
Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
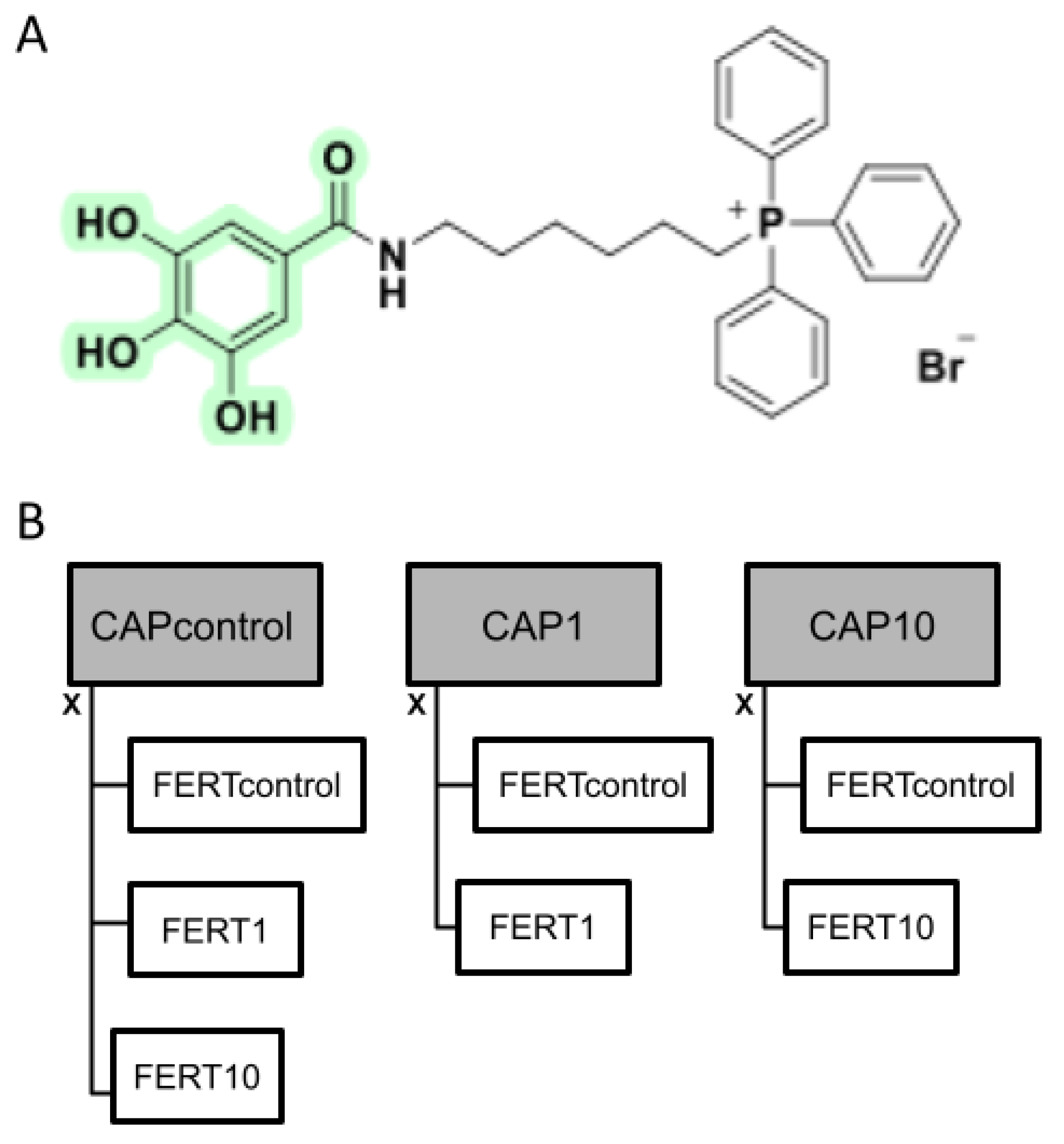
2.2. Synthesis of AntiOxBEN2
2.3. Experimental Design
2.4. Oocyte Collection and In Vitro Maturation
2.5. Sperm Capacitation and Evaluation
2.5.1. Sperm Motility Assessment
2.5.2. Sperm Vitality Assessment
2.5.3. Mitochondrial Membrane Potential
2.5.4. Sperm Intracellular Oxidative Stress
2.5.5. Cell Mass
2.5.6. Intracellular ATP Levels
2.5.7. Cellular Oxygen Consumption Rate
2.6. Oocyte In Vitro Fertilization and Embryo Development
2.7. Statistical Analysis
3. Results
3.1. AntiOxBEN2 Dose-Dependently Decreased Sperm Motility
3.2. AntiOxBEN2 Did Not Alter Semen Vitality nor Induced Sperm Abnormalities
3.3. AntiOxBEN2 Improved Sperm Mitochondrial Membrane Potential
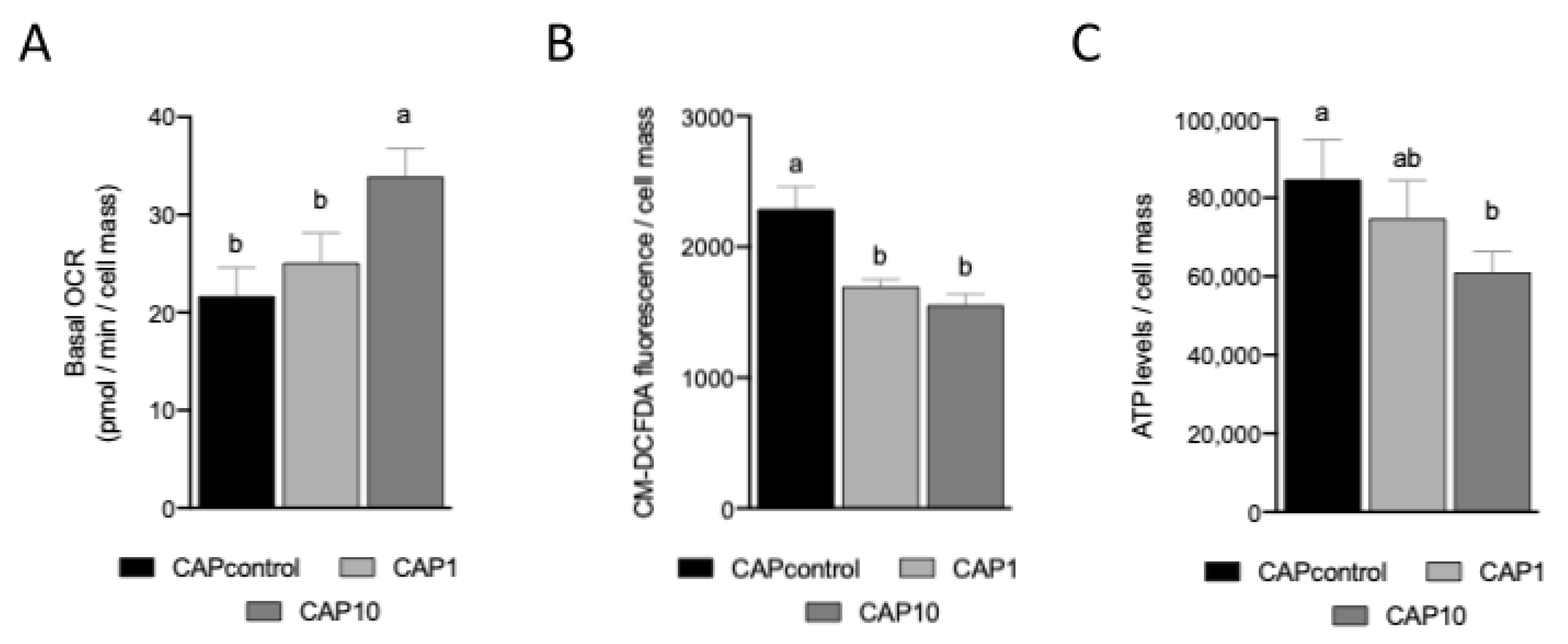
3.4. AntiOxBEN2 Dose-Dependently Increased Sperm Cellular (Basal) Oxygen Consumption
3.5. AntiOxBEN2 Decreased Sperm Cellular Oxidative Stress
3.6. AntiOxBEN2 Dose-Dependently Decreased Sperm Intracellular ATP Levels
3.7. AntiOxBEN2 Dose-Dependently Increased the Number of Produced Embryos
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aitken, R.J. Impact of oxidative stress on male and female germ cells: Implications for fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R. Reactive Oxygen Species as the Molecular Modulators of Calcium Oxalate Kidney Stone Formation: Evidence from Clinical and Experimental Investigations. Urol. J. 2013, 189, 803–811. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, J.; Oliveira, C.; Amorim, R.; Cagide, F.; Garrido, J.; Ribeiro, J.A.; Pereira., C.M.; Silva., A.F.; Andrade, P.B.; Oliveira, P.J.; et al. Development of hydroxybenzoic-based platforms as a solution to deliver dietary antioxidants to mitochondria. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef]
- Trevizan, J.T.; Carreira, J.T.; Carvalho, I.R.; Kipper, B.H.; Nagata, W.B.; Perri, S.H.V.; Franco Oliveira, M.E.; Pierucci, J.C.; Koivisto, M.B. Does lipid peroxidation and oxidative DNA damage differ in cryopreserved semen samples from young, adult and aged Nellore bulls? Anim. Reprod. Sci. 2018, 195, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and Male Fertility: From Molecular Studies to Clinical Evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Wyck, S.; Herrera, C.; Requena, C.E.; Bittner, L.; Hajkova, P.; Bollwein, H.; Santoro, R. Oxidative stress in sperm affects the epigenetic reprogramming in early embryonic development. Epigenetics Chromatin 2018, 11, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panner Selvam, M.K.; Agarwal, A.; Henkel, R.; Finelli, R.; Robert, K.; Iovine, C.; Baskaran, S. The effect of oxidative and reductive stress on semen parameters and functions of physiologically normal human spermatozoa. Free Radic. Biol. Med. 2020, 152, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Baker, M.A. Oxidative stress, sperm survival and fertility control. Mol. Cell. Endocrinol. 2006, 250, 66–69. [Google Scholar] [CrossRef]
- De Assis, P.M.; Castro, L.S.; Siqueira, A.F.P.; De Carvalho Delgado, J.; Dos Santos Hamilton, T.R.; Goissis, M.D.; Mendes, C.M.; Nichi, M.; Visintin, J.A.; Assumpção, M.E. System for evaluation of oxidative stress on in-vitro-produced bovine embryos. Reprod. BioMed. Online 2015, 31, 577–580. [Google Scholar] [CrossRef] [Green Version]
- Bermejo-Álvarez, P.; Lonergan, P.; Rizos, D.; Gutiérrez-Adan, A. Low oxygen tension during IVM improves bovine oocyte competence and enhances anaerobic glycolysis. Reprod. BioMed. Online 2010, 20, 341–349. [Google Scholar] [CrossRef] [Green Version]
- Kasai, T.; Ogawa, K.; Mizuno, K.; Nagai, S.; Uchida, Y.; Ohta, S.; Fujie, M.; Suzuki, K.; Hirata, S.; Hoshi, K. Relationship between sperm mitochondrial membrane potential, sperm motility, and fertility potential. Asian J. Androl. 2002, 4, 97–103. [Google Scholar] [PubMed]
- Leite, R.F.; Annes, K.; Ispada, J.; de Lima, C.B.; Dos Santos, É.C.; Fontes, P.K.; Nogueira, M.F.G.; Milazzotto, M.P. Oxidative stress alters the profile of transcription factors related to early development on in vitro produced embryos. Oxid. Med. Cell. Longev. 2017, 2017, 1502489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.A.J.; Hartley, R.C.; Cochemé, H.M.; Murphy, M.P. Mitochondrial pharmacology. Trends Pharmacol. Sci. 2012, 33, 341–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, J.; Deus, C.M.; Borges, F.; Oliveira, P.J. Mitochondria: Targeting mitochondrial reactive oxygen species with mitochondriotropic polyphenolic-based antioxidants. Int. J. Biochem. Cell Biol. 2018, 97, 98–103. [Google Scholar] [CrossRef]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.R.P.; Ferreira, I.C.F.R. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [Green Version]
- Pezzini, I.; Mattoli, V.; Ciofani, G. Mitochondria and neurodegenerative diseases: The promising role of nanotechnology in targeted drug delivery. Expert Opin. Drug Deliv. 2017, 14, 513–523. [Google Scholar] [CrossRef]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The Neuroprotective Effects of Phenolic Acids: Molecular Mechanism of Action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, C.; Cagide, F.; Teixeira, J.; Amorim, R.; Sequeira, L.; Mesiti, F.; Silva, T.; Garrido, J.; Remião, F.; Vilar, S.; et al. Hydroxybenzoic Acid Derivatives as Dual-Target Ligands: Mitochondriotropic Antioxidants and Cholinesterase Inhibitors. Front. Chem. 2018, 6, 126. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Cagide, F.; Borges, F.; Montezinho, L.; Oliveira, P.; Pereira, R.M.L.N. Effect of a new mitochondriotropic antioxidant on oocyte maturation and embryo production. In Book of Abstracts of the 71st Annual Meeting of the European Federation of Animal Science Virtual Meeting–1 to 4 December; Scientific Committee, Ed.; Wageningen academic publishers: Wageningen, The Netherland, 2020; p. 672. [Google Scholar] [CrossRef]
- Lapa, M.; Marques, C.C.; Alves, S.P.; Vasques, M.I.; Baptista, M.C.; Carvalhais, I.; Silva Pereira, M.; Horta, A.E.M.; Bessa, R.J.B.; Pereira, R.M. Effect of trans-10 cis-12 conjugated linoleic acid on Bovine Oocyte Competence and Fatty Acid Composition. Reprod. Dom. Anim. 2011, 46, 904–910. [Google Scholar] [CrossRef]
- Fonseca, E.; Mesquita, P.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Matos, J.E.; Soveral, G.; Pereira, R.M.L.N. Modulation of P2Y2 receptors in bovine cumulus oocyte complexes: Effects on intracellular calcium, zona hardening and developmental competence. Purinergic Signal. 2020, 16, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.G.; Hasler, J.F. A 100-Year Review: Reproductive technologies in dairy science. J. Dairy Sci. 2017, 100, 10314–10331. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.M.; Marques, C.C.; Baptista, M.C.; Vasques, M.I.; Horta, A.E. Effect of arachidonic acid supplementation and cyclooxygenase/lipoxygenase inhibition on the development of early bovine embryos. Rev. Bras. Zootec. 2006, 35, 422–427. [Google Scholar] [CrossRef] [Green Version]
- Pereira, R.M.; Mesquita, P.; Pires, V.; Baptista, M.C.; Barbas, J.P.; Pimenta, J.; Horta, A.E.M.; Prates, J.A.M.; Marques, C.C. Prion protein testis specific (PRNT) gene polymorphisms and transcript level in ovine spermatozoa: Implications in freezability, fertilization and embryo production. Theriogenology 2018, 115, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Gamboa, S.; Rodrigues, A.S.; Henriques, L.; Batista, C.; Ramalho-Santos, J. Seasonal functional relevance of sperm characteristics in equine spermatozoa. Theriogenology 2010, 73, 950–958. [Google Scholar] [CrossRef]
- Vichai, V.; Kirtikara, K. Sulforhodamine B colorimetric assay for cytotoxicity screening. Nat. Protoc. 2006, 1, 1112–1116. [Google Scholar] [CrossRef]
- Deus, C.M.; Zehowski, C.; Nordgren, K.; Wallace, K.B.; Skildum, A.; Oliveira, P.J. Stimulating basal mitochondrial respiration decreases doxorubicin apoptotic signaling in H9c2 cardiomyoblasts. Toxicology 2015, 334, 1–11. [Google Scholar] [CrossRef]
- Pereira, R.M.; Marques, C.C.; Baptista, M.C.; Vasques, M.I.; Horta, A.E.M. Embryos and culture cells: A model for studying the effect of progesterone. Anim. Reprod. Sci. 2009, 111, 31–40. [Google Scholar] [CrossRef]
- Stringfellow, D.A.; Seidel, S.M. (Eds.) Manual of the International Embryo Transfer Society; International Embryo Transfer Society, INC: Savoy, IL, USA, 1998. [Google Scholar]
- Attia, S.; Katila, T.; Andersson, M. The Effect of Sperm Morphology and Sire Fertility on Calving Rate of Finnish Ayrshire AI Bulls. Reprod. Dom. Anim. 2016, 51, 54–58. [Google Scholar] [CrossRef]
- Darr, C.R.; Cortopassi, G.A.; Datta, S.; Varner, D.D.; Meyers, S.A. Mitochondrial oxygen consumption is a unique indicator of stallion spermatozoal health and varies with cryopreservation media. Theriogenology 2016, 86, 1382–1392. [Google Scholar] [CrossRef]
- Ford, W.C.L. Regulation of sperm function by reactive oxygen species. Hum. Reprod. Update 2004, 10, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Garret, L.J.A.; Revell, S.G.; Leese, H.J. Adenosine Triphosphate Production by Bovine Spermatozoa and Its Relationship to Semen Fertilizing Ability. J. Androl. 2008, 29, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Magdanz, V.; Boryshpolets, S.; Ridzewski, C.; Eckel, B.; Reinhardt, K. The motility-based swim-up technique separates bull sperm based on differences in metabolic rates and tail length. PLoS ONE 2019, 14, e0223576. [Google Scholar] [CrossRef]
- Zarbakhsh, S. Effect of antioxidants on preimplantation embryo development in vitro: A review. Zygote 2021, 14, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Anchordoquy, J.P.; Lizarraga, R.M.; Anchordoquy, J.M.; Nikoloff, N.; Rosa, D.E.; Fabra, M.C.; Peral-García, P.; Furnus, C.C. Effect of cysteine, glutamate and glycine supplementation to in vitro fertilization medium during bovine early embryo development. Reprod. Biol. 2019, 19, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Santana, P.P.B.; da Silva, B.B.; Silva, T.V.G.; Costa, N.N.; Cordeiro, M.S.; Santos, S.S.D.; Ohashi, O.M.; Miranda, M.S. Addition of L-arginine to the fertilization medium enhances subsequent bovine embryo development rates. Theriogenology 2016, 85, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Sapanidou, V.; Taitzoglou, I.; Tsakmakidis, I.; Kourtzelis, I.; Fletouris, D.; Theodoridis, A.; Lavrentiadou, S.; Tsantarliotou, M. Protective effect of crocetin on bovine spermatozoa against oxidative stress during in vitro fertilization. Andrology 2016, 4, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Tirpák, F.; Slanina, T.; Kováčik, A.; Ondruška, L.; Massányi, P.; Halo, M.; Massányi, P. Low taurine concentrations possitively affect rabbit spermatozoa properties in later time intervals. J. Microbiol. Biotechnol. Food Sci. 2017, 7, 128–131. [Google Scholar] [CrossRef] [Green Version]
- Marei, W.F.A.; Van Den Bosch, L.; Pintelon, I.; Mohey-Elsaeed, O.; Bols, P.E.J.; Leroy, J.L.M.R. Mitochondria-targeted therapy rescues development and quality of embryos derived from oocytes matured under oxidative stress conditions: A bovine in vitro model. Hum. Reprod. 2019, 34, 1984–1998. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Garner, D.L.; Thomas, C.A. Organelle-specific probe JC-1 identifies membrane potential differences in the mitochondrial function of bovine sperm. Mol. Reprod. Dev. 1999, 53, 222–229. [Google Scholar] [CrossRef]
- Amaral, A.; Ramalho-Santos, J. Assessment of mitochondrial potential: Implications for the correct monitoring of human sperm function. Int. J. Androl. 2010, 33, e180–e186. [Google Scholar] [CrossRef] [PubMed]
- Nichi, M.; Rijsselaere, T.; Losano, J.; Angrimani, D.; Kawai, G.; Goovaerts, I.; Van Soom, A.; Barnabe, V.; De Clercq, J.; Bols, P. Evaluation of epididymis storage temperature and cryopreservation conditions for improved mitochondrial membrane potential, membrane integrity, sperm motility and in vitro fertilization in bovine epididymal sperm. Reprod. Dom. Anim. 2017, 52, 257–263. [Google Scholar] [CrossRef]
- Fraser, L.R. The “switching on” of mammalian spermatozoa: Molecular events involved in promotion and regulation of capacitation. Mol. Reprod. Dev. 2010, 77, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Krishnakumar, S.; Whiteside, D.; Elkin, B.; Thundathil, J. Effect of Reproductive Seasonality on Gamete Quality in the North American Bison ( Bison bison bison ). Reprod. Domest. Anim. 2015, 50, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, S.; Matoba, S.; Hashiyada, Y.; Aikawa, Y.; Ohtake, M.; Matsuda, H.; Kobayashi, S.; Konishi, K.; Imai, K. Oxidative phosphorylation-linked respiration in individual bovine oocytes. J. Reprod. Dev. 2012, 58, 636–641. [Google Scholar] [CrossRef] [Green Version]
- Marques, C.C.; Santos-Silva, C.; Rodrigues, C.; Matos, J.E.; Moura, T.; Baptista, M.C.; Horta, A.E.M.; Bessa, R.J.B.; Alves, S.P.; Soveral, G.; et al. Bovine oocyte membrane permeability and cryosurvival: Effects of different cryoprotectants and calcium in the vitrification media. Cryobiology 2018, 81, 4–11. [Google Scholar] [CrossRef]
- Reily, C.; Mitchell, T.; Chacko, B.K.; Benavides, G.; Murphy, M.P.; Darley-Usmar, V. Mitochondrially targeted compounds and their impact on cellular bioenergetics. Redox Biol. 2013, 1, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Valera-Alberni, M.; Canto, C. Mitochondrial stress management: A dynamic journey. Cell Stress 2018, 2, 253–274. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinematic Parameters | Groups | ||
|---|---|---|---|
| CAPcontrol | CAP1 | CAP10 | |
| Total Motility (%) | 79.6 ± 7.03 | 75.8 ± 7.03 | 69.8 ± 7.03 |
| Progressive Motility (%) | 42.8 ± 5.85 a | 37.9 ± 5.85 ab | 28.5 ± 5.85 b |
| Visual Motility (%) | 56.5 ± 5.16 a | 50.7 ± 5.16 a | 44.0 ± 5.16 b |
| VCL (µms−1) | 114.6 ± 14.4 | 99.5 ± 14.4 | 97.5 ± 14.4 |
| VAP (µms−1) | 59.1 ± 7.02 | 51.6 ± 7.02 | 50.5 ± 7.02 |
| Slow motile spz (n) | 4.9 ± 2.47 | 8.3 ± 2.47 | 11.8 ± 2.47 |
| Medium motile spz (n) | 20.9 ± 6.05 | 26.4 ± 6.05 | 25.1 ± 6.05 |
| Rapid motile spz (n) | 145.1 ± 35.16 | 136.4 ± 35.16 | 92.1 ± 35.16 |
| VSL (µms−1) | 45.2 ± 6.21 | 37.4 ± 6.21 | 35.6 ± 6.21 |
| ALH (µm) | 2.8 ± 0.30 | 2.5 ± 0.30 | 2.6 ± 0.30 |
| Linearity | 39.1 ± 1.95 | 37.0 ± 1.95 | 37.3 ± 1.95 |
| Straightness | 75.4 ± 2.72 | 71.6 ± 2.72 | 70.8 ± 2.72 |
| Wobble VAP/VCL | 51.8 ± 1.23 | 51.5 ± 1.23 | 52.5 ± 1.23 |
| BCF (Hz) | 13.5 ± 1.18 | 13.3 ± 1.18 | 11.9 ± 1.18 |
| Kinematic Parameters | Groups | ||
|---|---|---|---|
| CAPcontrol | CAP1 | CAP10 | |
| Vitality (%) | 47.3 ± 6.96 | 44.9 ± 6.96 | 45.8 ± 6.96 |
| Normal spz (%) | 79.8 ± 1.65 | 81.2 ± 1.65 | 80.5 ± 1.65 |
| Head defect (%) | 7.3 ± 0.84 | 7.4 ± 0.84 | 9.1 ± 0.84 |
| Int.piece defect (%) | 6.8 ± 1.17 | 5.8 ± 1.17 | 5.1 ± 1.17 |
| Tail defect (%) | 6.7 ± 0.88 | 6.2 ± 0.88 | 5.8 ± 0.88 |
| Groups | Inseminated Oocytes (n) | Cleavage (%) | D7 Embryos (n) | D7 Embryos (%) |
|---|---|---|---|---|
| CAPcontrol × FERTcontrol | 306 | 61.8 ± 2.84 bc | 27 | 15.9 ± 2.85 |
| CAPcontrol × FERT1 | 266 | 69.4 ± 2.86 ab | 29 | 18.4 ± 3.12 |
| CAPcontrol × FERT10 | 243 | 57.2 ± 3.22 c | 13 | 10.9 ± 2.87 |
| CAP1 × FERTcontrol | 248 | 61.4 ± 3.14 bc | 25 | 17.7 ± 3.24 |
| CAP1 × FERT1 | 264 | 72.7 ± 2.77 a | 37 | 21.6 ± 3.16 |
| CAP10 × FERTcontrol | 293 | 64.4 ± 2.85 bc | 20 | 12.2 ± 2.58 |
| CAP10 × FERT10 | 244 | 66.9 ± 3.06 ab | 16 | 11.6 ± 2.66 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, J.C.; Marques, C.C.; Baptista, M.C.; Pimenta, J.; Teixeira, J.; Montezinho, L.; Cagide, F.; Borges, F.; Oliveira, P.J.; Pereira, R.M.L.N. Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production. Animals 2022, 12, 804. https://doi.org/10.3390/ani12070804
Santos JC, Marques CC, Baptista MC, Pimenta J, Teixeira J, Montezinho L, Cagide F, Borges F, Oliveira PJ, Pereira RMLN. Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production. Animals. 2022; 12(7):804. https://doi.org/10.3390/ani12070804
Chicago/Turabian StyleSantos, João Campos, Carla Cruz Marques, Maria Conceição Baptista, Jorge Pimenta, José Teixeira, Liliana Montezinho, Fernando Cagide, Fernanda Borges, Paulo J. Oliveira, and Rosa M. L. N. Pereira. 2022. "Effect of a Novel Hydroxybenzoic Acid Based Mitochondria Directed Antioxidant Molecule on Bovine Sperm Function and Embryo Production" Animals 12, no. 7: 804. https://doi.org/10.3390/ani12070804