
Early Pregnancy in Jennies in the Caribbean: Corpus Luteum Development and Progesterone Production, Uterine and Embryo Dynamics, Conceptus Growth and Maturation

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Progesterone Analysis
2.3. Statistical Analyses
3. Results
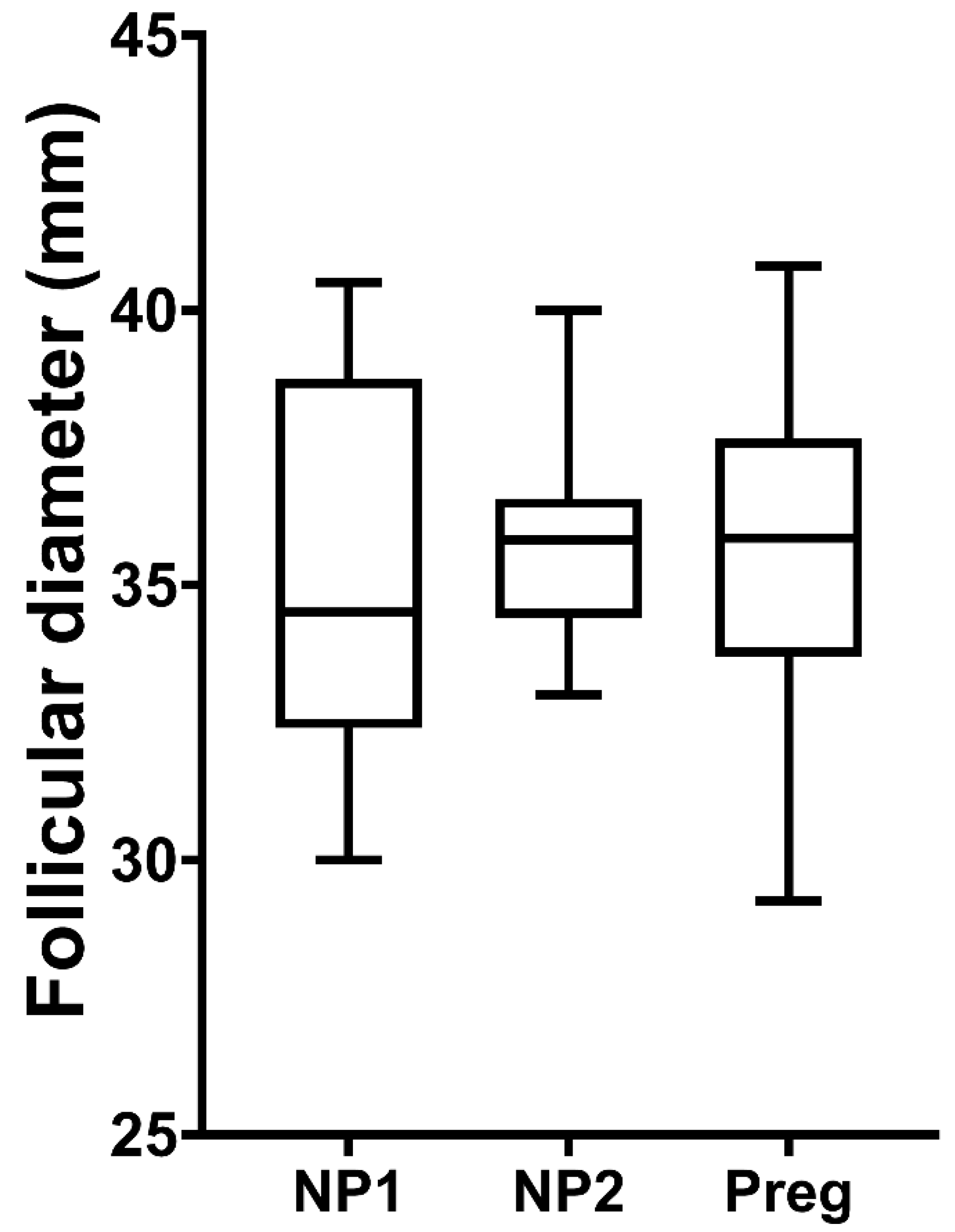
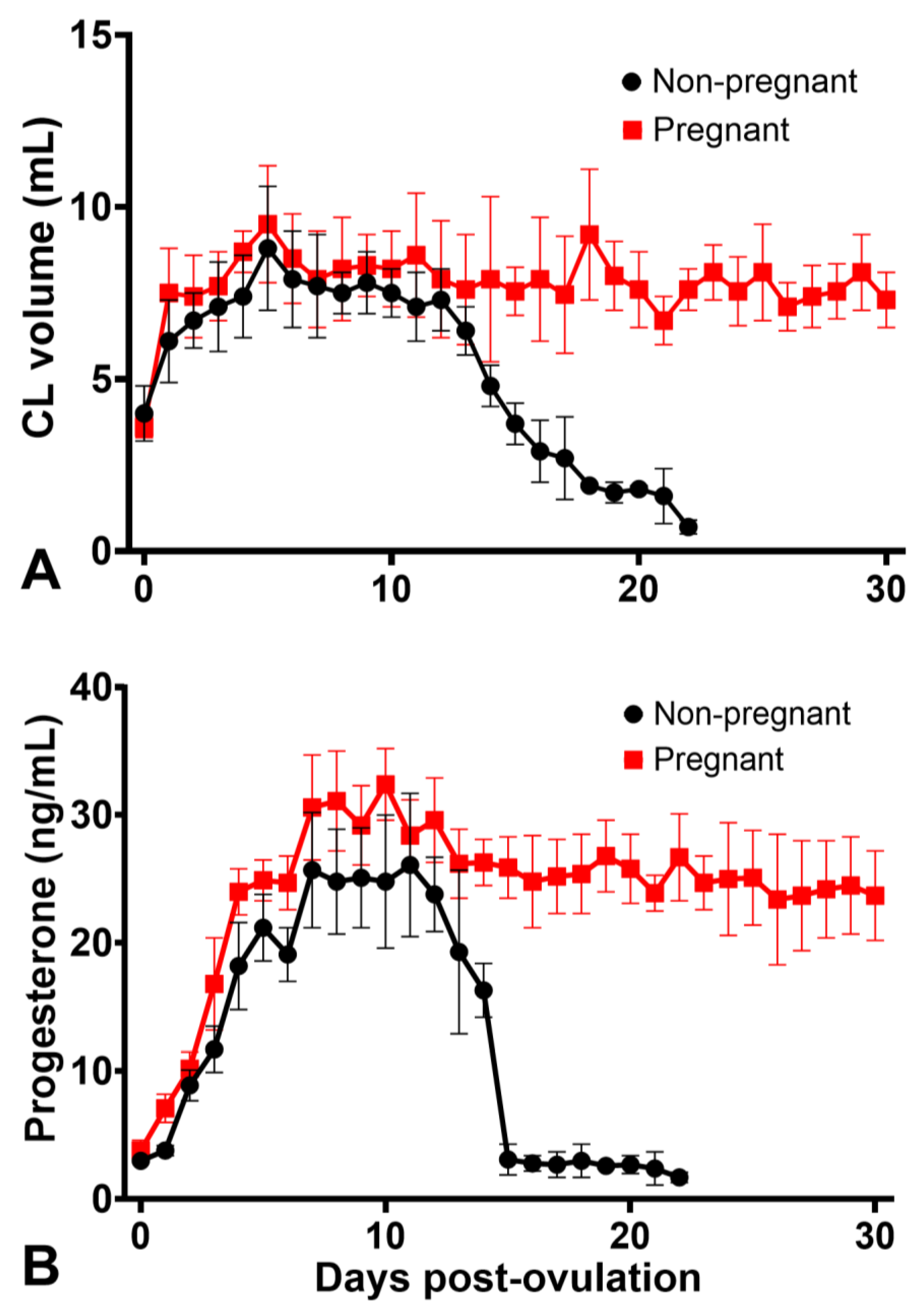
3.1. Corpus Luteum and Progesterone Profiles
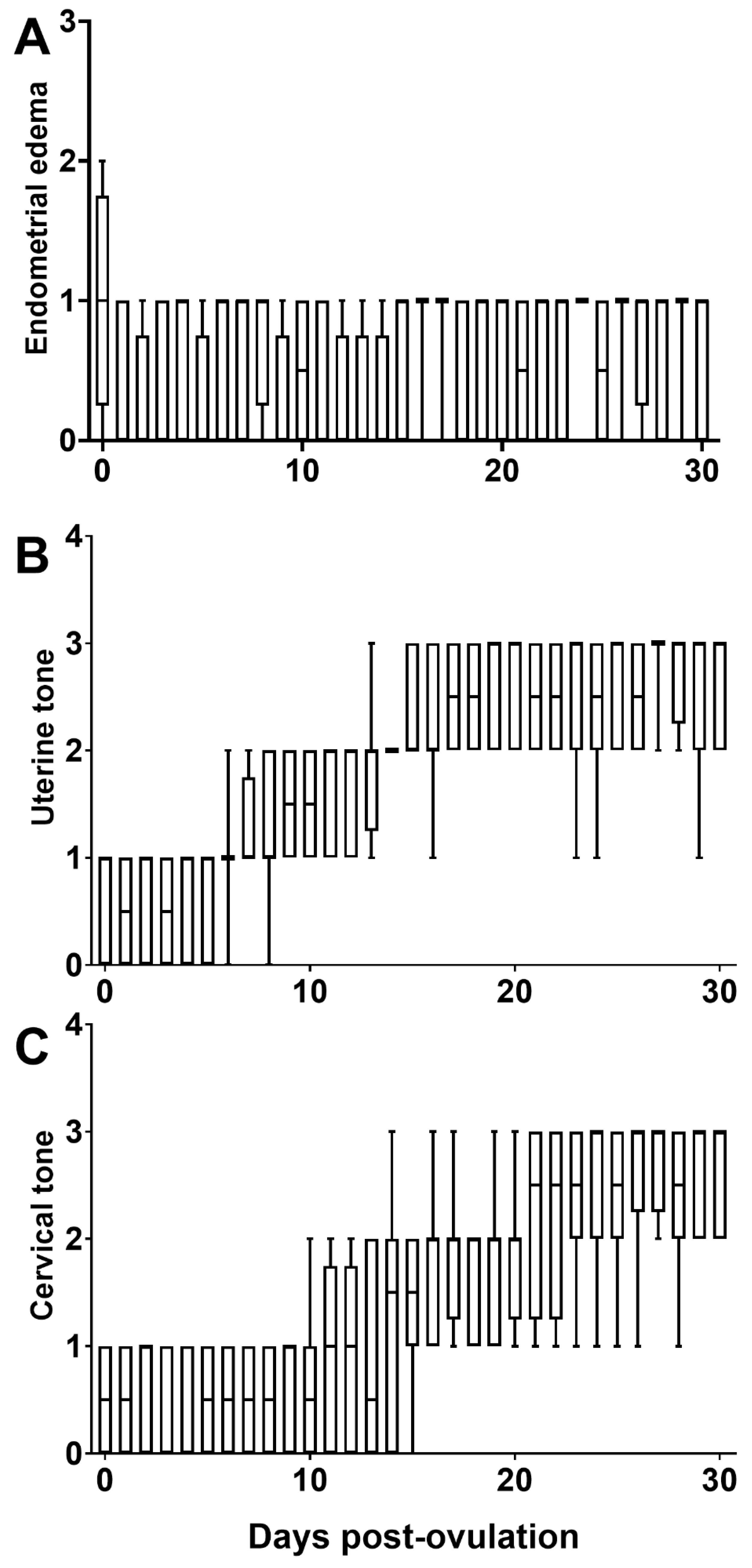
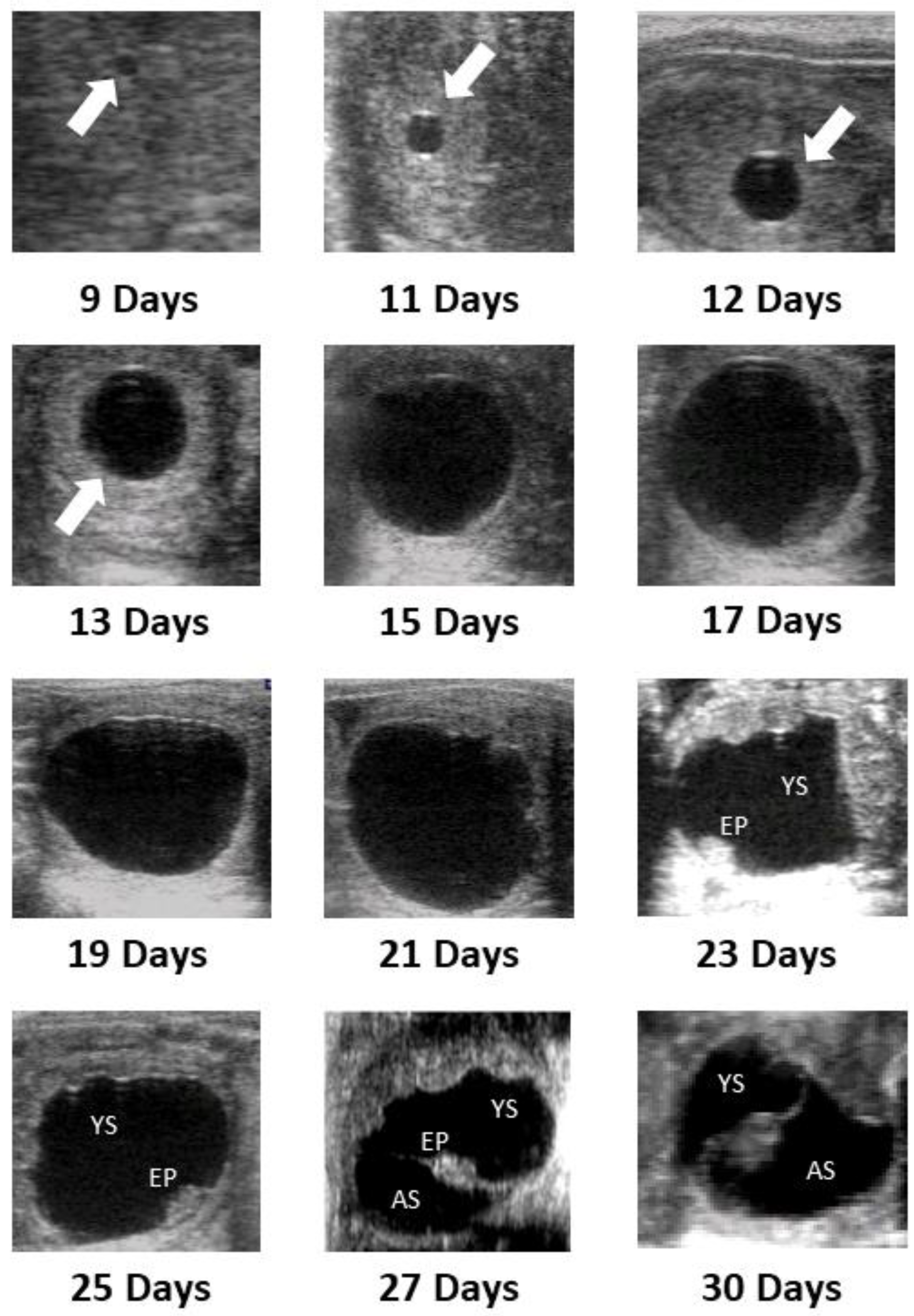
3.2. Uterine Dynamics and Embryo Development
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arfuso, F.; Quartuccio, M.; Bazzano, M.; Fazio, F.; Piccione, G. Erythrocyte osmotic fragility and select hematologic variables in postparturient mares and their foals. Vet. Clin. Pathol. 2016, 45, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Arfuso, F.; Giannetto, C.; Rizzo, M.; Fazio, F.; Giudice, E.; Piccione, G. Theriogenology Serum levels of mitochondrial uncoupling protein 1, leptin, and lipids during late pregnancy and the early postpartum period in mares. Theriogenology 2016, 86, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Piccione, G.; Rizzo, M.; Arfuso, F.; Giannetto, C.; Di Pietro, S.; Bazzano, M.; Quartuccio, M. Leukocyte modifications during the first month after foaling in mares and their newborn foals. Pol. J. Vet. Sci. 2015, 18, 621–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piccione, G.; Arfuso, F.; Abbate, F.; Giannetto, C.; Panzera, M.; Rizzo, M.; Fazio, F. Adrenocorticotrophic hormone and cortisol levels during late pregnancy and post-foaling period in mares. Anim. Sci. Pap. Rep. 2017, 35, 173–180. [Google Scholar]
- Arfuso, F.; Giannetto, C.; Bazzano, M.; Assenza, A.; Piccione, G. Physiological correlation between hypothalamic–pituitary–adrenal axis, leptin, UCP1 and lipid panel in mares during late pregnancy and early postpartum period. Animals 2021, 11, 2051. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.; Ferreira, J.C.P.; Papa, F.O.; Henry, M. Ovarian activity and plasma concentrations of progesterone and estradiol during pregnancy in jennies. Theriogenology 1998, 49, 1465–1473. [Google Scholar] [CrossRef]
- Meira, C.; Ferreira, J.C.P.; Papa, F.O.; Henry, M. Ultrasonographic evaluation of the conceptus from days 10 to 60 of pregnancy in jennies. Theriogenology 1998, 49, 1475–1482. [Google Scholar] [CrossRef]
- Crisci, A.; Rota, A.; Panzani, D.; Sgorbini, M.; Ousey, J.C.; Camillo, F. Clinical, ultrasonographic, and endocrinological studies on donkey pregnancy. Theriogenology 2014, 81, 275–283. [Google Scholar] [CrossRef]
- Nervo, T.; Bertero, A.; Poletto, M.; Pregel, P.; Leone, R.; Toffoli, V.; Vincenti, L. Field ultrasound evaluation of some gestational parameters in jennies. Theriogenology 2019, 126, 95–105. [Google Scholar] [CrossRef]
- Bergfelt, D.R.; Adams, G.P. Pregnancy. In Equine Reproduction; McKinnon, A.O., Squires, E.L., Vaala, W.E., Varner, D.D., Eds.; Wiley-Blackwell: Oxford, UK, 2011; pp. 2065–2079. [Google Scholar]
- Palmer, E.; Driancourt, M.A. Use of ultrasonic echography in equine gynecology. Theriogenology 1980, 13, 203–216. [Google Scholar] [CrossRef]
- Castro, T.; Jacob, J.C.; Stefani, G.; Domingues, R.R.; Ginther, O.J. Concentrations of progesterone and a PGF2α metabolite during the interovulatory interval compared to the corresponding days of pregnancy in mares. Theriogenology 2021, 165, 10–17. [Google Scholar] [CrossRef]
- McKinnon, A.O.; Squires, E.L.; Vaala, W.E.; Varner, D.D. Physiology and endocrinology. In Equine Reproduction; McKinnon, A.O., Squires, E.L., Vaala, W.E., Varner, D.D., Eds.; Willey-Blackwell: Oxford, UK, 2011; pp. 1601–1688. [Google Scholar]
- Canisso, I.F.; Panzani, D.; Miró, J.; Ellerbrock, R.E. Key Aspects of Donkey and Mule Reproduction. Vet. Clin. N. Am. Equine Pract. 2019, 35, 607–642. [Google Scholar] [CrossRef]
- Squires, E.L.; Ginther, O.J. Collection Technique and Progesterone Concentration of Ovarian and Uterine Venous Blood in Mares. J. Anim. Sci. 1975, 40, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urwin, V.E.; Allen, W.R. Pituitary and chorionic gonadotrophic control of ovarian function during early pregnancy in equids. J. Reprod. Fertil. Suppl. 1982, 32, 371–381. [Google Scholar]
- Ginther, O.J. Embryonic loss in mares: Nature of loss after experimental induction by ovariectomy or prostaglandin F2α. Theriogenology 1985, 24, 87–98. [Google Scholar] [CrossRef]
- Camillo, F.; Rota, A.; Biagini, L.; Tesi, M.; Fanelli, D.; Panzani, D. The current situation and trend of donkey industry in Europe. J. Equine Vet. Sci. 2018, 65, 44–49. [Google Scholar] [CrossRef]
- Carneiro, G.F.; Cavalcante Lucena, J.E.; de Oliveira Barros, L. The current situation and trend of the donkey industry in South America. J. Equine Vet. Sci. 2018, 65, 106–110. [Google Scholar] [CrossRef]
- Canisso, I.F.; Davies Morel, M.C.G.; McDonnell, S. Strategies for the management of donkey jacks in intensive breeding systems. Equine Vet. Educ. 2009, 21, 652–659. [Google Scholar] [CrossRef] [Green Version]
- Miragaya, M.H.; Neild, D.M.; Alonso, A.E. A review of reproductive biology and biotechnologies in donkeys. J. Equine Vet. Sci. 2018, 65, 55–61. [Google Scholar] [CrossRef]
- Mccue, P.; Scoggin, C.; Lindholm, A. Estrus. In Equine Reproduction; Wiley: Hoboken, NJ, USA, 2011; Volume 1, pp. 1716–1727. ISBN 9788578110796. [Google Scholar]
- Penitente-Filho, J.M.; Jimenez, C.R.; Zolini, A.M.; Carrascal, E.; Azevedo, J.L.; Silveira, C.O.; Oliveira, F.A.; Torres, C.A.A. Influence of corpus luteum and ovarian volume on the number and quality of bovine oocytes. Anim. Sci. J. 2015, 86, 148–152. [Google Scholar] [CrossRef]
- Ginther, O.J. Intrauterine movement of the early conceptus in barren and postpartum mares. Theriogenology 1984, 21, 633–644. [Google Scholar] [CrossRef]
- Kollipara, S.; Bende, G.; Agarwal, N.; Varshney, B.; Paliwal, J. International guidelines for bioanalytical method validation: A comparison and discussion on current scenario. Chromatographia 2011, 73, 201–217. [Google Scholar] [CrossRef]
- Canisso, I.F.; Gallacher, K.; Gilbert, M.A.; Korn, A.; Schweizer, C.M.; Bedford-Guaus, S.J.; Gilbert, R.O. Preovulatory progestagen treatment in mares fails to delay ovulation. Vet. J. 2013, 197, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Cuervo-Arango, J.; Newcombe, J.R. Repeatability of preovulatory follicular diameter and uterine edema pattern in two consecutive cycles in the mare and how they are influenced by ovulation inductors. Theriogenology 2008, 69, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Segabinazzi, L.G.T.M.; Oba, E.; Alvarenga, M.A. The combination of hCG and GnRH analog to hasten ovulation in mares does not change luteal function and pregnancy outcome in embryo recipient mares. J. Equine Vet. Sci. 2021, 105, 103691. [Google Scholar] [CrossRef]
- Ishak, G.M.; Bashir, S.T.; Gastal, M.O.; Gastal, E.L. Pre-ovulatory follicle affects corpus luteum diameter, blood flow, and progesterone production in mares. Anim. Reprod. Sci. 2017, 187, 1–12. [Google Scholar] [CrossRef]
- Díaz-Duran, M.; Zarco, L.; Boeta, A.M. Ovarian dynamics and estrous cycle length in the donkey (Equus asinus). Theriogenology 2017, 103, 1–8. [Google Scholar] [CrossRef]
- Squires, E.L.; Wentworth, B.C.; Ginther, O.J. Progesterone concentration in blood of mares during the estrous cycle, pregnancy and after hysterectomy. J. Anim. Sci. 1974, 39, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Utt, M.D.; Bergfelt, D.R.; Beg, M.A. Controlling interrelationships of progesterone/LH and estradiol/LH in mares. Anim. Reprod. Sci. 2006, 95, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.; Ferreira, J.C.P.; Papa, F.O.; Tornero, M.T.T.; Bicudo, S.D. Study of the estrous cycle in donkeys (Equus asinus) using ultrasonography and plasma progesterone concentrations. Biol. Reprod. 1995, 52, 403–410. [Google Scholar] [CrossRef]
- Ferreira, J.C.; Filho, L.F.N.; Boakari, Y.L.; Canesin, H.S.; Thompson, D.L.; Lima, F.S.; Meira, C. Hemodynamics of the corpus luteum in mares during experimentally impaired luteogenesis and partial luteolysis. Theriogenology 2018, 107, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ginther, O.J.; Gastal, E.L.; Gastal, M.O.; Utt, M.D.; Beg, M.A. Luteal blood flow and progesterone production in mares. Anim. Reprod. Sci. 2007, 99, 213–220. [Google Scholar] [CrossRef]
- Ginther, O.J.; Rodrigues, B.L.; Ferreira, J.C.; Araujo, R.R.; Beg, M.A. Characterisation of pulses of 13,14-dihydro-15-keto-PGF2alpha (PGFM) and relationships between PGFM pulses and luteal blood flow before, during, and after luteolysis in mares. Reprod. Fertil. Dev. 2008, 20, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J. A 40-year odyssey into the mysteries of equine luteolysis. Theriogenology 2009, 72, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Allen, W.E.; Hadley, J.C. Blood progesterone concentrations in pregnant and non-pregnant mares. Equine Vet. J. 1974, 6, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Sevinga, M.; Schukken, Y.H.; Hesselink, J.W.; Jonker, F.H. Relationship between ultrasonic characteristics of the corpus luteum, plasma progesterone concentration and early pregnancy diagnosis in Friesian mares. Theriogenology 1999, 52, 585–592. [Google Scholar] [CrossRef]
- Richard, H.; Appleton, P.; Kesler, D.J.; Ferguson, C.E. Progesterone supplementation day 3 to 5 post-ovulation in ai-bred mares. Reprod. Fertil. Dev. 2013, 25, 153. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, B.A.; Daels, P.F. Immunolocalization of 3 beta-hydroxysteroid dehydrogenase, cytochrome P450 17 alpha-hydroxylase/17,20-lyase and cytochrome P450 aromatase in the equine corpus luteum of dioestrus and early pregnancy. J. Reprod. Fertil. 1997, 111, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, B.A.; MacLeod, J.N.; Daels, P.F. Differential transcription of steroidogenic enzymes in the equine primary corpus luteum during diestrus and early pregnancy. Biol. Reprod. 1997, 56, 821–829. [Google Scholar] [CrossRef] [Green Version]
- Watson, E.D.; Thomson, S.R.M.; Howie, A.F. Detection of steroidogenic acute regulatory protein in equine ovaries. J. Reprod. Fertil. 2000, 119, 187–192. [Google Scholar] [CrossRef]
- Klein, C.; Troedsson, M.H.T. Maternal recognition of pregnancy in the horse: A mystery still to be solved. Reprod. Fertil. Dev. 2011, 23, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Troedsson, M.H.T.; Ababneh, M.M.; Ohlgren, A.F.; Madill, S.; Vetscher, N.; Gregas, M. Effect of periovulatory prostaglandin F2α on pregnancy rates and luteal function in the mare. Theriogenology 2001, 55, 1891–1899. [Google Scholar] [CrossRef]
- Kimball, F.A.; Wyngarden, L.J. Prostaglandin F2α specific binding in equine corpora lutea. Prostaglandins 1977, 13, 553–564. [Google Scholar] [CrossRef]
- Flood, P.F.; Jong, A.; Betteridge, K.J. The location of eggs retained in the oviducts of mares. J. Reprod. Fertil. 1979, 57, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Onuma, H.; Ohnami, Y. Retention of tubal eggs in mares. J. Reprod. Fertil. 1975, 23 (Suppl. 1), 507–511. [Google Scholar]
- Freeman, D.A.; Weber, J.A.; Geary, R.T.; Woods, G.L. Time of embryo transport through the mare oviduct. Theriogenology 1991, 36, 823–830. [Google Scholar] [CrossRef]
- Weber, A.J.; Freeman, D.A.; Vanderwall, D.K.; Woods, G.L. Prostaglandin E2 Hastens Oviductal Transport of Equine Embryos. Biol. Reprod. 1991, 45, 544–546. [Google Scholar] [CrossRef]
- Kowalewski, M.P.; Fox, B.; Gram, A.; Boos, A.; Reichler, I. Prostaglandin E2 functions as a luteotrophic factor in the dog. Reproduction 2013, 145, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Shelton, K.; Parkinson, T.J.; Hunter, M.G.; Kelly, R.W.; Lamming, G.E. Prostaglandin E-2 as a potential luteotrophic agent during early pregnancy in cattle. J. Reprod. Fertil. 1990, 90, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCue, P.; Vanderwall, D.; Keith, S.; Squires, E. Equine embryo transfer: Influence of endogenous progesterone concentration in recipients on pregnancy outcome. Theriogenology 1999, 51, 267. [Google Scholar] [CrossRef]
- Bergfelt, D.R.; Woods, J.A.; Ginther, O.J. Role of the embryonic loss in mares. J. Reprod. Fertil. 1992, 95, 339–347. [Google Scholar] [CrossRef] [Green Version]
- Canisso, I.F.; Beltaire, K.A.; Bedford-Guaus, S.J. Premature luteal regression in a pregnant mare and subsequent pregnancy maintenance with the use of oral altrenogest. Equine Vet. J. 2012, 45, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Garcia, M.C.; Bergfelt, D.R.; Leith, G.S.; Scraba, S.T. Embryonic loss in mares: Pregnancy rate, length of interovulatory intervals, and progesterone concentrations associated with loss during days 11 to 15. Theriogenology 1985, 24, 409–417. [Google Scholar] [CrossRef]
- Klein, C.; Scoggin, K.E.; Ealy, A.D.; Troedsson, M.H.T. Transcriptional profiling of equine endometrium during the time of maternal recognition of pregnancy. Biol. Reprod. 2010, 83, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Merkl, M.; Ulbrich, S.E.; Otzdorff, C.; Herbach, N.; Wanke, R.; Wolf, E.; Handler, J.; Bauersachs, S. Microarray analysis of equine endometrium at days 8 and 12 of pregnancy. Biol. Reprod. 2010, 83, 874–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonergan, P.; Forde, N.; Spencer, T. Role of progesterone in embryo development in cattle. Reprod. Fertil. Dev. 2016, 28, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, E.M.; Ramirez, R.J.; Squires, E.L.; Alvarenga, M.A.; Vanderwall, D.K.; McCue, P.M. Factors affecting pregnancy rates and early embryonic death after equine embryo transfer. Theriogenology 2000, 54, 965–979. [Google Scholar] [CrossRef]
- Bessent, C.; Ginther, O.J. Comparison of early conceptus mobility between mares and jennies. Theriogenology 1988, 29, 913–920. [Google Scholar] [CrossRef]
- Ginther, O.J. Mobility of the early equine conceptus. Theriogenology 1983, 19, 603–611. [Google Scholar] [CrossRef]
- Stout, T.A.E.; Allen, W.R. Role of prostaglandins in intrauterine migration of the equine conceptus. Reproduction 2001, 121, 771–775. [Google Scholar] [CrossRef]
- Gastal, E.L.; Santos, G.F.; Henry, M.; Piedade, H.M. Embryonic and early foetal development in donkeys. Equine Vet. J. 1993, 15, 10–13. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day From First Detection | ||
|---|---|---|
| Mean ± SEM | Range | |
| Embryonic vesicle | 9.25 ± 0.2 | 9–10 |
| Fixation | 15 ± 0.3 | 13–16 |
| Loss of spherical shape | 18.6 ± 0.4 | 16–20 |
| Embryo proper | 20.8 ± 0.4 | 19–21 |
| Embryonic heartbeat | 22 ± 0.3 | 21–23 |
| Allantoic sac | 24.4 ± 0.3 | 23–26 |
| Day of Pregnancy | Total Moves | How Many Segments the EV Crossed (Distance Traveled) | Number of Horn Changes | Percentage of Time in the Body |
|---|---|---|---|---|
| 9 (n = 5) | 3.6 ± 2.0 | 4.2 ± 2.6 | 0.8 ± 0.8 | 58.6% |
| 10 (n = 7) | 3.6 ± 2.6 | 3.7 ± 2.9 | 0.3 ± 0.7 | 65.6% |
| 11 (n = 8) | 5.5 ± 2.6 | 6.5 ± 2.8 | 1.1 ± 1.1 | 35.0% |
| 12 (n = 8) | 5.0 ± 0.9 | 5.8 ± 1.1 | 1.1 ± 0.8 | 41.0% |
| 13 (n = 8) | 4.8 ± 2.3 | 5.8 ± 3.0 | 1.3 ± 1.1 | 37.0% |
| 14 (n = 7) | 5.0 ± 1.8 | 5.6 ± 2.2 | 0.9 ± 0.8 | 19.9% |
| 15 (n = 7) | 0.6 ± 0.9 | 0.6 ± 0.9 | 0.1 ± 0.3 | 2.1% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Segabinazzi, L.G.T.M.; Roberts, B.N.; Peterson, E.W.; Ambrosia, R.; Bergfelt, D.; Samper, J.; French, H.; Gilbert, R.O. Early Pregnancy in Jennies in the Caribbean: Corpus Luteum Development and Progesterone Production, Uterine and Embryo Dynamics, Conceptus Growth and Maturation. Animals 2022, 12, 127. https://doi.org/10.3390/ani12020127
Segabinazzi LGTM, Roberts BN, Peterson EW, Ambrosia R, Bergfelt D, Samper J, French H, Gilbert RO. Early Pregnancy in Jennies in the Caribbean: Corpus Luteum Development and Progesterone Production, Uterine and Embryo Dynamics, Conceptus Growth and Maturation. Animals. 2022; 12(2):127. https://doi.org/10.3390/ani12020127
Chicago/Turabian StyleSegabinazzi, Lorenzo G. T. M., Brandy N. Roberts, Erik W. Peterson, Rachael Ambrosia, Don Bergfelt, Juan Samper, Hilari French, and Robert O. Gilbert. 2022. "Early Pregnancy in Jennies in the Caribbean: Corpus Luteum Development and Progesterone Production, Uterine and Embryo Dynamics, Conceptus Growth and Maturation" Animals 12, no. 2: 127. https://doi.org/10.3390/ani12020127