
Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement


Abstract
:Simple Summary
Abstract
1. Introduction
2. Factors Affecting the Survival of Spermatozoa during Liquid Storage
2.1. Temperature
2.2. Concentration of Spermatozoa
2.3. Extenders
Key Extender Ingredients
3. Changes to Spermatozoa during Liquid Preservation
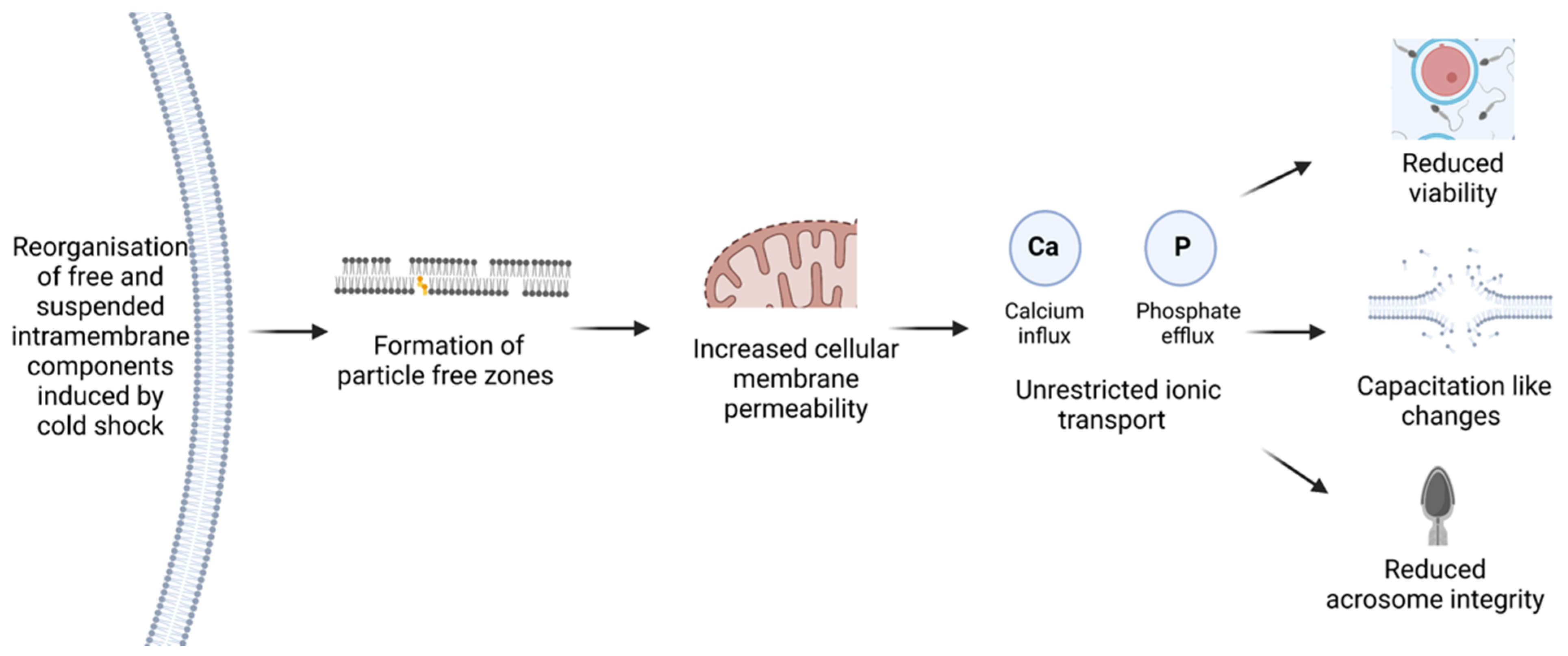
3.1. Membrane Redistribution
3.2. Disruption of Respiration and Subsequent Decrease in Motility
3.3. Lipid Peroxidation and Production of Reactive Oxygen Species
3.3.1. Lipid Peroxidation Biochemistry
3.3.2. Impact of Lipid Peroxidation on Sperm Function
3.3.3. Natural Defences to Reduce Lipid-Peroxidation-Induced Damage during Liquid Storage
4. The Use of Antioxidant Supplementation to Attenuate Lipid Peroxidation Stress and Promote Liquid Spermatozoa Survival during Storage at 23 °C
4.1. L-Carnitine
4.2. Pyruvate
4.3. Melatonin (N-Acetyl-5-Methoxytryptamine)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salamon, S.; Maxwell, W.M.C. Storage of Ram Semen. Anim. Reprod. Sci. 2000, 62, 77–111. [Google Scholar] [CrossRef]
- Brinsko, S.P. Insemination Doses: How Low Can We Go? Theriogenology 2006, 66, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Gillan, L.; Maxwell, W.M.C.; Evans, G. Preservation and Evaluation of Semen for Artificial Insemination. Reprod. Fertil. Dev. 2004, 16, 447–454. [Google Scholar] [CrossRef]
- Kameni, S.L.; Meutchieye, F.; Ngoula, F. Liquid Storage of Ram Semen: Associated Damages and Improvement. Open J. Anim. Sci. 2021, 11, 473–500. [Google Scholar] [CrossRef]
- Evans, G.; Maxwell, W.M.C. Salamon’s Artifical Insemination of Sheep and Goats; Butterworth Pty Limited: Tennant Creek, Australia, 1987. [Google Scholar]
- Pini, T.; Leahy, T.; De Graaf, S.P. Sublethal Sperm Freezing Damage: Manifestations and Solutions. Theriogenology 2018, 118, 172–181. [Google Scholar] [CrossRef]
- Salamon, S.; Maxwell, W.M.C. Frozen Storage of Ram Semen I. Processing, Freezing, Thawing and Fertility after Cervical Insemination. Anim. Reprod. Sci. 1995, 37, 185–249. [Google Scholar] [CrossRef]
- White, I. Lipids and Calcium Uptake of Sperm in Relation to Cold Shock and Preservation: A Review. Reprod. Fertil. Dev. 1993, 5, 639. [Google Scholar] [CrossRef] [PubMed]
- Salamon, S.; Maxwell, W.M.C. Frozen Storage of Ram Semen II. Causes of Low Fertility after Cervical Insemination and Methods of Improvement. Anim. Reprod. Sci. 1995, 38, 1–36. [Google Scholar] [CrossRef]
- Maxwell, W.M.; Salamon, S. Liquid Storage of Ram Semen: A Review. Reprod. Fertil. Dev. 1993, 5, 613–638. [Google Scholar] [CrossRef]
- Sathe, S.R. Laparoscopic Artificial Insemination Technique in Small Ruminants—A Procedure Review. Front. Vet. Sci. 2018, 5, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wusiman, A.; Wang, Y.-P.; Ren, K.; Zhou, G.-B.; Fu, X.-W.; Suo, L.; Fan, Z.-Q.; Wang, L.; Zhu, S.-E. Semen Storage at 23, 4 or -196c and Its Applications to Artifical Inseminatio in Small-Tail Han Sheep. Asian J. Anim. Vet. Adv. 2012, 7, 299–308. [Google Scholar] [CrossRef] [Green Version]
- Borza, C.; Muntean, D.; Dehelean, C.; Savoiu, G.; Serban, C.; Simu, G.; Andoni, M.; Butur, M.; Drag, S. Oxidative Stress and Lipid Peroxidation—A Lipid Metabolism Dysfunction; InTech: Rijeka, Croatia, 2013. [Google Scholar]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and Consequences of Oxidative Stress in Spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. Oxidative Stress, Sperm Survival and Fertility Control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef]
- Bruemmert, J.E.; Coy, R.C.; Squires, E.L.; Graham, J.K. Effect of Pyruvate on the Function of Stallion Spermatozoa Stored for up to 48 Hours1. J. Anim. Sci. 2002, 80, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Casao, A.; Cebrián, I.; Asumpção, M.E.; Pérez-Pé, R.; Abecia, J.A.; Forcada, F.; Cebrián-Pérez, J.A.; Muiño-Blanco, T. Seasonal Variations of Melatonin in Ram Seminal Plasma Are Correlated to Those of Testosterone and Antioxidant Enzymes. Reprod. Biol. Endocrinol. 2010, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dai, G.-C.; Meng, Y.; Zhang, L.-K.; Du, Y.-Q.; Wen, F.; Feng, T.-Y.; Hu, J.-H. Effect of Addition of Melatonin on Liquid Storage of Ram Semen at 4 °C. Andrologia 2019, 51, e13236. [Google Scholar] [CrossRef] [PubMed]
- Nagina, G.; Asima, A.; Nemat, U.; Shamim, A. Effect of Melatonin on Maturation Capacity and Fertilization of Nili-Ravi Buffalo (Bubalus Bubalis) Oocytes. Open Vet. J. 2016, 6, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paulenz, H.; Söderquist, L.; Pérez-Pé, R.; Berg, K.A. Effect of Different Extenders and Storage Temperatures on Sperm Viability of Liquid Ram Semen. Theriogenology 2002, 57, 823–836. [Google Scholar] [CrossRef]
- Kaabi, M.; Paz, P.; Alvarez, M.; Anel, E.; Boixo, J.C.; Rouissi, H.; Herraez, P.; Anel, L. Effect of Epididymis Handling Conditions on the Quality of Ram Spermatozoa Recovered Post-Mortem. Theriogenology 2003, 60, 1249–1259. [Google Scholar] [CrossRef]
- Alomar, M.; Zarkawi, M.; Alzoabi, M.A. Analysis of Awassi Sperm Motility in Two Media at Different Levels of Temperature, PH and Osmolality. Iran. J. Appl. Anim. Sci. 2018, 8, 431–438. [Google Scholar]
- Murphy, E.M.; Murphy, C.; O’Meara, C.; Dunne, G.; Eivers, B.; Lonergan, P.; Fair, S. A Comparison of Semen Diluents on the in Vitro and in Vivo Fertility of Liquid Bull Semen. J. Dairy Sci. 2017, 100, 1541–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, J.-H.; Li, Y.-W.; Xie, H.-L.; Li, Q.; Dong, H.-B.; Sun, M.-J.; Gao, W.-Q.; Tan, J.-H. Effects of Glucose Metabolism Pathways on Sperm Motility and Oxidative Status during Long-Term Liquid Storage of Goat Semen. Theriogenology 2016, 86, 839–849. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Arango, J.; Nivola, K.; Väihkönen, L.; Katila, T. The Effect of Storage Temperature of Stallion Semen on Pregnancy Rates. J. Equine Vet. Sci. 2015, 35, 611–616. [Google Scholar] [CrossRef]
- Province, C.A.; Squires, E.L.; Pickett, B.W.; Amann, R.P. Cooling Rates, Storage Temperatures and Fertility of Extended Equine Spermatozoa. Theriogenology 1985, 23, 925–934. [Google Scholar] [CrossRef]
- Holt, W.V.; North, R.D. Partially Irreversible Cold-Induced Lipid Phase Transitions in Mammalian Sperm Plasma Membrane Domains: Freeze-Fracture Study. J. Exp. Zool. 1984, 230, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.F. The Causes of Reduced Fertility with Cryopreserved Semen. Anim. Reprod. Sci. 2000, 60–61, 481–492. [Google Scholar] [CrossRef]
- Maxwell, W.M.C.; Johnson, L.A. Membrane Status of Boar Spermatozoa after Cooling or Cryopreservation. Theriogenology 1997, 48, 209–219. [Google Scholar] [CrossRef]
- Blackshaw, A.W. The Prevention of Temperature Shock of Bull and Ram Semen. Aust. J. Biol. Sci. 1954, 7, 573–582. [Google Scholar] [CrossRef] [Green Version]
- Pursel, V.G.; Johnson, L.A.; Schulman, L.L. Effect of Dilution, Seminal Plasma and Incubation Period on Cold Shock Susceptibility of Boar Spermatozoa. J. Anim. Sci. 1973, 37, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Flesch, F.M.; Brouwers, J.F.H.M.; Nievelstein, P.; Verkleij, A.J.; van Golde, L.M.G.; Colenbrander, B.; Gadella, B.M. Bicarbonate Stimulated Phospholipid Scramblinginduces Cholesterol Redistribution and Enablescholesterol Depletion in the Sperm Plasma Membrane. J. Cell Sci. 2001, 114, 3543–3555. [Google Scholar] [CrossRef] [PubMed]
- Kessel, A.; Ben-Tal, N.; May, S. Interactions of Cholesterol with Lipid Bilayers: The Preferred Configuration and Fluctuations. Biophys. J. 2001, 81, 643–658. [Google Scholar] [CrossRef] [Green Version]
- Cross, N.L. Role of Cholesterol in Sperm Capacitation1. Biol. Reprod. 1998, 59, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parks, J.E.; Lynch, D.V. Lipid Composition and Thermotropic Phase Behavior of Boar, Bull, Stallion, and Rooster Sperm Membranes. Cryobiology 1992, 29, 255–266. [Google Scholar] [CrossRef]
- Moore, A.I.; Squires, E.L.; Graham, J.K. Adding Cholesterol to the Stallion Sperm Plasma Membrane Improves Cryosurvival. Cryobiology 2005, 51, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Labbé, C.; Bussière, J.-F.; Guillouet, P.; Leboeuf, B.; Magistrini, M. Cholesterol/Phospholipid Ratio in Sperm of Several Domestic Species Does Not Directly Predict Sperm Fitness for Cryopreservation. Genet. Sel. Evol. 2001, 33, S61. [Google Scholar] [CrossRef] [Green Version]
- Peker akalin, P.; Bucak, M.N.; Güngör, Ş.; Başpinar, N.; Çoyan, K.; Dursun, Ş.; İli, P.; Aksoy, A.; Karaşör, Ö.F.; Bilgili, A.; et al. Influence of Lycopene and Cysteamine on Sperm and Oxidative Stress Parameters during Liquid Storage of Ram Semen at 5 °C. Small Rumin. Res. 2016, 137, 117–123. [Google Scholar] [CrossRef]
- Maxwell, W.M.C.; Johnson, L.A. Physiology of Spermatozoa at High Dilution Rates: The Influence of Seminal Plasma. Theriogenology 1999, 52, 1353–1362. [Google Scholar] [CrossRef]
- Harrison, R.a.P.; Dott, H.M.; Foster, G.C. Bovine Serum Albumin, Sperm Motility, and the “Dilution Effect”. J. Exp. Zool. 1982, 222, 81–88. [Google Scholar] [CrossRef]
- Leahy, T.; Marti, J.I.; Mendoza, N.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Evans, G.; Maxwell, W.M.C. High Pre-Freezing Dilution Improves Post-Thaw Function of Ram Spermatozoa. Anim. Reprod. Sci. 2010, 119, 137–146. [Google Scholar] [CrossRef]
- Castellini, C.; Lattaioli, P.; Moroni, M.; Minelli, A. Effect of Seminal Plasma on the Characteristics and Fertility of Rabbit Spermatozoa. Anim. Reprod. Sci. 2000, 63, 275–282. [Google Scholar] [CrossRef]
- Garner, D.L.; Thomas, C.A.; Gravance, C.G.; Marshall, C.E.; DeJarnette, J.M.; Allen, C.H. Seminal Plasma Addition Attenuates the Dilution Effect in Bovine Sperm. Theriogenology 2001, 56, 31–40. [Google Scholar] [CrossRef]
- Patil, S.; Kumar, P.; Singh, G.; Bala, R.; Jerome, A.; Patil, C.S.; Kumar, D.; Singh, S.; Sharma, R.K. ‘Semen Dilution Effect’ on Sperm Variables and Conception Rate in Buffalo. Anim. Reprod. Sci. 2020, 214, 106304. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, S.; Gallego, R.D.; Garcia-Colomer, B.; Gomez, E.A.; Yaniz, J.; Gosalvez, J.; Lopez- Fernandez, C.; Silvestre, M.A. Effect of Sperm Concentration and StorageTemperature on Goat Spermatozoa DuringLiquid Storage. Biology 2020, 9, 300. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Vázquez, J.M.; García, E.M.; Roca, J.; Martínez, E.A.; Calvete, J.J.; Sanz, L.; Ekwall, H.; Rodríguez-Martínez, H. Immunolocalization and Possible Functional Role of PSP-I/PSP-II Heterodimer in Highly Extended Boar Spermatozoa. J. Androl. 2006, 27, 766–773. [Google Scholar] [CrossRef] [Green Version]
- Forouzanfar, M.; Sharafi, M.; Hosseini, S.M.; Ostadhosseini, S.; Hajian, M.; Hosseini, L.; Abedi, P.; Nili, N.; Rahmani, H.R.; Nasr-Esfahani, M.H. In Vitro Comparison of Egg Yolk–Based and Soybean Lecithin–Based Extenders for Cryopreservation of Ram Semen. Theriogenology 2010, 73, 480–487. [Google Scholar] [CrossRef]
- Albiaty, N.M.H.; Alobaidi, H.J.K.; Kareem, A.F.; Al-Hakim, A.M.; Alnaed, A.; Alkhazraji, A.A.H. Effect of Extenders and Preservation Periods in Some Semen Characteristics of Awassi Ram. World J. Pharm. Res. 2016, 5, 234–243. [Google Scholar]
- Arando, A.; Delgado, J.V.; León, J.M.; Nogales, S.; Navas-González, F.J.; Pizarro, M.G.; Pérez-Marín, C.C. Effect of Three Commercial Extenders on Sperm Motility and Fertility in Liquid Ram Semen Stored at 15 °C or 5 °C. Acta Vet. Hung. 2019, 67, 430–444. [Google Scholar] [CrossRef]
- Arando, A.; Gonzalez, A.; Delgado, J.V.; Arrebola, F.A.; Perez-Marín, C.C. Storage Temperature and Sucrose Concentrations Affect Ram Sperm Quality after Vitrification. Anim. Reprod. Sci. 2017, 181, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Stojanov, T. Studies on the Liquid Storage of Ram Spermatozoa in the Presnce or Absence of Antioxidants. Ph.D. Thesis, University of Sydney, Sydney, Australia, 1995. [Google Scholar]
- Bergeron, A.; Crête, M.-H.; Brindle, Y.; Manjunath, P. Low-Density Lipoprotein Fraction from Hen’s Egg Yolk Decreases the Binding of the Major Proteins of Bovine Seminal Plasma to Sperm and Prevents Lipid Efflux from the Sperm Membrane1. Biol. Reprod. 2004, 70, 708–717. [Google Scholar] [CrossRef] [Green Version]
- Yaniz, J.; Marco-Aguado, M.-A.; Mateos, J.A.; Santolaria, P. Bacterial Contamination of Ram Semen, Antibiotic Sensitivities, and Effects on Sperm Quailty during Storage at 15c. Anim. Reprod. Sci. 2010, 122, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, D.E.; Mercer, B.G.; Wiklendt, A.M.; Aitken, R.J. Quantitative Analysis of Gene-Specific DNA Damage in Human Spermatozoa. Mutat. Res. Mol. Mech. Mutagen. 2003, 529, 21–34. [Google Scholar] [CrossRef]
- Gibb, Z.; Lambourne, S.R.; Quadrelli, J.; Smith, N.D.; Aitken, R.J. L-Carnitine and Pyruvate Are Prosurvival Factors during the Storage of Stallion Spermatozoa at Room Temperature. Biol. Reprod. 2015, 93, 104. [Google Scholar] [CrossRef] [PubMed]
- Drobnis, E.Z.; Crowe, L.M.; Berger, T.; Anchordoguy, T.J.; Overstreet, J.W.; Crowe, J.H. Cold Shock Damage Is Due to Lipid Phase Transitions in Cell Membranes: A Demonstration Using Sperm as a Model. J. Exp. Zool. 1993, 265, 432–437. [Google Scholar] [CrossRef] [PubMed]
- Leeuw, F.E.D.; Chen, H.-C.; Colenbrander, B.; Verkleij, A.J. Cold-Induced Ultrastructural Changes in Bull and Boar Sperm Plasma Membranes. Cryobiology 1990, 27, 171–183. [Google Scholar] [CrossRef]
- Holt, W.V.; North, R.D. Thermotropic Phase Transitions in the Plasma Membrane of Ram Spermatozoa. Reproduction 1986, 78, 447–457. [Google Scholar] [CrossRef]
- Sanocka, D.; Kurpisz, M. Reactive Oxygen Species and Sperm Cells. Reprod. Biol. Endocrinol. RBE 2004, 2, 12. [Google Scholar] [CrossRef] [Green Version]
- Nevo, A.C.; Rikmenspoel, R. Diffusion of ATP in Sperm Flagella. J. Theor. Biol. 1970, 26, 11–18. [Google Scholar] [CrossRef]
- Plaza Davila, M.; Martín, P.; Tapia, J.; Ferrusola, O.; Balão da Silva, C.; Peña, F. Inhibition of Mitochondrial Complex I Leads to Decreased Motility and Membrane Integrity Related to Increased Hydrogen Peroxide and Reduced ATP Production, While the Inhibition of Glycolysis Has Less Impact on Sperm Motility. PLoS ONE 2015, 10, e0138777. [Google Scholar] [CrossRef] [Green Version]
- Losano, J.; Angrimani, D.; Dalmazzo, A.; Rui, B.; Brito, M.; Mendes, C.; Kawai, G.; Vannucchi, C.; Assumpção, M.; Barnabe, V.; et al. Effect of Mitochondrial Uncoupling and Glycolysis Inhibition on Ram Sperm Functionality. Reprod. Domest. Anim. 2017, 52, 289–297. [Google Scholar] [CrossRef]
- Bilodeau, J.-F.; Chatterjee, S.; Sirard, M.A.; Gagnon, C. Levels of Antioxidant Defenses Are Decreased in Bovine Spermatozoa After a Cycle of Freezing and Thawing. Mol. Reprod. Dev. 2000, 55, 282–288. [Google Scholar] [CrossRef]
- Tourmente, M.; Villar-Moya, P.; Rial, E.; Roldan, E.R.S. Differences in ATP Generation Via Glycolysis and Oxidative Phosphorylation and Relationships with Sperm Motility in Mouse Species. J. Biol. Chem. 2015, 290, 20613–20626. [Google Scholar] [CrossRef] [Green Version]
- Aitken, J.; Fisher, H. Reactive Oxygen Species Generation and Human Spermatozoa: The Balance of Benefit and Risk. BioEssays 1994, 16, 259–267. [Google Scholar] [CrossRef]
- Srivastava, S.; Chandra, A.; Bhatnagar, A.; Srivastava, S.K.; Ansari, N.H. Lipid Peroxidation Product, 4-Hydroxynonenal and Its Conjugate with GSH Are Excellent Substrates of Bovine Lens Aldose Reductase. Biochem. Biophys. Res. Commun. 1995, 217, 741–746. [Google Scholar] [CrossRef]
- Fritz, K.S.; Petersen, D.R. Exploring the Biology of Lipid Peroxidation-Derived Protein Carbonylation. Chem. Res. Toxicol. 2011, 24, 1411–1419. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative Stress and Sperm Function: A Systematic Review on Evaluation and Management. Arab J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Gibb, Z.; Mitchell, L.A.; Lambourne, S.R.; Connaughton, H.S.; De Iuliis, G.N. Sperm Motility Is Lost In Vitro as a Consequence of Mitochondrial Free Radical Production and the Generation of Electrophilic Aldehydes but Can Be Significantly Rescued by the Presence of Nucleophilic Thiols. Biol. Reprod. 2012, 87. [Google Scholar] [CrossRef] [Green Version]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, Y.P.; Vanha-Perttula, T. Glutathione, L-Glutamic Acid and γ-Glutamyl Transpeptidase in the Bull Reproductive Tissues. Int. J. Androl. 1988, 11, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Ntemka, A.; Tsakmakidis, I.A.; Kiossis, E.; Milovanović, A.; Boscos, C.M. Current Status and Advances in Ram Semen Cryopreservation. J. Hell. Vet. Med. Soc. 2018, 69, 911. [Google Scholar] [CrossRef] [Green Version]
- Câmara, D.R.; Silva, S.V.; Almeida, F.C.; Nunes, J.F.; Guerra, M.M.P. Effects of Antioxidants and Duration of Pre-Freezing Equilibration on Frozen-Thawed Ram Semen. Theriogenology 2011, 76, 342–350. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Hamilton, T.R.; de Castro, L.S.; de Carvalho Delgado, J.; de Assis, P.M.; Siqueira, A.F.P.; Mendes, C.M.; Goissis, M.D.; Muiño-Blanco, T.; Cebrián-Pérez, J.Á.; Nichi, M.; et al. Induced Lipid Peroxidation in Ram Sperm: Semen Profile, DNA Fragmentation and Antioxidant Status. Reproduction 2016, 151, 379–390. [Google Scholar] [CrossRef]
- Ashworth, P.J.; Harrison, R.A.; Miller, N.G.; Plummer, J.M.; Watson, P.F. Survival of Ram Spermatozoa at High Dilution: Protective Effect of Simple Constituents of Culture Media as Compared with Seminal Plasma. Reprod. Fertil. Dev. 1994, 6, 173–180. [Google Scholar] [CrossRef]
- Amidi, F.; Pazhohan, A.; Shabani Nashtaei, M.; Khodarahmian, M.; Nekoonam, S. The Role of Antioxidants in Sperm Freezing: A Review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Gharagozloo, P.; Aitken, R.J. The Role of Sperm Oxidative Stress in Male Infertility and the Significance of Oral Antioxidant Therapy. Hum. Reprod. 2011, 26, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Allai, L.; Benmoula, A.; Marciane Da Silva, M.; Nasser, B.; El Amiri, B. Supplementation of Ram Semen Extender to Improve Seminal Quality and Fertility Rate. Anim. Reprod. Sci. 2018, 192, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Vernon, R.G.; Go, V.L.; Fritz, I.B. Studies on Spermatogenesis in Rats. II. Evidence That Carnitine Acetyltransferase Is a Marker Enzyme for the Investigation of Germ Cell Differentiation. Can. J. Biochem. 1971, 49, 761–767. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.E. Carnitine, Acetylcarnitine and the Activity of Carnitine Acyltransferases in Seminal Plasma and Spermatozoa of Men, Rams and Rats. Reproduction 1979, 56, 667–673. [Google Scholar] [CrossRef] [Green Version]
- Day-Francesconi, M.; Casillas, E.R. The Intracellular Localization and Properties of Carnitine Acetyltransferase from Ram Spermatozoa. Arch. Biochem. Biophys. 1982, 215, 206–214. [Google Scholar] [CrossRef]
- Pruneda, A.; Yeung, C.-H.; Bonet, S.; Pinart, E.; Cooper, T.G. Concentrations of Carnitine, Glutamate and Myo-Inositol in Epididymal Fluid and Spermatozoa from Boars. Anim. Reprod. Sci. 2007, 97, 344–355. [Google Scholar] [CrossRef]
- Solarska, K.; Lewińska, A.; Karowicz-Bilińska, A.; Bartosz, G. The Antioxidant Properties of Carnitine in Vitro. Cell. Mol. Biol. Lett. 2010, 15. [Google Scholar] [CrossRef]
- Yang, K.; Wang, N.; Guo, H.-T.; Wang, J.-R.; Sun, H.-H.; Sun, L.-Z.; Yue, S.-L.; Zhou, J.-B. Effect of L-Carnitine on Sperm Quality during Liquid Storage of Boar Semen. Asian-Australas. J. Anim. Sci. 2019, 33, 1763. [Google Scholar] [CrossRef]
- Aliabadi, E.; Soleimani Mehranjani, M.; Borzoei, Z.; Talaei-Khozani, T.; Mirkhani, H.; Tabesh, H. Effects of L-Carnitine and L-Acetyl-Carnitine on Testicular Sperm Motility and Chromatin Quality. Iran. J. Reprod. Med. 2012, 10, 77–82. [Google Scholar] [PubMed]
- Deana, R.; Rigoni, F.; Francesconi, M.; Cavallini, L.; Arslan, P.; Siliprandi, N. Effect of L-Carnitlne and L-Aminocarnitine on Calcium Transport, Motility, and Enzyme Release from Ejaculated Bovine Spermatozoa. Biol. Reprod. 1989, 41, 949–955. [Google Scholar] [CrossRef] [Green Version]
- Galarza, D.A.; Guevara, M.L.D.; Beltrán-Breña, P.; Sánchez-Calabuig, M.J.; Rizos, D.; López-Sebastián, A.; Santiago-Moreno, J. Influence of Sperm Filtration and the Addition of Glycerol to UHT Skimmed Milk- and TEST-Based Extenders on the Quality and Fertilizing Capacity of Chilled Ram Sperm. Theriogenology 2019, 133, 29–37. [Google Scholar] [CrossRef]
- Galarza, D.A.; Santiago-Moreno, J.; Sebastian, L. Effect of L-Carnitine in a Skimmed Milk Based-Diluent on Membranes Preservation and Kinetic Activity of Ram Sperm under Chilled Conditions; Zaragoza. 2019. Available online: https://www.cabdirect.org/cabdirect/welcome/?target=%2fcabdirect%2fabstract%2f20193404681 (accessed on 24 November 2021).
- Souza, C.V.D.; Brandão, F.Z.; Santos, J.D.R.; Alfradique, V.A.P.; Santos, V.M.B.D.; Morais, M.C.D.C.; Rangel, P.S.C.; Silva, A.A.D.; Souza-Fabjan, J.M.G. Effect of Different Concentrations of L-Carnitine in Extender for Semen Cryopreservation in Sheep. Cryobiology 2019, 89, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.M.; Shi, L.Z.; Tam, J.; Chandsawangbhuwana, C.; Durrant, B.; Botvinick, E.L.; Berns, M.W. Comparison of Glycolysis and Oxidative Phosphorylation as Energy Sources for Mammalian Sperm Motility, Using the Combination of Fluorescence Imaging, Laser Tweezers, and Real-Time Automated Tracking and Trapping. J. Cell. Physiol. 2008, 217, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Hereng, T.H.; Elgstøen, K.B.P.; Cederkvist, F.H.; Eide, L.; Jahnsen, T.; Skålhegg, B.S.; Rosendal, K.R. Exogenous Pyruvate Accelerates Glycolysis and Promotes Capacitation in Human Spermatozoa. Hum. Reprod. 2011, 26, 3249–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz-Rodríguez, J.M.; Martín-Cano, F.E.; Gaitskell-Phillips, G.L.; Silva, A.; Ortega-Ferrusola, C.; Gil, M.C.; Peña, F.J. Low Glucose and High Pyruvate Reduce the Production of 2-Oxoaldehydes, Improving Mitochondrial Efficiency, Redox Regulation, and Stallion Sperm Function†. Biol. Reprod. 2021, 2021, ioab073. [Google Scholar] [CrossRef]
- Giandomenico, A.R.; Cerniglia, G.E.; Biaglow, J.E.; Stevens, C.W.; Koch, C.J. The Importance of Sodium Pyruvate in Assessing Damage Produced by Hydrogen Peroxide. Free Radic. Biol. Med. 1997, 23, 426–434. [Google Scholar] [CrossRef]
- Upreti, G.C.; Oliver, J.E.; Duganzich, D.M.; Munday, R.; Smith, J.F. Development of a Chemically Defined Ram Semen Diluent (RSD-1). Anim. Reprod. Sci. 1995, 37, 143–157. [Google Scholar] [CrossRef]
- Wang, H.; Lu, J.; Kulkarni, S.; Zhang, W.; Gorka, J.E.; Mandel, J.A.; Goetzman, E.S.; Prochownik, E.V. Metabolic and Oncogenic Adaptations to Pyruvate Dehydrogenase Inactivation in Fibroblasts. J. Biol. Chem. 2019, 294, 5466–5486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darr, C.R.; Varner, D.D.; Teague, S.; Cortopassi, G.A.; Datta, S.; Meyers, S.A. Lactate and Pyruvate Are Major Sources of Energy for Stallion Sperm with Dose Effects on Mitochondrial Function, Motility, and ROS Production. Biol. Reprod. 2016, 95, 34. [Google Scholar] [CrossRef]
- Ashrafi, I.; Kohram, H.; Naijian, H.; Bahreini, M.; Poorhamdollah, M. Protective Effect of Melatonin on Sperm Motility Parameters on Liquid Storage of Ram Semen at 5 °C. Afr. J. Biotechnol. 2011, 10, 6670–6674. [Google Scholar]
- Jang, H.; Kim, Y.; Kim, B.; Park, I.; Cheong, H.; Kim, J.; Park, C.; Kong, H.; Lee, H.; Yang, B. Ameliorative Effects of Melatonin against Hydrogen Peroxide-Induced Oxidative Stress on Boar Sperm Characteristics and SubsequentIn VitroEmbryo Development. Reprod. Domest. Anim. 2010, 45, 943–950. [Google Scholar] [CrossRef]
- Balao de Silva, C.M.; Macias-Garcia, B.; Miro-Moran, A.; Gonzalez-Fernandez, L.; Morillo-Rodriguez, A.; Ortega- Fervisola, C.; Gallardo- Balanos, J.M.; Stillwell, G.; Topea, J.A.; Pena, F.J. Melatonin Reduces Lipid Peroxidation and Apoptotic-like Changes in Stallion Spermatozoa. J. Pineal Res. 2011, 51, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Succu, S.; Berlinguer, F.; Pasciu, V.; Satta, V.; Leoni, G.G.; Naitana, S. Melatonin Protects Ram Spermatozoa from Cryopreservation Injuries in a Dose-Dependent Manner: Melatonin Mitigates Cryopreservation Injuries of Ram Spermatozoa. J. Pineal Res. 2011, 50, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.; Manchester, L.; Qi, W. Biochemical Reactvity of Melatonin with Oxygen and Nitrogen Species. Cell Biochem. Biophys. 2001, 34, 237–256. [Google Scholar]
- Li, C.-Y.; Hao, H.-S.; Zhao, Y.-H.; Zhang, P.-P.; Wang, H.-Y.; Pang, Y.-W.; Du, W.-H.; Zhao, S.-J.; Liu, Y.; Huang, J.-M.; et al. Melatonin Improves the Fertilization Capacity of Sex-Sorted Bull Sperm by Inhibiting Apoptosis and Increasing Fertilization Capacitation via MT1. Int. J. Mol. Sci. 2019, 20, 3921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Plessis, S.S.; Hagenaar, K.; Lampiao, F. Thein Vitroeffects of Melatonin on Human Sperm Function and Its Scavenging Activities on NO and ROS. Andrologia 2010, 42, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Casao, A.; Mendoza, N.; Pérez-Pé, R.; Grasa, P.; Abecia, J.-A.; Forcada, F.; Cebrián-Pérez, J.A.; Muino-Blanco, T. Melatonin Prevents Capacitation and Apoptotic-like Changes of Ram Spermatozoa and Increases Fertility Rate. J. Pineal Res. 2010, 48, 39–46. [Google Scholar] [CrossRef]
- Martin-Hildago, D.; Baron, F.J.; Bragado, M.J.; Carmona, P.; Robina, A.; Garcia- Marin, L.; Gil, M.C. The Effect of Melatonin on the Quality of Extended Boar Semen after Long-Term Storage at 17 °C. Theriogenology 2011, 75, 1550–1560. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.W.; Kleszczyński, K.; Hardkop, L.H.; Kruse, N.; Zillikens, D. Melatonin Enhances Antioxidative Enzyme Gene Expression (CAT, GPx, SOD), Prevents Their UVR-Induced Depletion, and Protects against the Formation of DNA Damage (8-Hydroxy-2’-Deoxyguanosine) in Ex Vivo Human Skin. J. Pineal Res. 2013, 54, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Gibb, Z.; Aitken, R.J. Recent Developments in Stallion Semen Preservation. J. Equine Vet. Sci. 2016, 43, S29–S36. [Google Scholar] [CrossRef]
- MaríN-Briggiler, C.I.; Tezón, J.G.; Miranda, P.V.; Vazquez-Levin, M.H. Effect of Incubating Human Sperm at Room Temperature on Capacitation-Related Events. Fertil. Steril. 2002, 77, 252–259. [Google Scholar] [CrossRef]
- Kamada, Y.; Wakayama, S.; Shibasaki, I.; Ito, D.; Kamimura, S.; Ooga, M.; Wakayama, T. Assessing the Tolerance to Room Temperature and Viability of Freeze-Dried Mice Spermatozoa over Long-Term Storage at Room Temperature under Vacuum. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Pauw, I.M.C.D.; Soom, A.V.; Mintiens, K.; Verberckmoes, S.; Kruif, A.D. In Vitro Survival of Bovine Spermatozoa Stored at Room Temperature under Epididymal Conditions. Theriogenology 2003, 59, 1093–1107. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Temp | Extender | Result | Reference |
|---|---|---|---|---|
| Ram | 4 °C 23 °C −196 °C | Citrate–glucose–EY 1 | Higher fertility at 23 °C and 4 °C when compared to −196 °C at 24 h | [12] |
| 5 °C 20 °C | Milk-based Sodium-citrate-based Tris–citrate–fructose + EY 1 | Sodium-based and TRIS with and without EY maintained the highest viability at 72 h | [20] | |
| 5 °C 24 °C | Tes–Tris–fructose solution with EY * | Higher sperm abnormality in sperm stored at 24 °C when compared to 5 °C at 48 h | [21] | |
| Higher embryo cleavage at 24 °C when compared to 5 °C at 48 h | ||||
| 4 °C 20 °C 37 °C | Tyrode’s albumin lactate pyruvate + EY 1 | Higher TM at 20 °C and 37 °C when compared to 4 °C at 72 h | [22] | |
| Bull | 5 °C 18 °C | Cap 1 BioXcell INRA96 | Higher TM in Cap extender when compared to BioXcell and INRA96 at 72 h | [23] |
| Buck | 5 °C 15 °C 25 °C | PBS supplemented with 10 mM pyruvate or lactase | Higher PM at 15 °C when compared to 5 °C and 25 °C at 168 h | [24] |
| Higher viability at 15 °C when compared to 5 °C and 25 °C at 168 h | ||||
| Stallion | 15–20 °C | INRA96 | Lower TM at 20 °C when compared to 5 °C at 12 h | [25] |
| 5 °C 15 °C 20 °C | SM 1 Cap 1 NFMS 1 | Higher fertility PM in Cap than SM and NFM at all temperatures at 72 h | [26] |
| Species | Cholesterol (mol %) | Ratio of Cholesterol: Phospholipid 1 | Resistance to Cold Shock | Reference |
|---|---|---|---|---|
| Ram | 27 | 0.43 | Low | [34] |
| Bull | 31 | 0.45 | High | [35] |
| Stallion | Not reported | 0.36 | Low | [36] |
| Boar | Not reported | 0.37 | Partial | [37] |
| Phase Transition Temp | Species | Metabolic Changes | Physiological Changes |
|---|---|---|---|
| 30–36 °C * | Ram | Intramembrane lipid scattering | Membrane fusogenicity |
| Phosphate discontinuity | Increased protein immobility and reconfiguration | ||
| 23–26 °C | Ram | Calcium transport discontinuity | ATPase activity reduced |
| Intramembrane lipid scattering and agglutination | |||
| 23 °C and 14 °C | Boar | Reduction in -CH2 absorbance | Inhibition of partial acquisition of cold shock resistance |
| 18 °C | Boar | 40% increase in potassium efflux | Acceleration of ATPase |
| 17 °C | Ram | Intramembrane lipid scattering | Membrane particle redistribution/aggregation Ultrastructural freeze fractures |
| Calcium influx | Capacitation initiated Irreversible membrane leakiness |
| Species | Temp (°C) | Extender | Result | Reference |
|---|---|---|---|---|
| Boar | 17 | Androhep | Higher membrane integrity at 50 mM when compared to 12.5 mM, 25 mM, and 100 mM at ten days | [84] |
| Ram | 5 | Skimmed milk extender | Higher TM at 10 mM when compared to 1 mM, 2.5 mM, 5 mM, and 7.5 mM at 96 h | [88] |
| Stallions | 23 | MBWW | Higher MMP (%) at 0 mM than at 10 mM at 72 h | [55] |
| Higher PM at 0 mM when compared to 10 mM at 72 h | ||||
| Higher lipid peroxidation at 0 mM when compared to 10 mM at 72 h | ||||
| Higher ROS at 0 mM when compared to 10 mM at 72 h | ||||
| Bovine | 5 20 37 | Sodium chlorine extender | Lower PM at 20 mM LC and 20 mM ALC when compared to 0 mM and 20 mM NaCl at 72 h | [86] |
| Species | Temp (°C) | Extender | Result | Reference |
|---|---|---|---|---|
| Bovine | 38.5 | TALP 1 | Higher TM when compared to 1 mM, 2 mM, and 5 mM at 6 h | [63] |
| Higher intracellular ATP at 5 mM than at 1 mM at 6 h | ||||
| Higher extracellular ATP at 1 mM than 5 mM at 6 h | ||||
| Stallion | 18 | Tyrode’s | Higher TM at 1 mM when compared to 0 mM at 48 h | [92] |
| Higher MMP 1 at 1 mM when compared to 0 mM at 48 h | ||||
| Higher ROS at 1 mM when compared to 0 mM at 48 h | ||||
| 37 | MBWW 1 | Higher TM at 5.5 mM when compared to 0 mM at 1 h Higher ROS at 5.5 mM when compared to 0 mM at 1 h | [96] | |
| 23 | MBWW 1 | Highest TM at 10 mM when compared to 1.25 mM, 2.5 mM, 5 mM, and 20 mM at 72 h Highest PM at 10 mM when compared to 1.25 mM, 2.5 mM, 5 mM, and 20 mM at 72 h | [55] |
| Species | Temp (°C) | Extender | Result | Reference |
|---|---|---|---|---|
| Ram | 39 | Saline medium | Higher maturation rate at 1 μm than 10 nm and 100 pm at 3 h | [104] |
| Higher % of capacitated cells at 10 nm than 1 um and 100 pm at 3 h | ||||
| Higher fertilisation rate at 100 pm than 1 μm and 100 pm at 3 h | ||||
| 5 | TRIS extender + EY 1 | Higher PM at 1 mM than at 0.1 and 3 mM at 48 h | [97] | |
| 4 | Tris-based extender | Higher plasma membrane integrity at 0.1 mM than at 0.05 mM, 0.2 mM and 0.4 mM at 120 h | [18] | |
| Higher MDA at 0.4 mM than at 0.1 mM at 120 h | ||||
| Boar | 17 | VL 1 | Higher TM at 1 μM than at 0 μM at 7 h | [105] |
| Buffalo | 39 | TALP 1 | Higher fertilisation at 500 mM when compared to 250 mM and 1000 μM at 18 h | [19] |
| Higher TM at 50 pm when compared to 100 pm, 200 pm and 1 μm at 18 h |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rizkallah, N.; Chambers, C.G.; de Graaf, S.P.; Rickard, J.P. Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement. Animals 2022, 12, 244. https://doi.org/10.3390/ani12030244
Rizkallah N, Chambers CG, de Graaf SP, Rickard JP. Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement. Animals. 2022; 12(3):244. https://doi.org/10.3390/ani12030244
Chicago/Turabian StyleRizkallah, Natalie, Caitlin G. Chambers, Simon P. de Graaf, and Jessica P. Rickard. 2022. "Factors Affecting the Survival of Ram Spermatozoa during Liquid Storage and Options for Improvement" Animals 12, no. 3: 244. https://doi.org/10.3390/ani12030244