
Factors Affecting the Efficiency of In Vitro Embryo Production in Prepubertal Mediterranean Water Buffalo
 , , , ,
, , , ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Animals
2.3. Ovarian Stimulation
2.4. Laparoscopic Ovum Pickup (LOPU)
2.5. Washing and Grading of COCs
2.6. In Vitro Embryo Production
2.6.1. In Vitro Maturation (IVM)
2.6.2. In Vitro Fertilization (IVF)
2.6.3. In Vitro Culture (IVC)
2.7. Embryo Vitrification and Thawing
2.8. Embryo Transfer and Pregnancy Detection
2.9. The Effect of Season
2.10. The Effect of Age
2.11. The Effect of Individual Variation
2.12. The Effect of Sire and Semen Concentration
2.13. Factors Impacting Embryo Transfer
2.14. Statistical Analysis
3. Results
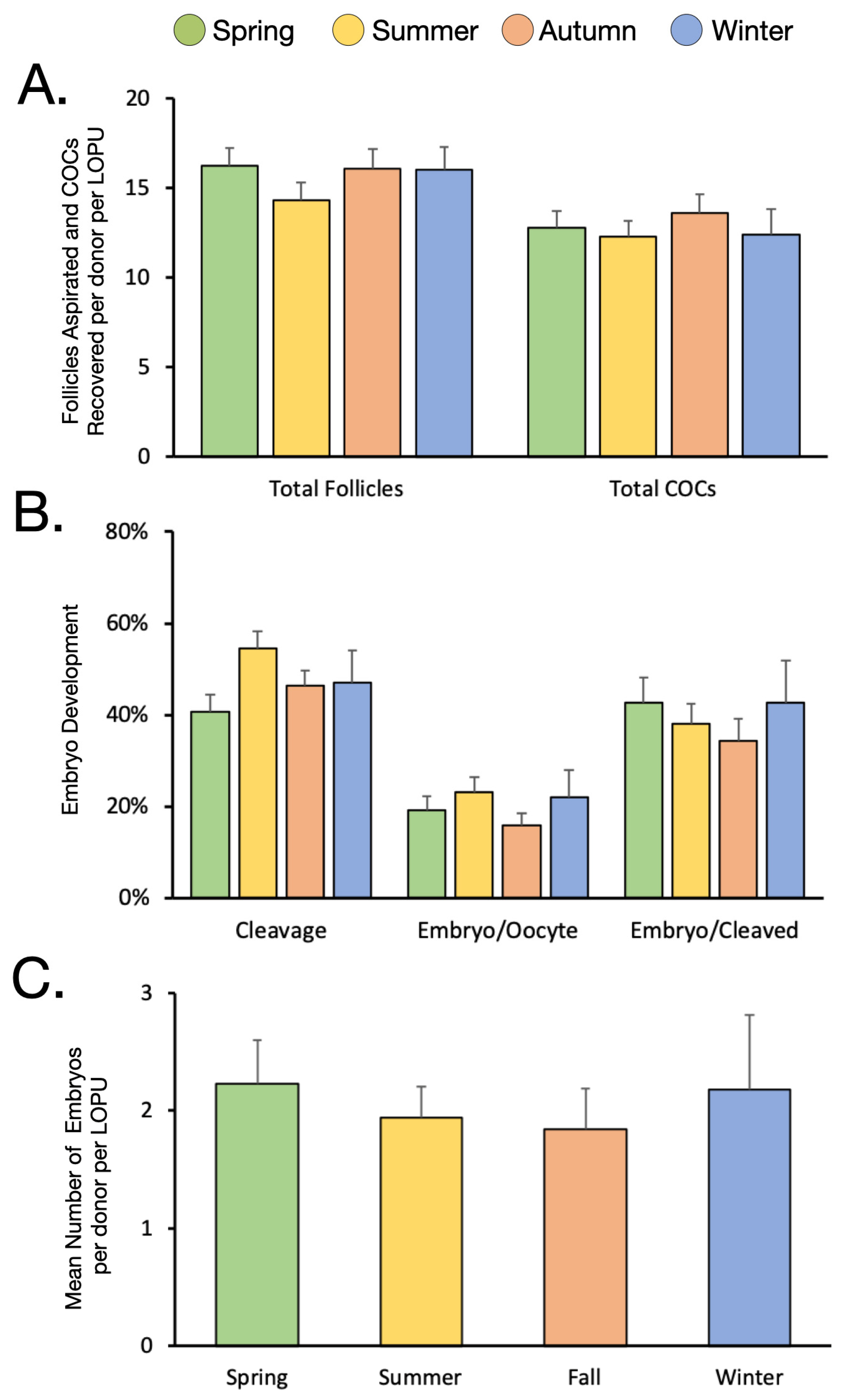
3.1. The Effect of Season
3.2. The Effect of Age
3.3. The Effect of Individual Variation
3.4. The Effect of Sire and Semen Concentration
3.5. The Effect of Embryo Transfer
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garau, V.; Manis, C.; Scano, P.; Caboni, P. Compositional Characteristics of Mediterranean Buffalo Milk and Whey. Dairy 2021, 2, 469–488. [Google Scholar] [CrossRef]
- Naveena, B.M.; Kiran, M. Buffalo meat quality, composition, and processing characteristics: Contribution to the global economy and nutritional security. Anim. Front. 2014, 4, 18–24. [Google Scholar] [CrossRef]
- Minervino, A.H.H.; Zava, M.; Vecchio, D.; Borghese, A. Bubalus bubalis: A Short Story. Front. Vet. Sci. 2020, 7, 570413. [Google Scholar] [CrossRef]
- Oswin Perera, B.M. Reproduction in water buffalo: Comparative aspects and implications for management. J. Reprod. Fertil. Suppl. 1999, 54, 157–168. [Google Scholar] [PubMed]
- Baruselli, P.S.; Soares, J.G.; Bayeux, B.M.; Silva, J.C.; Mingoti, R.D.; Carvalho, N.A. Assisted reproductive technologies (ART) in water buffaloes. Anim. Reprod. (AR) 2018, 15, 971–983. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.K.; Joshi, B.V.; Agrawala, P.L.; Kasiraj, R.; Sivaiah, S.; Randareddi, N.S.; Siddiqui, M.U. Multiple ovulation and embryo transfer in Indian buffaloes (Bubalus bubalis). Theriogenology 1990, 33, 289. [Google Scholar] [CrossRef]
- Salzano, A.; De Canditiis, C.; Della Ragione, F.; Prandi, A.; Zullo, G.; Neglia, G.; Campanile, G.; Gasparrini, B. Evaluation of factors involved in the failure of ovum capture in superovulated buffaloes. Theriogenology 2018, 122, 102–108. [Google Scholar] [CrossRef]
- Di Francesco, S.; Novoa, M.V.S.; Vecchio, D.; Neglia, G.; Boccia, L.; Campanile, G.; Zicarelli, L.; Gasparrini, B. Ovum pick-up and in vitro embryo production (OPU-IVEP) in Mediterranean Italian buffalo performed in different seasons. Theriogenology 2012, 77, 148–154. [Google Scholar] [CrossRef]
- Saliba, W.; Gimenes, L.; Drumond, R.; Bayao, H.; Alvim, M.; Baruselli, P.; Bastianetto, E.; Leite, R.; Gasparrini, B. Efficiency of OPU-IVEP-ET of fresh and vitrified embryos in buffaloes. Buffalo Bull. 2013, 32, 385–388. [Google Scholar]
- Gasparrini, B. Effects of reproductive season on embryo development in the buffalo. Reprod. Fertil. Dev. 2019, 31, 68–81. [Google Scholar] [CrossRef]
- Zicarelli, L. Considerations on the breeding and weaning of buffalo calf. J. Buffalo Sci. 2020, 9, 84–91. [Google Scholar] [CrossRef]
- Currin, L.; Baldassarre, H.; Bordignon, V. In vitro production of embryos from prepubertal holstein cattle and mediterranean water buffalo: Problems, progress and potential. Animals 2021, 11, 2275. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, H. Laparoscopic Ovum Pick-Up Followed by In Vitro Embryo Production and Transfer in Assisted Breeding Programs for Ruminants. Animals 2021, 11, 216. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, H.; Bordignon, V. Laparoscopic ovum pick-up for in vitro embryo production from dairy bovine and buffalo calves. Anim. Reprod. 2018, 15, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.B.; Rezende, R.G.; Colli, M.H.A.; Bayeux, B.M.; Mingoti, R.D.; Ojeda-Rojas, O.A.; Basso, A.C.; Naves, J.; Baruselli, P.S. In vitro embryo production in buffalo: Comparison between calves, prepubertal Heifers and lactating cows. Anim. Reprod. 2017, 14, 766. [Google Scholar]
- Currin, L.; Michalovic, L.; Bellefleur, A.-M.; Gutierrez, K.; Glanzner, W.; Schuermann, Y.; Bohrer, R.C.; Dicks, N.; da Rosa, P.R.; De Cesaro, M.P. The effect of age and length of gonadotropin stimulation on the in vitro embryo development of Holstein calf oocytes. Theriogenology 2017, 104, 87–93. [Google Scholar] [CrossRef]
- Steeves, T.; Gardner, D.; Zuelke, K.; Squires, T.; Fry, R. In vitro development and nutrient uptake by embryos derived from oocytes of pre-pubertal and adult cows. Mol. Reprod. Dev. 1999, 54, 49–56. [Google Scholar] [CrossRef]
- Baldassarre, H.; Currin, L.; Michalovic, L.; Bellefleur, A.-M.; Gutierrez, K.; Mondadori, R.G.; Glanzner, W.G.; Schuermann, Y.; Bohrer, R.C.; Dicks, N. Interval of gonadotropin administration for in vitro embryo production from oocytes collected from Holstein calves between 2 and 6 months of age by repeated laparoscopy. Theriogenology 2018, 116, 64–70. [Google Scholar] [CrossRef]
- Baldassarre, H.; Wang, B.; Pierson, J.; Neveu, N.; Sneek, L.; Lapointe, J.; Cote, F.; Kafidi, N.; Keefer, C.; Lazaris, A. Prepubertal propagation of transgenic cloned goats by laparoscopic ovum pick-up and in vitro embryo production. Cloning Stem Cells 2004, 6, 25–29. [Google Scholar] [CrossRef]
- Leoni, G.G.; Succu, S.; Satta, V.; Paolo, M.; Bogliolo, L.; Bebbere, D.; Spezzigu, A.; Madeddu, M.; Berlinguer, F.; Ledda, S. In vitro production and cryotolerance of prepubertal and adult goat blastocysts obtained from oocytes collected by laparoscopic oocyte-pick-up (LOPU) after FSH treatment. Reprod. Fertil. Dev. 2009, 21, 901–908. [Google Scholar] [CrossRef]
- O’Brien, J.; Catt, S.; Ireland, K.; Maxwell, W.; Evans, G. In vitro and in vivo developmental capacity of oocytes from prepubertal and adult sheep. Theriogenology 1997, 47, 1433–1443. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Dwarte, D.; Ryan, J.; Maxwell, W.; Evans, G. Developmental capacity, energy metabolism and ultrastructure of mature oocytes from prepubertal and adult sheep. Reprod. Fertil. Dev. 1996, 8, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Currin, L.; Baldassarre, H.; Priotto de Macedo, M.; Giehl Glanzner, W.; Gutierrez, K.; Lazaris, K.; da Silva, Z.; Guay, V.; Carrillo Herrera, M.E.; Brown, C.; et al. Optimization of gonadotropin stimulation protocols for in vitro embryo production in prepubertal Mediterranean water buffalo. Theriogenology 2023, 197, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, B.; Neglia, G.; Di Palo, R.; Vecchio, D.; Albero, G.; Esposito, L.; Campanile, G.; Zicarelli, L. Influence of oocyte donor on in vitro embryo production in buffalo. Anim. Reprod. Sci. 2014, 144, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, S.; Boccia, L.; Campanile, G.; Di Palo, R.; Vecchio, D.; Neglia, G.; Zicarelli, L.; Gasparrini, B. The effect of season on oocyte quality and developmental competence in Italian Mediterranean buffaloes (Bubalus bubalis). Anim. Reprod. Sci. 2011, 123, 48–53. [Google Scholar] [CrossRef]
- Shahzad, Q.; Waqas, M.; Pu, L.; Wadood, A.A.; Xie, L.; Husna, A.U.; Yang, K.; Wang, J.; Xu, H.; Lu, K. Seasonality and photoperiod influence in vitro production of buffalo embryos. Reprod. Domest. Anim. 2020, 55, 1115–1123. [Google Scholar] [CrossRef]
- Chemineau, P.; Guillaume, D.; Migaud, M.; Thiéry, J.; Pellicer-Rubio, M.; Malpaux, B. Seasonality of Reproduction in Mammals: Intimate Regulatory Mechanisms and Practical Implications. Reprod. Domest. Anim. 2008, 43, 40–47. [Google Scholar] [CrossRef]
- D’Occhio, M.J.; Ghuman, S.S.; Neglia, G.; della Valle, G.; Baruselli, P.S.; Zicarelli, L.; Visintin, J.A.; Sarkar, M.; Campanile, G. Exogenous and endogenous factors in seasonality of reproduction in buffalo: A review. Theriogenology 2020, 150, 186–192. [Google Scholar] [CrossRef]
- Campanile, G.; Neglia, G.; Gasparrini, B.; Galiero, G.; Prandi, A.; Palo, R.D.; D’Occhio, M.J.; Zicarelli, L. Embryonic mortality in buffaloes synchronized and mated by AI during the seasonal decline in reproductive function. Theriogenology 2005, 63, 2334–2340. [Google Scholar] [CrossRef]
- Vecchio, D.; Neglia, G.; Gasparrini, B.; Russo, M.; Pacelli, C.; Prandi, A.; D’Occhio, M.J.; Campanile, G. Corpus luteum development and function and relationship to pregnancy during the breeding season in the Mediterranean buffalo. Theriogenology 2012, 77, 1811–1815. [Google Scholar] [CrossRef]
- Di Francesco, S.; Neglia, G.; Vecchio, D.; Rossi, P.; Russo, M.; Zicarelli, L.; D’Occhio, M.J.; Campanile, G. Influence of season on corpus luteum structure and function and AI outcome in the Italian Mediterranean buffalo (Bubalus bubalis). Theriogenology 2012, 78, 1839–1845. [Google Scholar] [CrossRef] [PubMed]
- Neglia, G.; de Nicola, D.; Esposito, L.; Salzano, A.; D’Occhio, M.J.; Fatone, G. Reproductive management in buffalo by artificial insemination. Theriogenology 2020, 150, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Misztal, T.; Romanowicz, K.; Barcikowski, B. Melatonin-a modulator of the GnRH/LH axis in sheep. Reprod. Biol. 2002, 2, 267–275. [Google Scholar] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkins, J.A.; Pohler, K.G.; Smith, M.F. Physiology and endocrinology of puberty in heifers. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 479–492. [Google Scholar] [CrossRef]
- Watanabe, Y.F.; de Souza, A.H.; Mingoti, R.D.; Ferreira, R.M.; Batista, E.O.S.; Dayan, A.; Watanabe, O.; Meirelles, F.V.; Nogueira, M.F.G.; Ferraz, J.B.S. Number of oocytes retrieved per donor during OPU and its relationship with in vitro embryo production and field fertility following embryo transfer. Anim. Reprod. (AR) 2018, 14, 635–644. [Google Scholar] [CrossRef]
- De Paz, P.; Sánchez, A.; De la Fuente, J.; Chamorro, C.; Alvarez, M.; Anel, E.; Anel, L. Ultrastructural and cytochemical comparison between calf and cow oocytes. Theriogenology 2001, 55, 1107–1116. [Google Scholar] [CrossRef]
- Kauffold, J.; Amer, H.A.H.; Bergfeld, U.; Weber, W.; Sobiraj, A. The In Vitro Developmental Competence of Oocytes from Juvenile Calves is Related to Follicular Diameter. J. Reprod. Dev. 2005, 51, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Michalovic, L.; Currin, L.; Gutierrez, K.; Bellefleur, A.M.; Glanzner, W.G.; Schuermann, Y.; de Macedo, M.P.; Bohrer, R.C.; Dicks, N.; Lopez, R. Granulosa cells of prepubertal cattle respond to gonadotropin signaling and upregulate genes that promote follicular growth and prevent cell apoptosis. Mol. Reprod. Dev. 2018, 85, 909–920. [Google Scholar] [CrossRef]
- Tervit, H. Laparoscopy/laparotomy oocyte recovery and juvenile breeding. Anim. Reprod. Sci. 1996, 42, 227–238. [Google Scholar] [CrossRef]
- Presicce, G.A.; Senatore, E.M.; De Santis, G.; Stecco, R.; Terzano, G.M.; Borghese, A.; De Mauro, G.J. Hormonal stimulation and oocyte maturational competence in prepuberal Mediterranean Italian buffaloes (Bubalus bubalis). Theriogenology 2002, 57, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Levesque, J.; Sirard, M. Proteins in oocytes from calves and adult cows before maturation: Relationship with their development capacity. Reprod. Nutr. Dev. 1994, 34, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Sherrer, E.; Rathbun, T.; Davis, D. Fertilization and blastocyst development in oocytes obtained from prepubertal and adult pigs. J. Anim. Sci. 2004, 82, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, K.; Takahashi, Y. Comparison of maturational and developmental parameters of oocytes recovered from prepubertal and adult pigs. Reprod. Fertil. Dev. 2003, 15, 215–221. [Google Scholar] [CrossRef]
- Baldassarre, H.; Wang, B.; Kafidi, N.; Keefer, C.; Lazaris, A.; Karatzas, C. Advances in the production and propagation of transgenic goats using laparoscopic ovum pick-up and in vitro embryo production technologies. Theriogenology 2002, 57, 275–284. [Google Scholar] [CrossRef]
- Baldassarre, H.; Karatzas, C. Advanced assisted reproduction technologies (ART) in goats. Anim. Reprod. Sci. 2004, 82, 255–266. [Google Scholar] [CrossRef]
- Baruselli, P.S.; de Carvalho, J.G.S.; Elliff, F.M.; da Silva, J.C.B.; Chello, D.; de Carvalho, N.A.T. Embryo transfer in buffalo (Bubalus bubalis). Theriogenology 2020, 150, 221–228. [Google Scholar] [CrossRef]
- Baldassarre, H.; De Matos, D.; Furnus, C.; Castro, T.; Fischer, E.C. Technique for efficient recovery of sheep oocytes by laparoscopic folliculocentesis. Anim. Reprod. Sci. 1994, 35, 145–150. [Google Scholar] [CrossRef]
- Vajta, G.; Holm, P.; Kuwayama, M.; Booth, P.J.; Jacobsen, H.; Greve, T.; Callesen, H. Open pulled straw (OPS) vitrification: A new way to reduce cryoinjuries of bovine ova and embryos. Mol. Reprod. Dev. 1998, 51, 53–58. [Google Scholar] [CrossRef]
- Government of Canada; Environment and Climate Change Canada (ECCC). Daily Data Report for Ste-Anne-de-Bellevue 1; 2022. Available online: https://climate.weather.gc.ca/ (accessed on 4 December 2022).
- Campanile, G.; Neglia, G.; D’Occhio, M.J. Embryonic and fetal mortality in river buffalo (Bubalus bubalis). Theriogenology 2016, 86, 207–213. [Google Scholar] [CrossRef]
- Saliba, W.P.; Gimenes, L.U.; Drumond, R.M.; Bayão, H.X.S.; Di Palo, R.; Gasparrini, B.; Rubessa, M.; Baruselli, P.S.; Sales, J.N.S.; Bastianetto, E.; et al. “Which Factors Affect Pregnancy Until Calving and Pregnancy Loss in Buffalo Recipients of in vitro Produced Embryos?”. Front. Vet. Sci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- da Silva, J.C.B.; Alves, M.B.R.; Bridi, A.; Bohrer, R.C.; Escobar, G.S.L.; de Carvalho, J.A.B.A.; Binotti, W.A.B.; Pugliesi, G.; Lemes, K.M.; Chello, D.; et al. Reproductive seasonality influences oocyte retrieval and embryonic competence but not uterine receptivity in buffaloes. Theriogenology 2021, 170, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Zicarelli, L. Enhancing reproductive performance in domestic dairy water buffalo (Bubalus bubalis). In Reproduction in Domestic Ruminants VII; Lucy, M., Pate, J., Smith, M., Spencer, T., Eds.; Nottingham University Press: Anchorage, AK, USA, 2010; pp. 443–455. [Google Scholar]
- Barile, V.L. Reproductive efficiency in female buffaloes. In Buffalo Production and Research; Borghese, A., Ed.; Food and Agriculture Organization of the United Nations: Rome, Italy, 2005; Volume 67, pp. 77–107. [Google Scholar]
- Duby, R.T.; Damiani, P.; Looney, C.R.; Fissore, R.A.; Robl, J.M. Prepuberal calves as oocyte donors: Promises and problems. Theriogenology 1996, 45, 121–130. [Google Scholar] [CrossRef]
- Totey, S.; Singh, G.; Taneja, M.; Pawshe, C.; Talwar, G. In vitro maturation, fertilization and development of follicular oocytes from buffalo (Bubalus bubalis). Reproduction 1992, 95, 597–607. [Google Scholar] [CrossRef]
- Das, G.; Jain, G.; Solanki, V.; Tripathi, V. Efficacy of various collection methods for oocyte retrieval in buffalo. Theriogenology 1996, 46, 1403–1411. [Google Scholar] [CrossRef]
- Kumar, A.; Solanki, V.S.; Jindal, S.K.; Tripathi, V.N.; Jain, G.C. Oocyte retrieval and histological studies of follicular population in buffalo ovaries. Anim. Reprod. Sci. 1997, 47, 189–195. [Google Scholar] [CrossRef]
- Yousaf, M.R.; Chohan, K.R. Nuclear morphology, diameter and meiotic competence of buffalo oocytes relative to follicle size. Reprod. Fertil. Dev. 2003, 15, 223–229. [Google Scholar] [CrossRef]
- Pierson, J.; Wang, B.; Neveu, N.; Sneek, L.; Cote, F.; Karatzas, C.; Baldassarre, H. Effects of repetition, interval between treatments and season on the results from laparoscopic ovum pick-up in goats. Reprod. Fertil. Dev. 2004, 16, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Liang, A.; Salzano, A.; D’Esposito, M.; Comin, A.; Montillo, M.; Yang, L.; Campanile, G.; Gasparrini, B. Anti-Mullerian hormone (AMH) concentration in follicular fluid and mRNA expression of AMH receptor type II and LH receptor in granulosa cells as predictive markers of good buffalo (Bubalus bubalis) donors. Theriogenology 2016, 86, 963–970. [Google Scholar] [CrossRef]
- Batista, E.; Guerreiro, B.; Freitas, B.; Silva, J.; Vieira, L.; Ferreira, R.; Rezende, R.; Basso, A.; Lopes, R.; Rennó, F. Plasma anti-Müllerian hormone as a predictive endocrine marker to select Bos taurus (Holstein) and Bos indicus (Nelore) calves for in vitro embryo production. Domest. Anim. Endocrinol. 2016, 54, 1–9. [Google Scholar] [CrossRef]
- Rubessa, M.; Di Fenza, M.; Mariotti, E.; Di Francesco, S.; de Dilectis, C.; Di Palo, R.; Zicarelli, L.; Gasparrini, B. Kinetics of Sperm Penetration is Correlated with In Vitro Fertility of Buffalo (Bubalus bubalis) Bulls. Reprod. Fertil. Dev. 2008, 21, 206–207. [Google Scholar] [CrossRef]
- Sun, W.; Li, Y.; Su, J.; Bao, X.; Ding, R.; Zhao, G.; Cao, G.; Hu, S.; Wang, J.; Sun, Q.; et al. Correlation between in vitro fertilization and artificial insemination in Holstein bulls. Anim. Biosci. 2021, 34, 1879–1885. [Google Scholar] [CrossRef] [PubMed]
- Zicarelli, L.; Mariotti, E.; Di Francesco, S.; Velotto, S.; Rubessa, M.; Neglia, G. Effect of bull on in vitro sperm capacitation induced by different agents in buffalo species (Bubalus bubalis). Ital. J. Anim. Sci. 2009, 8, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Totey, S.M.; Pawshe, C.H.; Singh, G.P. Effects of bull and heparin and sperm concentrations on in vitro fertilization of buffalo (Bubalusbubalis) oocytes matured in vitro. Theriogenology 1993, 39, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Gasparrini, B. In vitro embryo production in buffalo species: State of the art. Theriogenology 2002, 57, 237–256. [Google Scholar] [CrossRef]
- Boccia, L.; Rosa, A.D.; Attanasio, L.; Neglia, G.; Vecchio, D.; Campanile, G.; Zicarelli, L.; Gasparrini, B. Developmental Speed Affects the Cryotolerance of In Vitro Produced Buffalo (Bubalus bubalis) Embryos. Ital. J. Anim. Sci. 2013, 12, e80. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Chaves, M.S.; Silva, A.F.B.; Vale, W.G.; Negreiros, N.A.B.; Arcce, I.M.L.; Melo, L.M.; Freitas, V.J.d.F. Factors affecting the cryopreservation of oocytes and embryos in buffalo (Bubalus bubalis): A review. Res. Soc. Dev. 2022, 11, e25111427337. [Google Scholar] [CrossRef]
- Albarracín, J.L.; Morató, R.; Rojas, C.; Mogas, T. Effects of vitrification in open pulled straws on the cytology of in vitro matured prepubertal and adult bovine oocytes. Theriogenology 2005, 63, 890–901. [Google Scholar] [CrossRef]
- Dubeibe Marin, D.F.; Nogueira da Costa, N.; di Paula Bessa Santana, P.; Baia de Souza, E.; Rolim filho, S.T.; da Silva Cordeiro, M.; Ohashi, O.M. Influence of l-carnitine on lipid metabolism of buffalo cumulus-oocyte complexes matured in either fetal bovine serum or fatty acid-free bovine serum albumin. Theriogenology 2020, 158, 382–390. [Google Scholar] [CrossRef]
- Dubeibe Marin, D.F.; da Costa, N.N.; di Paula Bessa Santana, P.; de Souza, E.B.; Ohashi, O.M. Importance of lipid metabolism on oocyte maturation and early embryo development: Can we apply what we know to buffalo? Anim. Reprod. Sci. 2019, 211, 106220. [Google Scholar] [CrossRef]
- Zolini, A.M.; Carrascal-Triana, E.; de King, A.R.; Hansen, P.J.; Torres, C.A.A.; Block, J. Effect of addition of L-carnitine to media for oocyte maturation and embryo culture on development and cryotolerance of bovine embryos produced in vitro. Theriogenology 2019, 133, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Inaba, Y.; Somfai, T.; Kaneda, M.; Geshi, M.; Nagai, T.; Manabe, N. Supplementation of culture medium with L-carnitine improves development and cryotolerance of bovine embryos produced in vitro. Reprod. Fertil. Dev. 2013, 25, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.; Pandey, S.; Bhat, I.A.; Mukesh, B.; Anand, J.; Chandra, V.; Sharma, G.T. Impact of L-carnitine on lipid content and post thaw survivability of buffalo embryos produced in vitro. Cryobiology 2018, 82, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Boccia, L.; De Blasi, M.; Zullo, G.; Longobardi, V.; Vecchio, D.; Gasparrini, B. L-Carnitine during in vitro culture enhances the cryotolerance of buffalo (Bubalus bubalis) in vitro-derived embryos. Reprod. Fertil. Dev. 2012, 25, 214. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Mean T (°C) | Min T (°C) | Max T (°C) | Humidity (%) | Precipitation (mm) | Daylength (h) |
|---|---|---|---|---|---|---|
| Mar–May | 7.51 ± 7.2 | 2.41 ± 6.8 | 12.60 ± 8.1 | 63.8 ± 14.8 | 2.76 ± 6.9 | 13.55 ± 1.2 |
| Jun–Aug | 19.46 ± 3.4 | 14.51 ± 3.5 | 24.39 ± 3.7 | 68.4 ± 12.6 | 3.39 ± 7.8 | 15.30 ± 0.7 |
| Sept–Nov | 8.34 ± 6.4 | 3.82 ± 6.1 | 12.85 ± 7.3 | 73.3 ± 10.0 | 2.48 ± 5.0 | 10.69 ± 1.2 |
| Dec–Feb | −6.07 ± 6.0 | −10.92 ± 7.0 | −1.22 ± 6.0 | 73.3 ± 10.5 | 4.18 ± 7.7 | 9.80 ± 0.8 |
| Season | N | Pregnancies |
|---|---|---|
| Mar–May | 31 | 8 (25.8%) |
| June–Aug | 31 | 7 (22.6%) |
| Sept–Nov | 23 | 7 (30.4%) |
| Dec–Feb | 33 | 11 (33.3%) |
| Variable | Treatment | N | Pregnancies |
|---|---|---|---|
| Number of embryos transferred | 1 | 18 | 4 (22.2%) |
| 2 | 99 | 29 (29.3%) | |
| Stage of embryo development | Morula | 20 | 4 (20.0%) |
| Blastocyst | 19 | 9 (47.4%) | |
| Hatched | 15 | 4 (26.7%) | |
| Vitrification | Fresh | 87 | 25 (28.7%) |
| Vitrified | 28 | 8 (28.6%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Currin, L.; Baldassarre, H.; de Macedo, M.P.; Glanzner, W.G.; Gutierrez, K.; Lazaris, K.; Guay, V.; Herrera, M.E.C.; da Silva, Z.; Brown, C.; et al. Factors Affecting the Efficiency of In Vitro Embryo Production in Prepubertal Mediterranean Water Buffalo. Animals 2022, 12, 3549. https://doi.org/10.3390/ani12243549
Currin L, Baldassarre H, de Macedo MP, Glanzner WG, Gutierrez K, Lazaris K, Guay V, Herrera MEC, da Silva Z, Brown C, et al. Factors Affecting the Efficiency of In Vitro Embryo Production in Prepubertal Mediterranean Water Buffalo. Animals. 2022; 12(24):3549. https://doi.org/10.3390/ani12243549
Chicago/Turabian StyleCurrin, Luke, Hernan Baldassarre, Mariana Priotto de Macedo, Werner Giehl Glanzner, Karina Gutierrez, Katerina Lazaris, Vanessa Guay, María Elena Carrillo Herrera, Zigomar da Silva, Caitlin Brown, and et al. 2022. "Factors Affecting the Efficiency of In Vitro Embryo Production in Prepubertal Mediterranean Water Buffalo" Animals 12, no. 24: 3549. https://doi.org/10.3390/ani12243549