Dead Brood of Apis mellifera Is Removed More Effectively from Small-Cell Combs Than from Standard-Cell Combs


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Characteristics of Bee Colonies
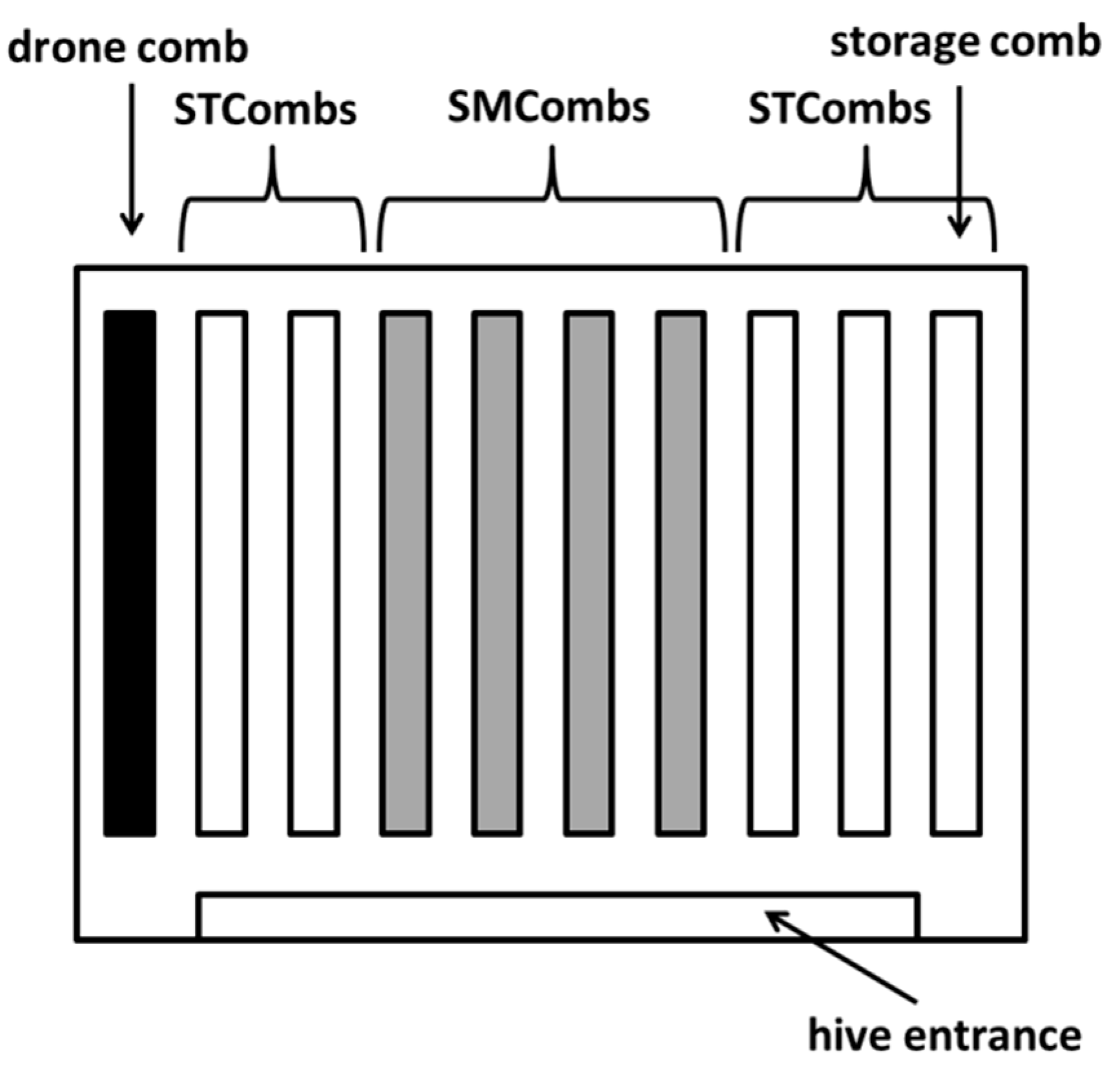
2.2. Assessment of Hygienic Behaviour
2.3. Measurements of Comb Cell Width
2.4. Statistical Analysis
3. Results
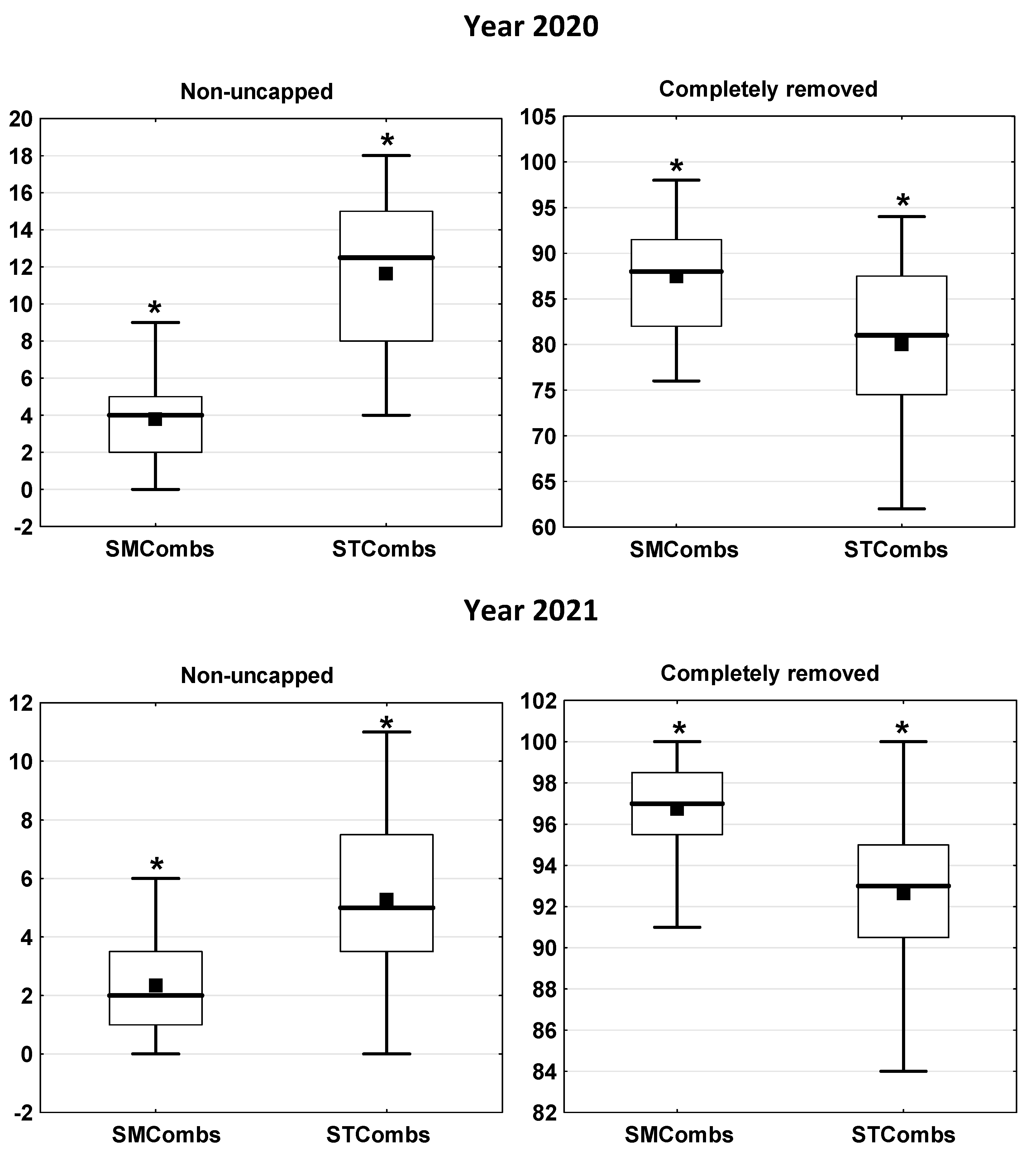
3.1. The Efficiency of Hygienic Behaviour
3.2. Comb Cell Width
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siva-Jothy, M.T.; Moret, Y.; Rolff, J. Evolutionary ecology of insect immunity. Adv. Insect. Physiol. 2005, 32, 1–48. [Google Scholar] [CrossRef]
- Cremer, S.; Armitage, S.A.; Schmid-Hempel, P. Social immunity. Curr. Biol. 2007, 17, 693–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.D.; Spivak, M. Socialized medicine: Individual and communal disease barriers in honey bees. J. Invertebr. Pathol. 2010, 103, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Davis, H.E.; Meconcelli, S.; Radek, R.; McMahon, D.P. Termites shape their collective behavioural response based on stage of infection. Sci. Rep. 2018, 8, 14433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pull, C.D.; Ugelvig, L.V.; Wiesenhofer, F.; Grasse, A.V.; Tragust, S.; Schmitt, T.; Brown, M.J.F.; Cremer, S. Destructive disinfection of infected brood prevents systemic disease spread in ant colonies. Elife 2018, 7, e32073. [Google Scholar] [CrossRef] [PubMed]
- Currie, C.R.; Stuart, A.E. Weeding and grooming of pathogens in agriculture by ants. Proc. R. Soc. B 2001, 268, 1033–1039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Toufailia, H.; Alves, D.A.; Bento, J.M.S.; Marchini, L.C.; Ratnieks, F.L.W. Hygienic behaviour in Brazilian stingless bees. Biol. Open 2016, 11, 1712–1718. [Google Scholar] [CrossRef] [Green Version]
- Michener, C.D. The Social Behavior of the Bees; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Spivak, M.; Danka, R.G. Perspectives on hygienic behavior in Apis mellifera and other social insects. Apidologie 2021, 52, 1–16. [Google Scholar] [CrossRef]
- Rothenbuhler, W.C. Behaviour genetics of nest cleaning in honey bees. I. Responses of four inbred lines to disease- killed brood. Anim. Behav. 1964, 12, 578–583. [Google Scholar] [CrossRef]
- Spivak, M. Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie 1996, 27, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Arathi, H.S.; Burns, I.; Spivak, M. Ethology of hygienic behaviour in the honey bee Apis mellifera L. (hymenoptera: Apidae) behavioural repertoire of hygienic bees. Ethology 2000, 106, 368–379. [Google Scholar] [CrossRef] [Green Version]
- Büchler, R.; Berg, S.; Le Conte, Y. Breeding for resistance to Varroa destructor in Europe. Apidologie 2010, 41, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M.; Reuter, G.S. Resistance to American foulbrood disease by honey bee colonies Apis mellifera bred for hygienic behavior. Apidologie 2001, 32, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Invernizzi, C.; Rivas, F.; Betticci, L. Resistance to Chalkbrood Disease in Apis mellifera L. (Hymenoptera: Apidae) Colonies with Different Hygienic Behaviour. Neotrop. Entomol. 2011, 40, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Palacio, M.A.; Figini, E.E.; Ruffinengo, S.R.; Rodriguez, E.M.; del Hoyo, M.L.; Bedascarrasbure, E.L. Changes in a population of Apis mellifera L. selected for hygienic behaviour and its relation to brood disease tolerance. Apidologie 2000, 31, 471–478. [Google Scholar] [CrossRef]
- Rinderer, T.E.; Harris, J.W.; Hunt, G.J.; de Guzman, L.I. Breeding for resistance to Varroa destructor in North America. Apidologie 2010, 41, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Milani, N. The resistance of Varroa jacobsoni Oud. to acaricides. Apidologie 1999, 30, 229–234. [Google Scholar] [CrossRef]
- Kochansky, J.; Knox, D.A.; Feldlaufer, M.; Pettis, J.S. Screening alternative antibiotics against oxytetracycline-susceptible and-resistant Paenibacillus larvae. Apidologie 2001, 32, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.D. Diverse origins of tetracycline resistance in the honey bee bacterial pathogen Paenibacillus larvae. J. Invertbr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Sammataro, D.; Untalan, P.; Guerrero, F.; Finley, J. The resistance of varroa mites (Acari: Varroidae) to acaricides and the presence of esterase. Int. J. Acarol. 2005, 31, 67–74. [Google Scholar] [CrossRef]
- Wallner, K. Varroacides and their residues in bee products. Apidologie 1999, 30, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Serra-Bonvehi, J.; Orantes-Bermejo, J. Acaricides and their residues in Spanish commercial beeswax. Pest Manag. Sci. 2010, 66, 1230–1235. [Google Scholar] [CrossRef] [PubMed]
- Gerdts, J.; Dewar, R.L.; Simone-Finstrom, M.; Edwards, T.; Angove, M. Hygienic behaviour selection via freeze-killed honey bee brood not associated with chalkbrood resistance in eastern Australia. PLoS ONE 2018, 13, e0203969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olszewski, K.; Borsuk, G.; Paleolog, J.; Strachecka, A.; Bajda, M. Hygienic behaviour of colonies kept on small-cell combs. Med. Weter. 2014, 70, 774–776. [Google Scholar]
- De la Rua, P.; Jaffe, R.; Dall’Olio, R.; Munoz, I.; Serrano, J. Biodiversity, conservation and current threats to European honeybees. Apidologie 2009, 40, 263–284. [Google Scholar] [CrossRef] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, 96–119. [Google Scholar] [CrossRef]
- Seeley, T.D.; Griffin, S.R. Small-cell comb does not control Varroa mites in colonies of honeybees of European origin. Apidologie 2011, 42, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Singer, H.J.; van Praagh, J.P.; Paulus, H.F. Interactions between honeybees and Varroa mites influenced by cell sizes and hygienic behaviour. Entomol. Gen. 2019, 38, 255–273. [Google Scholar] [CrossRef]
- Maggi, M.; Damiani, N.; Ruffinengo, S.; De Jong, D.; Principal, J.; Eguaras, M. Brood cell size of Apis mellifera modifies the reproductive behavior of Varroa destructor. Exp. Appl. Acarol. 2010, 50, 269–279. [Google Scholar] [CrossRef]
- Message, D.; Goncalves, L.S. Effect of the size of worker brood cells of Africanized honey bees on infestation and of the ectoparasitic mite Varroa jacobsoni Oud. Apidologie 1995, 26, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Piccirillo, G.A.; De Jong, D. The influence of brood comb cell size on the reproductive behavior of the ectoparasitic mite Varroa destructor in Africanized honey bee colonies. Genet. Mol. Res. 2003, 2, 36–42. [Google Scholar] [PubMed]
- Ellis, A.M.; Hayes, G.W.; Ellis, J.D. The efficacy of small cell foundation as a varroa mite (Varroa destructor) control. Exp. Appl. Acarol. 2009, 47, 311–316. [Google Scholar] [CrossRef]
- Berry, J.A.; Owens, W.B.; Delaplane, K.S. Small-cell comb foundation does not impede varroa mite population growth in honey bee colonies. Apidologie 2010, 41, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.A.; Goodwin, R.M.; McBrydie, H.M.; Cox, H.M. The effect of honey bee worker brood cell size on Varroa destructor infestation and reproduction. J. Apicult. Res. 2008, 47, 239–242. [Google Scholar] [CrossRef]
- Coffey, M.F.; Breen, J.; Brown, M.J.F.; McMullan, J.B. Brood-cell size has no influence on the population dynamics of Varroa destructor mites in the native western honey bee, Apis mellifera mellifera. Apidologie 2010, 41, 522–530. [Google Scholar] [CrossRef] [Green Version]
- Dziechciarz, P.; Borsuk, G.; Olszewski, K. Possibility to change the body size in worker bees by a combination of small-cell and standard-cell combs in the same nest. Apidologie 2021, 52, 1017–1032. [Google Scholar] [CrossRef]
- McMullan, J.B.; Brown, M.J.F. The influence of small-cell brood combs on the morphometry of honeybees (Apis mellifera). Apidologie 2006, 37, 665–672. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, K.; Borsuk, G.; Paleolog, J.; Strachecka, A. Life span of worker honeybees reared in colonies kept on small-cell combs. Med. Weter. 2014, 70, 777–780. [Google Scholar]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 2013, 52, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Olszewski, K.; Borsuk, G.; Paleolog, J.; Strachecka, A. Validation of the methods of hygienic behaviour evaluation in the honeybee. Med. Weter. 2013, 69, 749–751. [Google Scholar]
- McAfee, A.; Chapman, A.; Iovinella, I.; Gallagher-Kurtzke, Y.; Collins, T.F.; Higo, H.; Higo, H.; Madilao, L.L.; Pelosi, P.; Foster, L.J. A death pheromone, oleic acid, triggers hygienic behavior in honey bees (Apis mellifera L.). Sci. Rep. 2018, 8, 5719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak, M.; Masterman, R.; Ross, R.; Mesce, K.A. Hygienic behavior in the honey bee (Apis mellifera L.) and the modulatory role of octopamine. J. Neurobiol. 2003, 55, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Masterman, R.; Ross, R.; Mesce, K.; Spivak, M. Olfactory and behavioral response thresholds to odors of diseased brood differ between hygienic and non-hygienic honey bees (Apis mellifera L.). J. Comp. Physiol. A 2001, 187, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Gramacho, K.P.; Spivak, M. Differences in olfactory sensitivity and behavioral responses among honey bees bred for hygienic behavior. Behav. Ecol. Sociobiol. 2003, 54, 472–479. [Google Scholar] [CrossRef]
- Olszewski, K.; (University of Life Sciences in Lublin, Lublin, Poland). Elucidation of the phenomenon of behavioural overdominance of honeybee colonies kept on two types of combs with standard- and small-cell size. Personal communication, 2021, project unpublished outcomes. [Google Scholar]
- Garófalo, C.A. Bionomics of Bombus (fervidobombus) morio: 2. Body size and length of life of workers. J. Apic. Res. 1978, 17, 130–136. [Google Scholar] [CrossRef]
- Spaethe, J.; Weidenmüller, A. Size variation and foraging rate in bumblebees (Bombus terrestris). Insectes Sociaux 2002, 49, 142–146. [Google Scholar] [CrossRef] [Green Version]
- Worden, B.D.; Skemp, A.K.; Papaj, D.R. Learning in two contexts: The effects of interference and body size in bumblebees. J. Exp. Biol. 2005, 208, 2045–2053. [Google Scholar] [CrossRef] [Green Version]
- Strachecka, A.; Olszewski, K.; Kuszewska, K.; Paleolog, J.; Woyciechowski, M. Reproductive Potential Accelerates Preimaginal Development of Rebel Workers in Apis mellifera. Animals 2021, 11, 3245. [Google Scholar] [CrossRef]
- Strachecka, A.; Olszewski, K.; Kuszewska, K.; Chobotow, J.; Wójcik, Ł.; Paleolog, J.; Woyciechowski, M. Segmentation of the subcuticular fat body in Apis mellifera females with different reproductive potentials. Sci. Rep. 2021, 11, 13887. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dziechciarz, P.; Borsuk, G.; Olszewski, K. Dead Brood of Apis mellifera Is Removed More Effectively from Small-Cell Combs Than from Standard-Cell Combs. Animals 2022, 12, 418. https://doi.org/10.3390/ani12040418
Dziechciarz P, Borsuk G, Olszewski K. Dead Brood of Apis mellifera Is Removed More Effectively from Small-Cell Combs Than from Standard-Cell Combs. Animals. 2022; 12(4):418. https://doi.org/10.3390/ani12040418
Chicago/Turabian StyleDziechciarz, Piotr, Grzegorz Borsuk, and Krzysztof Olszewski. 2022. "Dead Brood of Apis mellifera Is Removed More Effectively from Small-Cell Combs Than from Standard-Cell Combs" Animals 12, no. 4: 418. https://doi.org/10.3390/ani12040418