
Fermentation Quality and Microbial Community of Corn Stover or Rice Straw Silage Mixed with Soybean Curd Residue

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Materials and Silage Preparation
2.2. Analysis of Microbial Population, Nutritional Composition, and Fermentation Quality
2.3. Microbial Diversity Analysis
2.4. Statistical Analyses
3. Results
3.1. Characteristics of Raw Materials before Ensiling
3.2. Nutritional Composition of CS or RS Silage Mixed with SCR
3.3. Fermentation Quality of CS or RS Silage Mixed with SCR
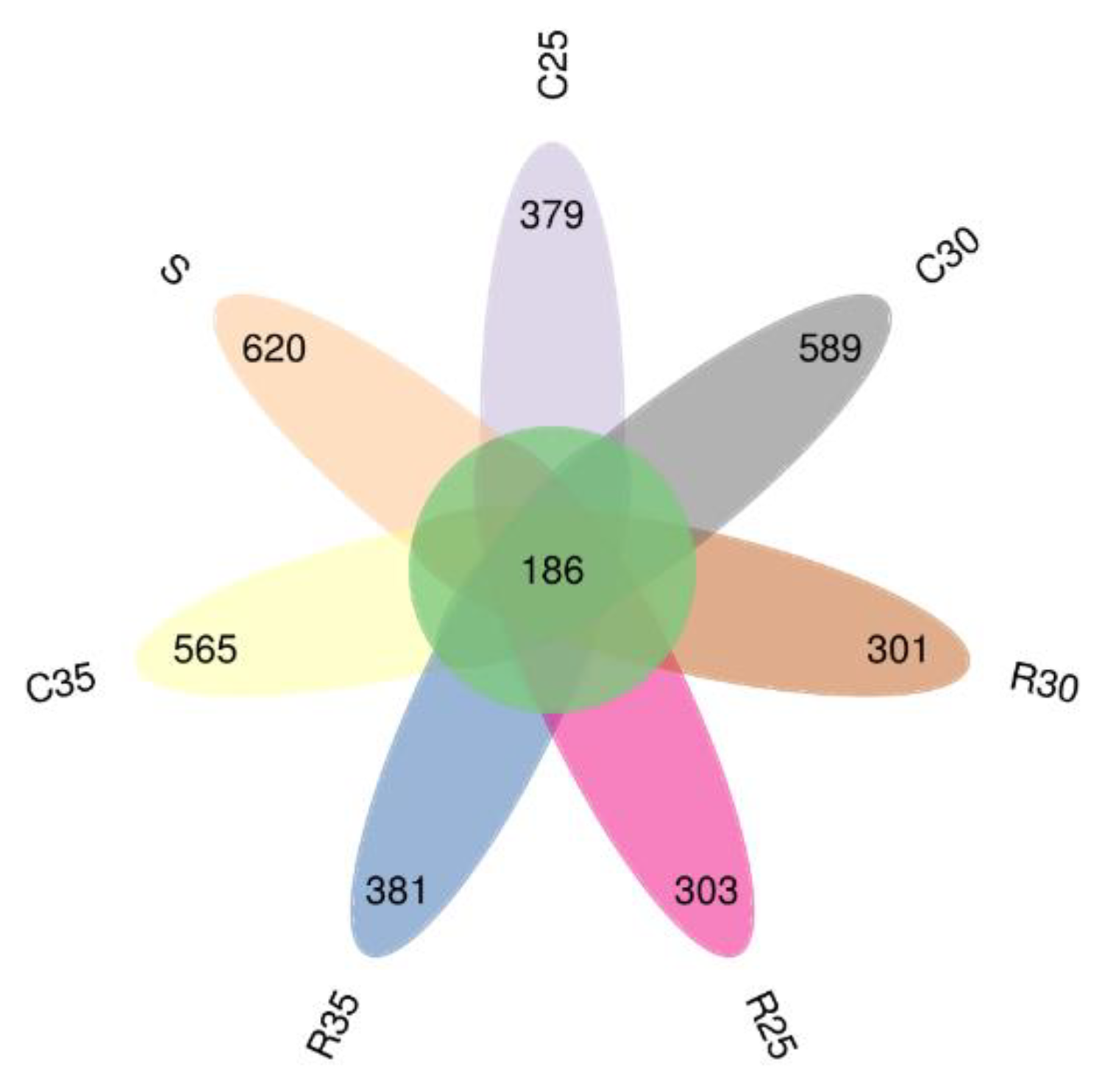
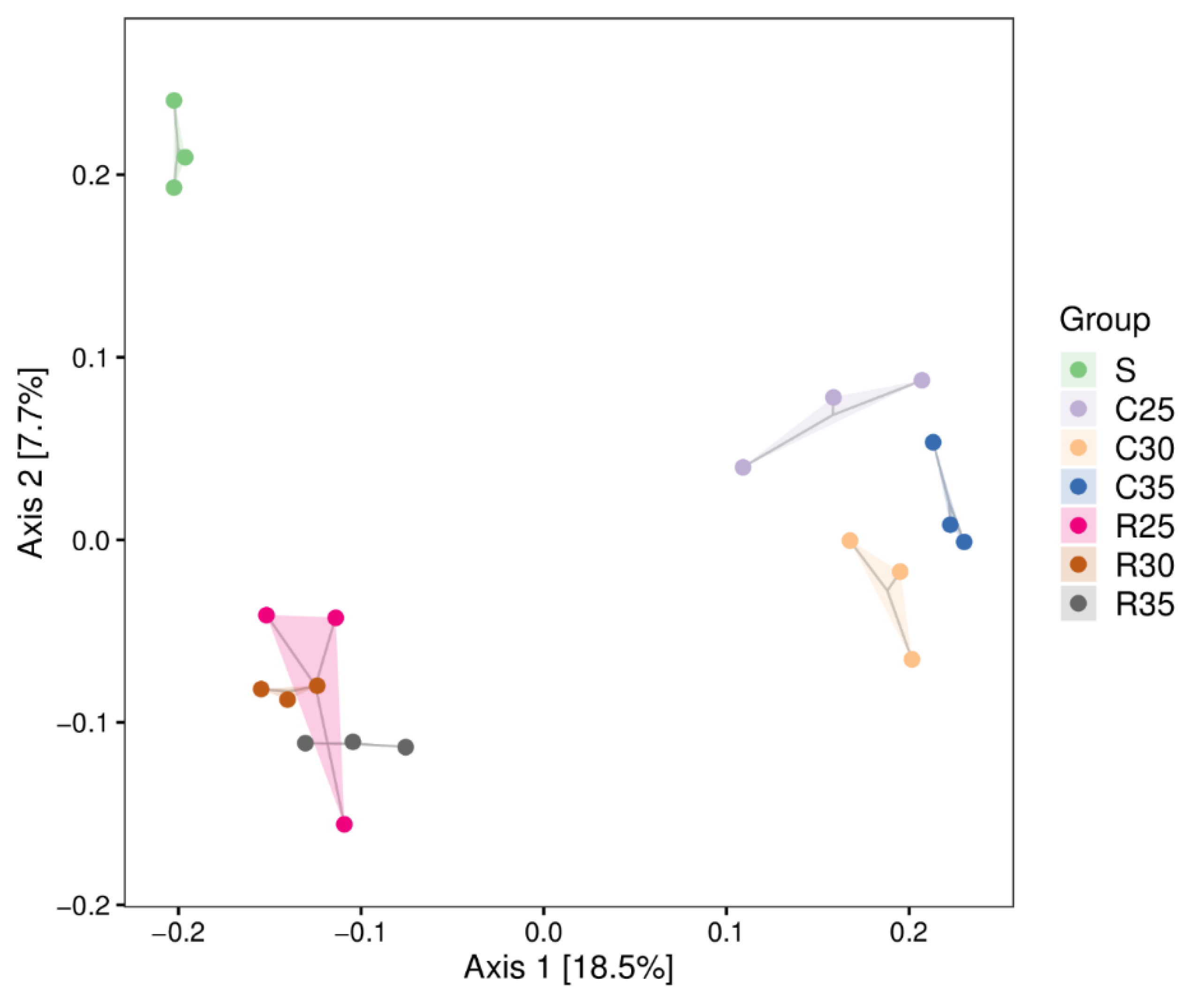
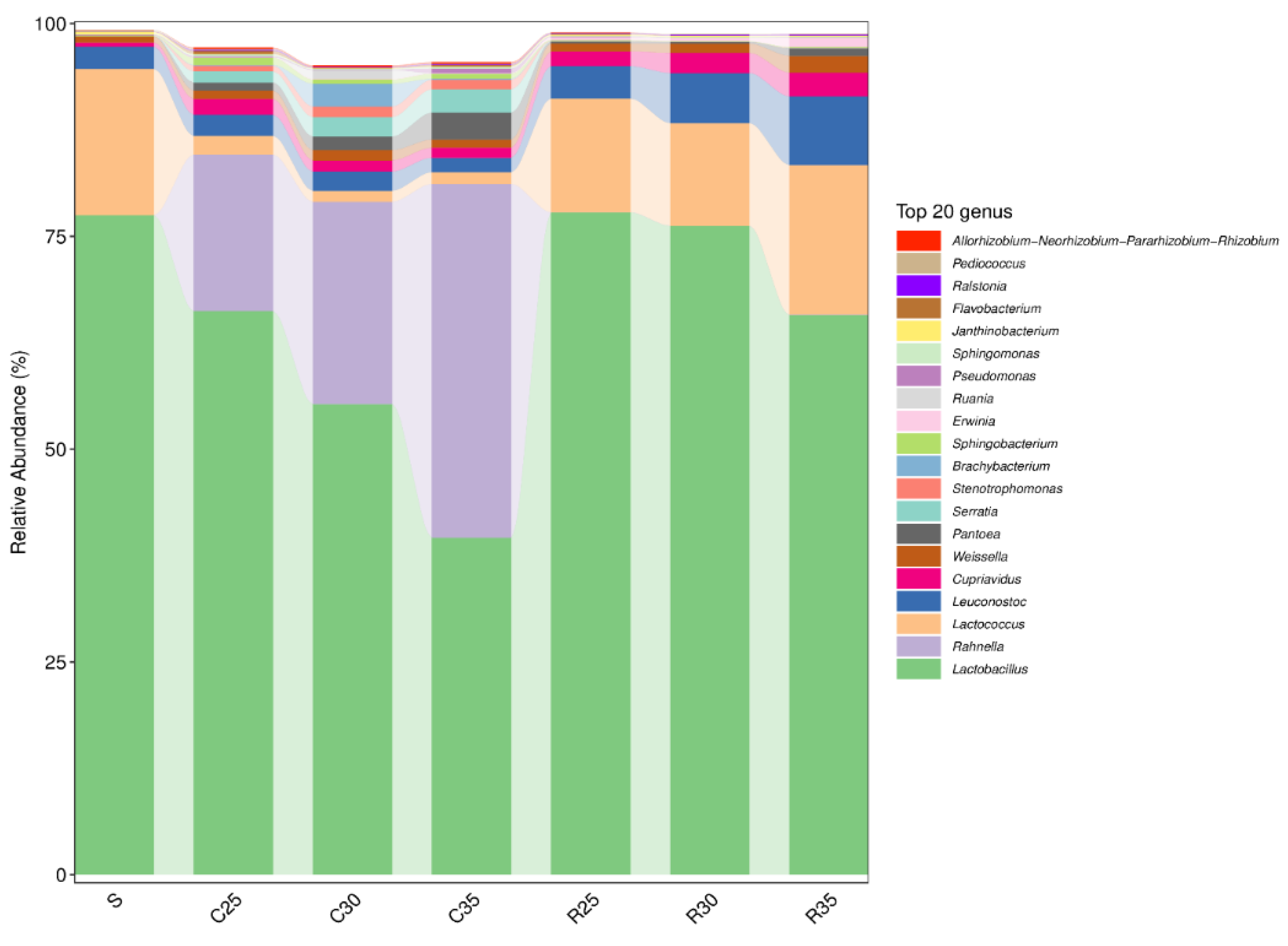
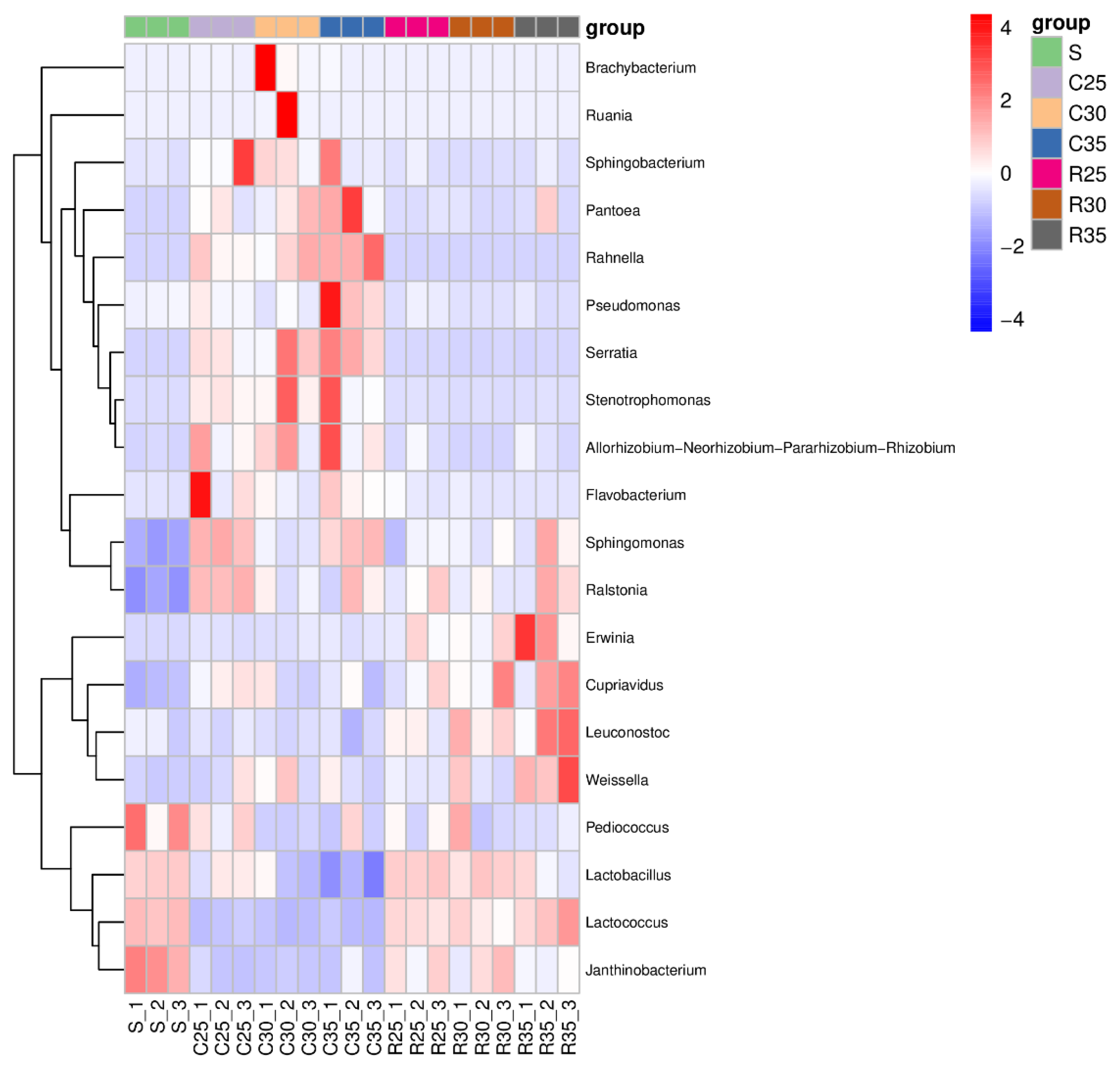
3.4. Microbial Community of CS or RS Silage Mixed with SCR
4. Discussion
4.1. Characteristics of Raw Materials before Ensiling
4.2. Nutritional Composition of CS or RS Silage Mixed with SCR
4.3. Fermentation Quality of CS or RS Silage Mixed with SCR
4.4. Microbial Community of CS or RS Silage Mixed with SCR
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, B.; Qiao, M.; Lu, F. Composition, Nutrition, and Utilization of Okara (Soybean Residue). Food Rev. Int. 2012, 28, 231–252. [Google Scholar] [CrossRef]
- Alvarez, M.V.; Cabred, S.; Ramirez, C.L.; Fanovich, M.A. Valorization of an agroindustrial soybean residue by supercritical fluid extraction of phytochemical compounds. J. Supercrit. Fluids 2019, 143, 90–96. [Google Scholar] [CrossRef]
- Borreani, G.; Tabacco, E.; Schmidt, R.J.; Holmes, B.J.; Muck, R.E. Silage review: Factors affecting dry matter and quality losses in silages. J. Dairy Sci. 2018, 101, 3952–3979. [Google Scholar] [CrossRef] [Green Version]
- Yang, C. Soybean milk residue ensiled with peanut hulls: Fermentation acids, cell wall composition, and silage utilization by mixed ruminal microorganisms. Bioresour. Technol. 2005, 96, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Scar, J.; Sánchez, A.B. Trends in biotechnological production of fuel ethanol from different feedstocks—ScienceDirect. Bioresour. Technol. 2008, 99, 5270–5295. [Google Scholar]
- Yan, C.; Yan, S.S.; Jia, T.Y.; Dong, S.K.; Gong, Z.P. Decomposition characteristics of rice straw returned to the soil in northeast China. Nutr. Cycl. Agroecosys. 2019, 114, 211–224. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, W.; Wang, C.; Yang, F.Y.; Chen, X.Y.; Zhang, Q. Effect on the ensilage performance and microbial community of adding Neolamarckia cadamba leaves to corn stalks. Microb. Biotechnol. 2020, 13, 1502–1514. [Google Scholar] [CrossRef]
- Aggarwal, N.K.; Goyal, V.; Saini, A.; Yadav, A.; Gupta, R. Enzymatic saccharification of pretreated rice straw by cellulases from Aspergillus niger BK01. Biotech 2017, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.U.; Hjort-Gregersen, K.; Vazifehkhoran, A.H.; Jin, M.T. Co-ensiling of straw with sugar beet leaves increases the methane yield from straw. Bioresour. Technol. 2017, 245, 106–115. [Google Scholar] [CrossRef]
- Ni, K.; Zhao, J.; Zhu, B.; Su, R.; Pan, Y.; Ma, J.; Zhou, G.; Yong, T.; Liu, X.; Zhong, J. Assessing the fermentation quality and microbial community of the mixed silage of forage soybean with crop corn or sorghum. Bioresour. Technol. 2018, 265, 563–567. [Google Scholar] [CrossRef]
- Rauha, J.P.; Remes, S.; Heinonen, M.; Hopia, A.; Kähkönen, M.; Kujala, T.; Pihlaja, K.; Vuorela, H.; Vuorela, P. Antimicrobial effects of Finnish plant extracts containing flavonoids and other phenolic compounds. Int. J. Food Microbiol. 2000, 56, 3–12. [Google Scholar] [CrossRef]
- AOAC, Official Methods of Analysis: 15th Edition. Trac Trends Anal. Chem. 1990, 9, VI.
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Mcdonald, P.; Henderson, A.R. Determination of water-soluble carbohydrates in grass. J. Sci. Food Agric. 2010, 15, 395–398. [Google Scholar] [CrossRef]
- Yuan, X.; Guo, G.; Wen, A.; Desta, S.T.; Wang, J.; Wang, Y.; Shao, T. The effect of different additives on the fermentation quality, in vitro digestibility and aerobic stability of a total mixed ration silage. Anim. Feed Sci. Technol. 2015, 207, 41–50. [Google Scholar] [CrossRef]
- Novamsky, I.; VanEck, R.; Schouwenburg, C.V.; Walinga, I. Total nitrogen determination in plant material by means of the indophenol-blue method. Neth. J. Agric. Sci. 1974, 22, 3–5. [Google Scholar] [CrossRef]
- Cai, Y.M.; Benno, Y.; Ogawa, M.; Ohmomo, S.; Nakase, T. Influence of lactobacillus spp. from An inoculant and of weissella and leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 1998, 64, 2982–2987. [Google Scholar] [CrossRef] [Green Version]
- Yitbarek, M.B.; Tamir, B. Silage Additives: Review. Open J. Appl. Sci. 2014, 4, 258–274. [Google Scholar] [CrossRef] [Green Version]
- Lowilai, P.; Kabata, K.; Okamoto, C. Effects of rice bran and wheat bran on fermentation quality and chemical composition of water hyacinth silage. Jpn. J. Grassl. Sci. 1994, 40, 271–277. [Google Scholar]
- Bao, W.; Mi, Z.; Xu, H.; Zheng, Y.; Kwok, L.Y.; Zhang, H.; Zhang, W. Assessing quality of Medicago sativa silage by monitoring bacterial composition with single molecule, real-time sequencing technology and various physiological parameters. Sci. Rep. 2016, 6, 28358. [Google Scholar] [CrossRef]
- Cai, Y.M.; Kumai, S.; Ogawa, M.; Benno, Y.; Nakase, T. Characterization and Identification of Pediococcus Species Isolated from Forage Crops and Their Application for Silage Preparation. Appl. Environ. Microbiol. 1999, 65, 2901–2906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.; Zanetti, D.; Rotta, P.; Avezedo, H. Energy and protein nutritional requirements for Nellore bulls. Rev. Bras. De Zootec. 2013, 41, 1516–1524. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.M.; Mat, K.; Ishigaki, G.; Akashi, R. A review of okara (soybean curd residue) utilization as animal feed: Nutritive value and animal performance aspects. Anim. Sci. J. 2021, 92, e13594. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zi, X.; Zhou, H.; Lv, R.; Tang, J.; Cai, Y. Silage fermentation and ruminal degradation of cassava foliage prepared with microbial additive. AMB Express 2019, 9, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Villares, A.; Rostagno, M.A.; García-Lafuente, A.; Guillamón, E.; Martínez, J.A. Content and Profile of Isoflavones in Soy-Based Foods as a Function of the Production Process. Food Bioprocess Technol. 2011, 4, 27–38. [Google Scholar] [CrossRef]
- Ungar, Y.; Osundahunsi, O.F.; Shimoni, E. Thermal Stability of Genistein and Daidzein and Its Effect on Their Antioxidant Activity. J. Agric. Food Chem. 2003, 51, 4394–4399. [Google Scholar] [CrossRef]
- Ulanowska, K.; Tkaczyk, A.; Konopa, G.; Wgrzyn, G. Differential antibacterial activity of genistein arising from global inhibition of DNA, RNA and protein synthesis in some bacterial strains. Arch. Microbiol. 2006, 184, 271–278. [Google Scholar] [CrossRef]
- Hong, H.; Landauer, M.R.; Foriska, M.A.; Ledney, G.D. Antibacterial activity of the soy isoflavone genistein. J. Basic Microbiol. 2006, 4, 329–335. [Google Scholar] [CrossRef]
- Li, R.; Jiang, D.; Zheng, M.; Tian, P.; Xu, C. Microbial community dynamics during alfalfa silage with or without clostridial fermentation. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Mu, L.; Xie, Z.; Hu, L.X.; Chen, G.H.; Zhang, Z.F. Cellulase interacts with Lactobacillus plantarum to affect chemical composition, bacterial communities, and aerobic stability in mixed silage of high-moisture amaranth and rice straw. Bioresour. Technol. 2020, 315, 123772. [Google Scholar] [CrossRef]
- Oude Elferink, S.J.; Krooneman, J.; Gottschal, J.C.; Spoelstra, S.F.; Faber, F.; Driehuis, F. Anaerobic Conversion of Lactic Acid to Acetic Acid and 1,2-Propanediol by Lactobacillus buchneri. Appl. Environ. Microbiol. 2001, 67, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogunade, I.M.; Jiang, Y.; Cervantes, A.A.; Kim, D.H.; Oliveira, A.S.; Vyas, D.; Weinberg, Z.G.; Jeong, K.C.; Adesogan, A.T. Bacterial diversity and composition of alfalfa silage as analyzed by Illumina MiSeq sequencing: Effects of Escherichia coli O157:H7 and silage additives. J. Dairy Sci. 2017, 101, 2048–2059. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Wang, C.; Xing, Y.; Zhou, W.; Zhang, Q. Ensiling characteristics, proteolysis and bacterial community of high-moisture corn stalk and stylo silage prepared with Bauhinia variegate flower. Bioresour. Technol. 2019, 296, 122336. [Google Scholar] [CrossRef] [PubMed]
- Penrose, D.M.; Glick, B.R. Methods for isolating and characterizing ACC deaminase-containing plant growth-promoting rhizobacteria. Physiol. Plant. 2010, 118, 10–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vyas, P.; Joshi, R.; Sharma, K.C.; Rahi, P.; Gulati, A. Cold-Adapted and Rhizosphere-Competent Strain of Rahnella sp. with Broad-Spectrum Plant Growth-Promotion Potential. J. Microbiol. Biotechnol. 2010, 20, 1724. [Google Scholar]
- Kung, L.; Shaver, R. Interpretation and use of silage fermentation analysis reports. Focus Forage 2001, 3, 1–5. [Google Scholar]
- Dunière, L.; Sindou, J.; Chaucheyras-Durand, F.; Chevallier, I.; Thévenot-Sergentet, D. Silage processing and strategies to prevent persistence of undesirable microorganisms. Anim. Feed Sci. Technol. 2013, 182, 1–15. [Google Scholar] [CrossRef]
- Jimtha, C.J.; Jishma, P.; Sreelekha, S.; Chithra, S.; Radhakrishnan, E. Antifungal properties of prodigiosin producing rhizospheric Serratia sp. Rhizosphere 2017, 3, 105–108. [Google Scholar] [CrossRef]
- Sasidharan, A.; Sasidharan, N.K.; Vasu, R.K.; Nataraja, A.V.; Bhaskaran, K. Antifungal activity of violacein purified from a novel strain of Chromobacterium sp. NIIST (MTCC 5522). J. Microbiol. 2015, 53, 694–701. [Google Scholar] [CrossRef]
- Anju, S.; Kumar, N.S.; Krishnakumar, B.; Kumar, B.S. Synergistic combination of violacein and azoles that leads to enhanced killing of major human pathogenic dermatophytic fungi Trichophyton rubrum. Front. Cell. Infect. Microbiol. 2015, 5, 57. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items 1 | Soybean Curd Residue | Corn Stover | Rice Straw |
|---|---|---|---|
| DM, g/kg FM | 166.2 ± 3.21 | 922.3 ± 8.45 | 953.9 ± 5.82 |
| CP, g/kg DM | 129.3 ± 0.31 | 36.7 ± 0.65 | 34.0 ± 0.94 |
| NDF, g/kg DM | 552.9 ± 8.01 | 659.0 ± 5.49 | 652.8 ± 5.77 |
| ADF, g/kg DM | 300.5 ± 4.26 | 399.1 ± 5.60 | 393.0 ± 3.67 |
| WSC, g/kg DM | 22.9 ± 0.13 | 123.8 ± 5.24 | 146.5 ± 4.65 |
| Items 1 | C35 | C30 | C25 | R35 | R30 | R25 |
|---|---|---|---|---|---|---|
| DM, g/kg FM | 431.2 ± 6.15 | 385.7 ± 3.76 | 349.1 ± 3.74 | 437.1 ± 3.05 | 402.9 ± 1.39 | 361.5 ± 4.42 |
| CP, g/kg DM | 96.2 ± 1.24 | 101.5 ± 0.38 | 107.1 ± 0.35 | 95.4 ± 0.63 | 101.3 ± 0.86 | 106.5 ± 0.55 |
| NDF, g/kg DM | 595.4 ± 1.55 | 584.8 ± 1.16 | 578.3 ± 6.77 | 586.7 ± 2.94 | 580.2 ± 3.06 | 574.5 ± 1.49 |
| ADF, g/kg DM | 337.2 ± 3.72 | 330.2 ± 1.54 | 322.3 ± 1.16 | 334.9 ± 2.88 | 326.8 ± 3.89 | 320.4 ± 2.31 |
| WSC, g/kg DM | 58.1 ± 0.51 | 53.3 ± 1.21 | 50.8 ± 0.37 | 67.7 ± 1.59 | 59.0 ± 1.12 | 53.7 ± 0.79 |
| Items 1 | Corn Stover | Rice Straw | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C35 | C30 | C25 | SEM | p Value | R35 | R30 | R25 | SEM | p Value | |
| DM (g/kg FM) | 421.0 a | 380.1 b | 347.9 c | 1.87 | <0.0001 | 430.1 a | 394.9 b | 354.2 c | 2.26 | <0.0001 |
| CP (g/kg DM) | 34.0 c | 55.6 b | 78.9 a | 1.27 | <0.0001 | 29.6 c | 52.0 b | 75.4 a | 0.73 | <0.0001 |
| NDF (g/kg DM) | 606.3 a | 581.4 b | 568.3 c | 4.36 | 0.0023 | 629.7 a | 609.3 b | 586.0 c | 4.78 | 0.0020 |
| ADF (g/kg DM) | 377.0 a | 357.6 b | 342.3 c | 2.27 | 0.0001 | 389.3 a | 370.7 b | 363.6 c | 3.01 | 0.0026 |
| WSC (g/kg DM) | 36.5 a | 32.3 b | 28.6 c | 0.77 | 0.0011 | 39.2 a | 34.7 ab | 31.8 c | 1.69 | 0.0560 |
| Items 1 | Corn Stover | Rice Straw | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| C35 | C30 | C25 | SEM | p Value | R35 | R30 | R25 | SEM | p Value | |
| pH | 4.01 a | 3.93 b | 3.86 b | 0.02 | 0.0030 | 3.96 a | 3.88 b | 3.85 c | 0.02 | 0.0019 |
| AN (g/kg TN) | 50.8 a | 47.8 b | 45.1 c | 0.59 | 0.0016 | 47.5 a | 43.6 b | 39.9 c | 0.67 | 0.0007 |
| LA (g/kg DM) | 15.8 c | 18.2 b | 21.5 a | 0.51 | 0.0007 | 11.5 c | 14.1 b | 16.4 a | 0.50 | 0.0010 |
| AA (g/kg DM) | 21.4 b | 27.3 a | 27.1 a | 0.58 | 0.0006 | 30.2 b | 43.3 a | 42.5 a | 0.63 | <0.0001 |
| PA (g/kg DM) | 5.37 | 2.61 | ND | - | - | 4.17 | ND | ND | - | - |
| BA (g/kg DM) | ND | ND | ND | - | - | ND | ND | ND | - | - |
| LAB (log10 CFU/g FM) | 7.11 c | 7.87 b | 8.25 a | 0.04 | <0.0001 | 7.33 b | 7.85 a | 7.96 a | 0.06 | 0.0004 |
| Yeast (log10 CFU/g FM) | 2.82 | 2.48 | <2.00 | - | - | 2.84 | 2.01 | <2.00 | - | - |
| Mold (log10 CFU/g FM) | <2.00 | <2.00 | <2.00 | - | - | <2.00 | <2.00 | <2.00 | - | - |
| Item | Corn Stover | Rice Straw | ||||
|---|---|---|---|---|---|---|
| C35 | C30 | C25 | R35 | R30 | R25 | |
| Chao1 | 833.64 | 901.73 | 775.93 | 657.68 | 707.66 | 710.39 |
| Shannon | 5.91 | 5.80 | 5.77 | 5.59 | 5.52 | 5.48 |
| Simpson | 0.918 | 0.948 | 0.954 | 0.936 | 0.943 | 0.949 |
| Coverage | 0.997 | 0.996 | 0.997 | 0.997 | 0.997 | 0.997 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Song, J.; Liu, Z.; Zhang, G.; Zhang, Y. Fermentation Quality and Microbial Community of Corn Stover or Rice Straw Silage Mixed with Soybean Curd Residue. Animals 2022, 12, 919. https://doi.org/10.3390/ani12070919
Wang X, Song J, Liu Z, Zhang G, Zhang Y. Fermentation Quality and Microbial Community of Corn Stover or Rice Straw Silage Mixed with Soybean Curd Residue. Animals. 2022; 12(7):919. https://doi.org/10.3390/ani12070919
Chicago/Turabian StyleWang, Xiaolin, Jiamei Song, Zihan Liu, Guangning Zhang, and Yonggen Zhang. 2022. "Fermentation Quality and Microbial Community of Corn Stover or Rice Straw Silage Mixed with Soybean Curd Residue" Animals 12, no. 7: 919. https://doi.org/10.3390/ani12070919