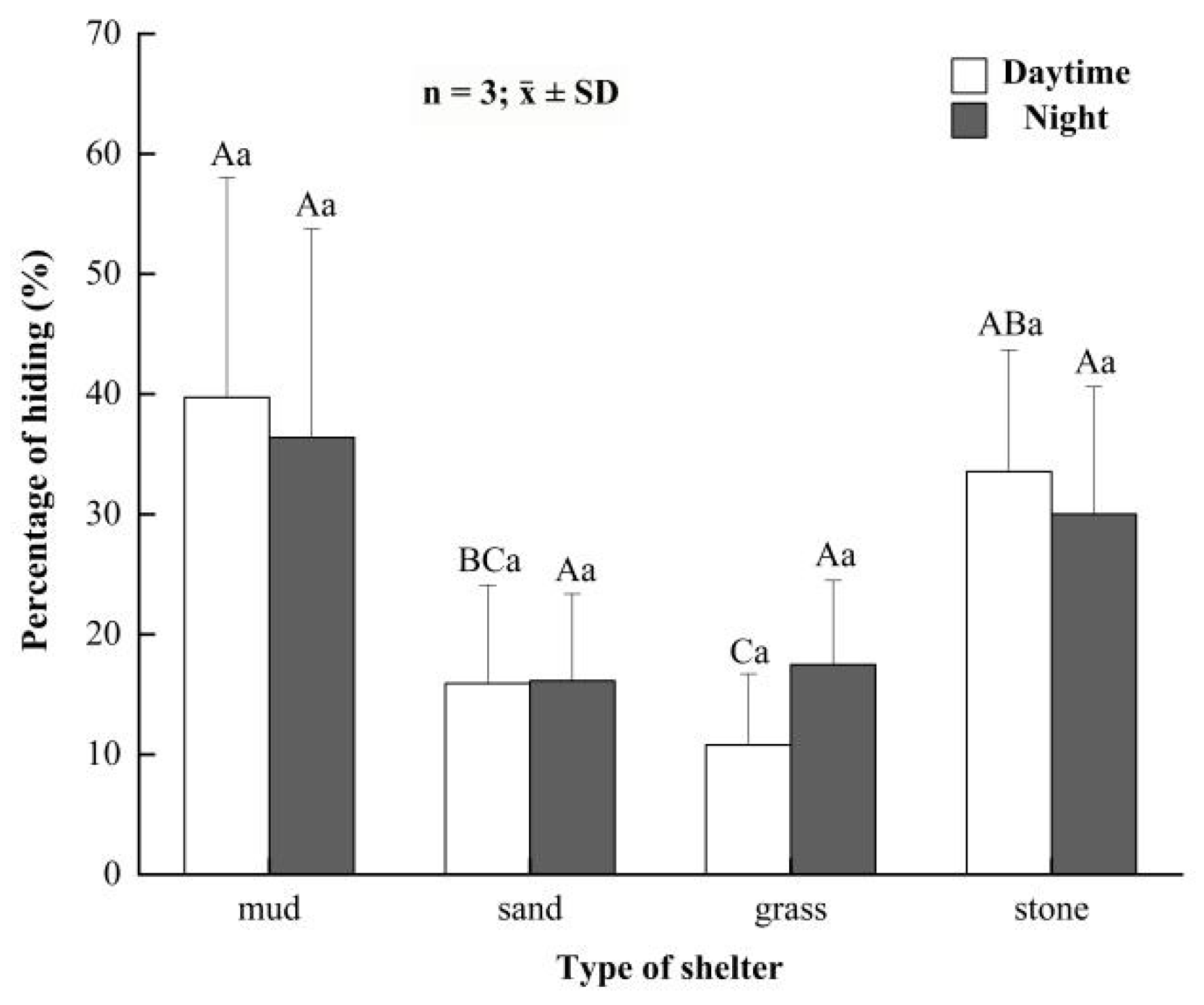
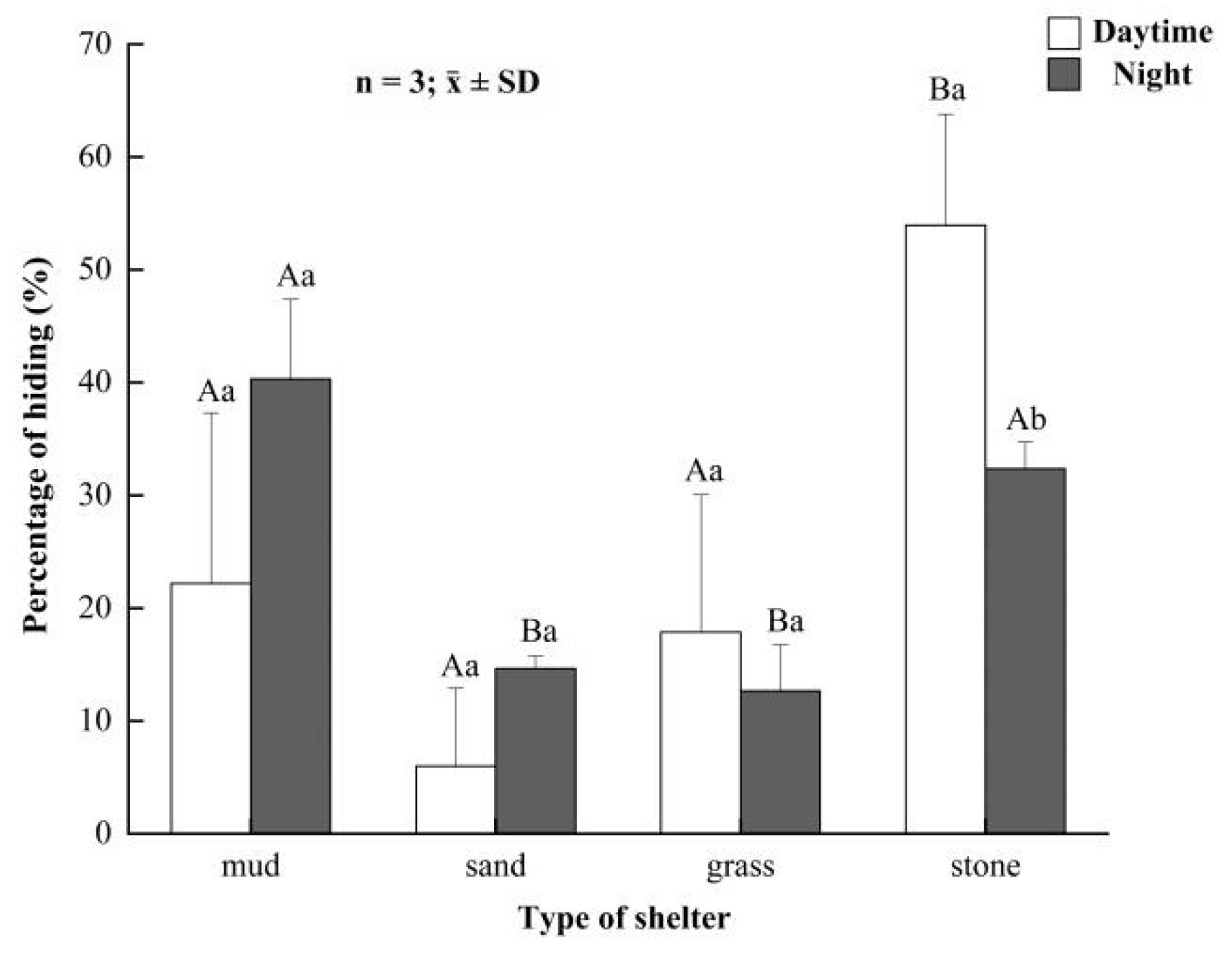
3.3. Preference for Shelters in the Juvenile Crabs
3.4. Preference for Shelters in the Button-Sized Crabs
3.5. Preference for Shelters in the Parent Crabs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feng, G.; Zhuang, P. Stock enhancement and genetic preservation of Chinese mitten crab (Eriocheir sinensis) in the Yangtze River estuary. In Aquaculture in China, Success Stories and Modern Trends; Gui, J., Tang, Q., Li, Z., Liu, J., De Silva, S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2018; pp. 514–526. [Google Scholar]
- Herborg, L.M.; Rushton, S.; Clare, A.; Bentley, M. The invasion of the Chinese mitten crab (Eriocheir sinensis) in the United Kingdom and its comparison to continental Europe. Biol. Invasions 2005, 7, 959–968. [Google Scholar] [CrossRef]
- Gilbey, V.; Attrill, M.; Coleman, R. Juvenile Chinese mitten crabs (Eriocheir sinensis) in the Thames estuary: Distribution, movement and possible interactions with the native crab Carcinus Maenas. Biol. Invasions 2008, 10, 67–77. [Google Scholar] [CrossRef]
- Liu, K.; Duan, J.R.; Xu, D.P.; Zhang, M.Y.; Shi, W.G. Studies on current resource and causes of catch fluctuation of brooders of mitten crab in estuary of the Changjiang River. J. Lake Sci. 2006, 2, 212–217, (In Chinese with English abstract). [Google Scholar]
- Du, N.S. Migration of Eriocheir sinensis. Fish. Sci. Technol. Inf. 2004, 31, 56–57+94. [Google Scholar]
- Huang, B.; Du, N.S. Effects of temperature and Ca2+ on the larval development of the decapoda crustacean: Eriocheir sinensis. Chin. J. Oceanol. Limnol. 2001, 19, 228–232. [Google Scholar]
- He, J.; Gao, Y.; Wang, W.; Xie, J.; Shi, H.; Wang, G.; Xu, W. Limb autotomy patterns in the juvenile swimming crab (Portunus trituberculatus) in earth ponds. Aquaculture 2016, 463, 189–192. [Google Scholar] [CrossRef]
- Sterner, A.W.; Ottmar, M.L.; Copeman, L.A. Temperature effects on the molting, growth, and lipid composition of newly-settled red king crab. J. Exp. Mar. Biol. Ecol. 2010, 393, 138–147. [Google Scholar] [CrossRef]
- Lu, Y.; Wang, F.; Zhao, Z.; Dong, S.; Ma, S. Effects of salinity on growth, molt and energy utilization of juvenile swimming crab Portunus trituberculatus. J. Fish. Sci. China 2012, 13, 237–245. [Google Scholar] [CrossRef]
- Liu, Y.K. Effect of Sand Sediment and Shelters on Behavior, Growth and Survivorship of Portunus Trituberculatus. Master’s Thesis, Ocean University of China, Shandong, China, June 2009. [Google Scholar]
- Zhang, T.; Kong, J.; Liu, P.; Zhang, Q.; Wang, Q. Effect of diet and stocking density on growth and survivorship of juvenile shrimp Fenneropenaeus chinensis. Mar Fish Res. 2008, 29, 41–47, (In Chinese with English abstract). [Google Scholar]
- Daleo, P.; Luppi, T.; Casariego, A.M.; Escapa, M.; Riberiro, P.; Silva, P.; Lribarne, O. The effect of size and cheliped autotomy on sexual competition between males of the mud crab Cyrtograpsus angulatus Dana. Mar Biol. 2009, 156, 269–275. [Google Scholar] [CrossRef]
- Dai, X.L.; Zhou, M.Y.; Ju, B.; Guo, Z.Q.; Jiang, F.; Su, J.; Ding, F.J. Effects of stocking density on growth, sexual differentiation and gonad development of Macrobrachium rosenbergii. J. Fish. China 2016, 40, 1874–1882, (In Chinese with English abstract). [Google Scholar]
- Jiang, Z.H.; Cong, N.; Qian, G.Y. Selection and setting of concealed objects in Macrobrachium rosenbergii breeding pond. Inland Fish. 1997, 6, 25–26. (In Chinese) [Google Scholar]
- Zhang, Q.; Wang, Z.B.; Yang, X.B.; Zhu, S.G. Effects of the coverage of waterweed in pond on the growth of Eriocheir sinensis. Anim. Husb. Feed Sci. 2014, 35, 66–67. [Google Scholar]
- Pile, A.J.; Lipcius, R.N.; Montfrans, J.; Orth, R.J. Density-dependent settler-recruit-juvenile relationships in blue crabs. Ecol. Monogr. 1996, 66, 277–300. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, F.; Song, C.; Gao, Y.; Geng, Z.; Zhuang, P. Effects of stereoscopic artificial floating wetlands on nekton abundance and biomass in the Yangtze Estuary. Chemosphere 2017, 183, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.M.; Zhang, S.H.; Zhang, X.L.; Qu, J.M. Application of movable and portable shelter in farming of sea cucumber. Shandong Fish. 2004, 21, 9–10. (In Chinese) [Google Scholar]
- Ut, V.N.; Le, V.L.; Nghia, T.T.; Hong, T.T.T. Development of nursery culture techniques for the mud crab Scylla paramamosain (Estampador). Aquac. Res. 2007, 38, 1563–1568. [Google Scholar] [CrossRef]
- Benhaïm, D.; Leblanc, C.A.; Lucas, G. Impact of a new artificial shelter on Arctic charr (Salvelinus alpinus, L.) behaviour and culture performance during the endogenous feeding period. Aquaculture 2009, 295, 38–43. [Google Scholar] [CrossRef]
- He, J.; Gao, Y.; Xu, W.; Yu, F.; Su, Z.; Xuan, F. Effects of different shelters on the molting, growth and culture performance of Portunus trituberculatus. Aquaculture 2017, 481, 133–139. [Google Scholar] [CrossRef]
- Eileen, P.V.; Wolcot, T.G.; Wolcott, D.L. Nursery role of seagrass beds: Enhanced growth of juvenile blue crabs (Callinectes sapidus Rathbun). J. Exp. Mar. Biol. Ecol. 1996, 198, 155–173. [Google Scholar]
- Chen, X.L.; Zhang, X.L.; Li, S.J.; Wang, G.Z.; Lin, Q.W. Shelter preference of megalopae and first juvenile of mud crab, Scylla paramamosain (Estampador, 1949). J. Xiamen Univ. (Nat. Sci.) 2009, 48, 594–599. [Google Scholar]
- Geng, Z.; Feng, G.; Zhao, F.; Zhang, T.; Yang, G.; Zhang, T.; Zhao, Y. Distribution of larval crabs (Eriocheir sinensis) and relationship with oceanographic features in the Yangtze Estuary. Ecoscience 2018, 25, 125–134. [Google Scholar] [CrossRef]
- Liu, R.Q. Preliminary report on physico-chemical properties of main channel and tributaries in upper and middle reaches of the Changjiang river, before and after damming of the three gorges project. Acta Hydrobiol. Sin. 2000, 24, 446–450. [Google Scholar]
- Polis, G.A. The evolution and dynamics of intraspecific predation. Annu. Rev. Ecol. Syst. 1981, 12, 225–251. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, F.; Song, C.; Gao, Y.; Chai, Y.; Zhong, L.; Zhuang, P. Hatchery technology restores the spawning ground of phytophilic fish in the urban river of Yangtze Estuary, China. Urban Ecosyst. 2020, 23, 1087–1098. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, F.; Song, C.; Chai, Y.; Wang, Q.; Zhuang, P. Larva fish assemblage structure in three-dimensional floating wetlands and non-floating wetlands in the Changjiang River estuary. J. Oceanol. Limnol. 2021, 39, 721–731. [Google Scholar] [CrossRef]
- Song, T.; Ding, F.; Chen, J.; Wu, S.; Wang, X. Effects of broodstock sizes, shelter, illumination and stocking density on breeding in red swamp crayfish Procambarus clarkii. Fish. Sci. 2012, 31, 549–553, (In Chinese with English abstract). [Google Scholar]
- Karnofsky, E.B.; Atema, J.; Elgin, R.H. Field observations of social behavior, shelter use, and foraging in the lobster, Homarus americanus. Biol. Bull. 2013, 176, 239. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.Y. A Preliminary Study on the Comparison of the Efficiency of Crayfish and Crab Culture in Jianghan Area. Master’s Thesis, Yangtze University, Jingzhou, China, April 2017. [Google Scholar]
- Huang, X.; Xu, Q.; Feng, G.; Huang, X.; Geng, Z.; Zhao, F.; Zhuang, P. Circadian rhythm oscillation affects the structural morphology of eyes in juvenile Eriocheir sinensis H. Milne-Edwards, 1853 (Decapoda: Varunidae) exposed to alternating dark/light conditions. Zoomorphol. Res. Artic. 2020, 72, 245–253. [Google Scholar]
- David, L.M.; Tom, A.; Beverley, K.; Trent, L.; Brian, P. Stocking density and artificial habitat influence stock structure and yield from intensive nursery systems for mud crabs Scylla serrara. Aquac. Res. 2007, 38, 1580–1587. [Google Scholar]
- Zeng, G.C.; Jin, F.L. Model test of artificial shelter in loach breeding pond. Inland Fish. 1992, 93, 14–16. (In Chinese) [Google Scholar]
- Maximino, C.; Brito, T.; Moraes, F.; Oliveira, F.; Taccolini, I. A comparative analysis of the preference for dark environments in five teleosts. Int. J. Comp. Psychol. 2007, 20, 351–367. [Google Scholar]
- Maximino, C.; Brito, T.M.D.; Colmanetti, R.; Pontes, A.A.; Castro, H.M.D.; Lacerda, R.I.T.D.; Jr, A.G. Parametric analyses of anxiety in zebra fish scototaxis. Behav. Brain Res. 2010, 210, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rudnick, D.; Resh, V. Stable isotopes, mesocosms and gut content analysis demonstrate trophic differences in two invasive decapod crustacea. Freshw. Biol. 2010, 50, 1323–1336. [Google Scholar] [CrossRef]
- Menge, B.A.; Sutherland, J.P. Community regulation: Variation in disturbance, competition, and predation in relation to environmental stress and recruitment. Am. Nat. 1987, 130, 730–757. [Google Scholar] [CrossRef]
- Smith, K.N.; Herrnkind, W.F. Predation on early juvenile spiny lobsters Panulirus argus (Latreille): Influence of size and shelter. J. Exp. Mar. Biol. Ecol. 1992, 157, 3–18. [Google Scholar] [CrossRef]
- Huang, S.; Wang, J.; Yue, W.; Chen, J.; Gaughan, S.; Lu, W.; Lu, G.; Wang, C. Transcriptomic variation of hepatopancreas reveals the energy metabolism and biological processes associated with molting in Chinese mitten crab, Eriocheir sinensis. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, T.; Saito, M.; Yamashiro, T.; Hamano, T.; Nakata, K. Shelter preference for the Japanese mitten crab Eriocheir japonica: A field experiment in a river, southern central Japan. Ecol. Civil Eng. 2016, 19, 1–11. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, Y.X.; Li, Y.S. Crab breeding and crab culture-the biology of Eriocheir sinensis. Fish. Sci. Technol. Inf. 2007, 34, 25–28, (In Chinese with English abstract). [Google Scholar]
- Cowan, J.; Brown, G. Foraging trade-offs and predator inspection in an ostariophysan fish: Switching from chemical to visual cues. Behaviour 2000, 137, 181–195. [Google Scholar] [CrossRef]
- Figueiredo, B.R.S.; Mormul, R.P.; Thomaz, S.M. Swimming and hiding regardless of the habitat: Prey fish do not choose between a native and a non-native macrophyte species as a refuge. Hydrobiologia 2015, 746, 285–290. [Google Scholar] [CrossRef]
- Jia, E.T.; Yan, M.J.; Lai, Q.C.; Luo, W.T.; Jiang, G.Z. Feeding rhythm of the Chinese mitten crab (Eriocheir sinensis). J. Fish. Sci. China. 2018, 25, 546–554. (In Chinese) [Google Scholar] [CrossRef]
- Wang, K.X. Shrimp and Crab Breeding; China Agricultural Press: Beijing, China, 1997; pp. 32–36. (In Chinese) [Google Scholar]
- Dai, A.Y.; Yang, S.L.; Song, Y.Z.; Chen, G.X. Marine Crab of China; Marine Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Chen, X.L.; Lin, Q.W.; Li, S.J.; Wang, G.Z.; Ai, Q.X.; Li, B.L.; Lin, T. Observations and studies on the cannibalism among post larvae of Kuruma prawn, Penaeus japonicus. J. Xiamen Univ. (Nat. Sci.) 2003, 42, 358–361. [Google Scholar]
- Chen, Z.Q.; Chen, C.S.; Wu, Z.Q.; Lin, C.C.; Jia, X.W. The roosting habit of the spiny lobster Panulirus stimpsoni. Mar. Sci. 2001, 25, 11–15. (In Chinese) [Google Scholar]
- Reymond, H.; Lagardère, J.P. Feeding rhythms and food of Penaeus japonicus Bate (Crustacea, Penaeidae) in salt marsh ponds; role of halophilic entomofauna. Aquaculture 1990, 84, 125–143. [Google Scholar] [CrossRef]
- Zhao, X.; Xiong, L.H. Plant-climbing behavior of a mudflat crab, Chiromantes neglectum in 24 hour duration at Phragmites-dominated tidal marshin Changjiang River Estuary. Chin. J. Zool. 2016, 51, 423–433. (In Chinese) [Google Scholar]
- Xi, J.; Zheng, Z.L.; Mou, Z.B.; Liu, F.; Liu, X.Y.; Shen, J.; Liu, H.P.; Zhou, Y. Study on the hiding behavior of the Glyptosternum maculatum larvae and juveniles. Acta Hydrobiol. Sin. 2021, 45, 1129–1137. (In Chinese) [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Shen, C.; Feng, G.; Huang, X.; Li, X. Preference for Shelters at Different Developmental Stages of Chinese Mitten Crab (Eriocheir sinensis). Animals 2022, 12, 918. https://doi.org/10.3390/ani12070918
Li C, Shen C, Feng G, Huang X, Li X. Preference for Shelters at Different Developmental Stages of Chinese Mitten Crab (Eriocheir sinensis). Animals. 2022; 12(7):918. https://doi.org/10.3390/ani12070918
Chicago/Turabian StyleLi, Chunbo, Chenchen Shen, Guangpeng Feng, Xiaorong Huang, and Xincang Li. 2022. "Preference for Shelters at Different Developmental Stages of Chinese Mitten Crab (Eriocheir sinensis)" Animals 12, no. 7: 918. https://doi.org/10.3390/ani12070918