
Association between Synonymous SNPs of SOX10 and Plumage Color and Reproductive Traits of Ducks

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples
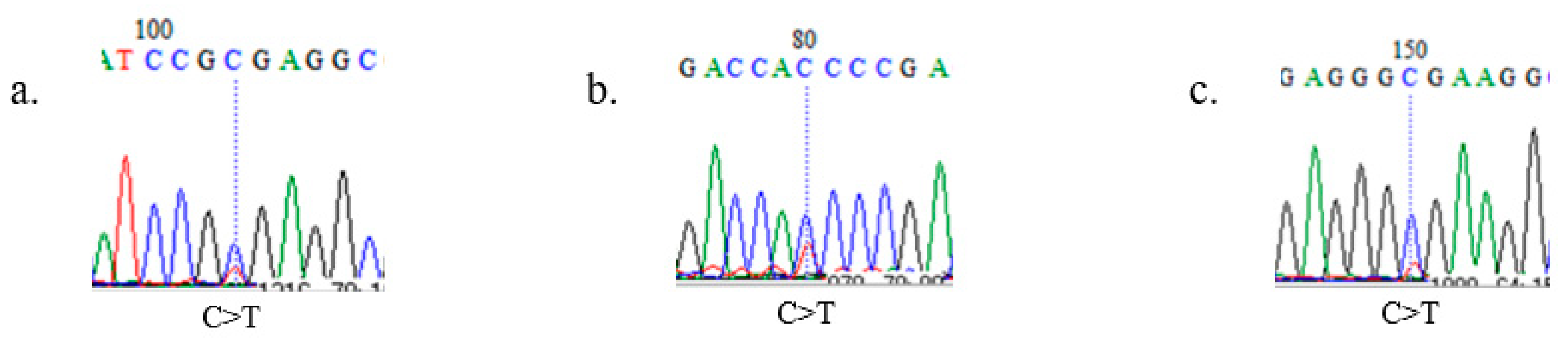
2.2. SOX10 Gene Sequencing and Identification of Polymorphisms
2.3. Trait Records
2.4. Statistics
3. Results
3.1. Segregation of Plumage Color and Reproductive Traits in F2 Ducks
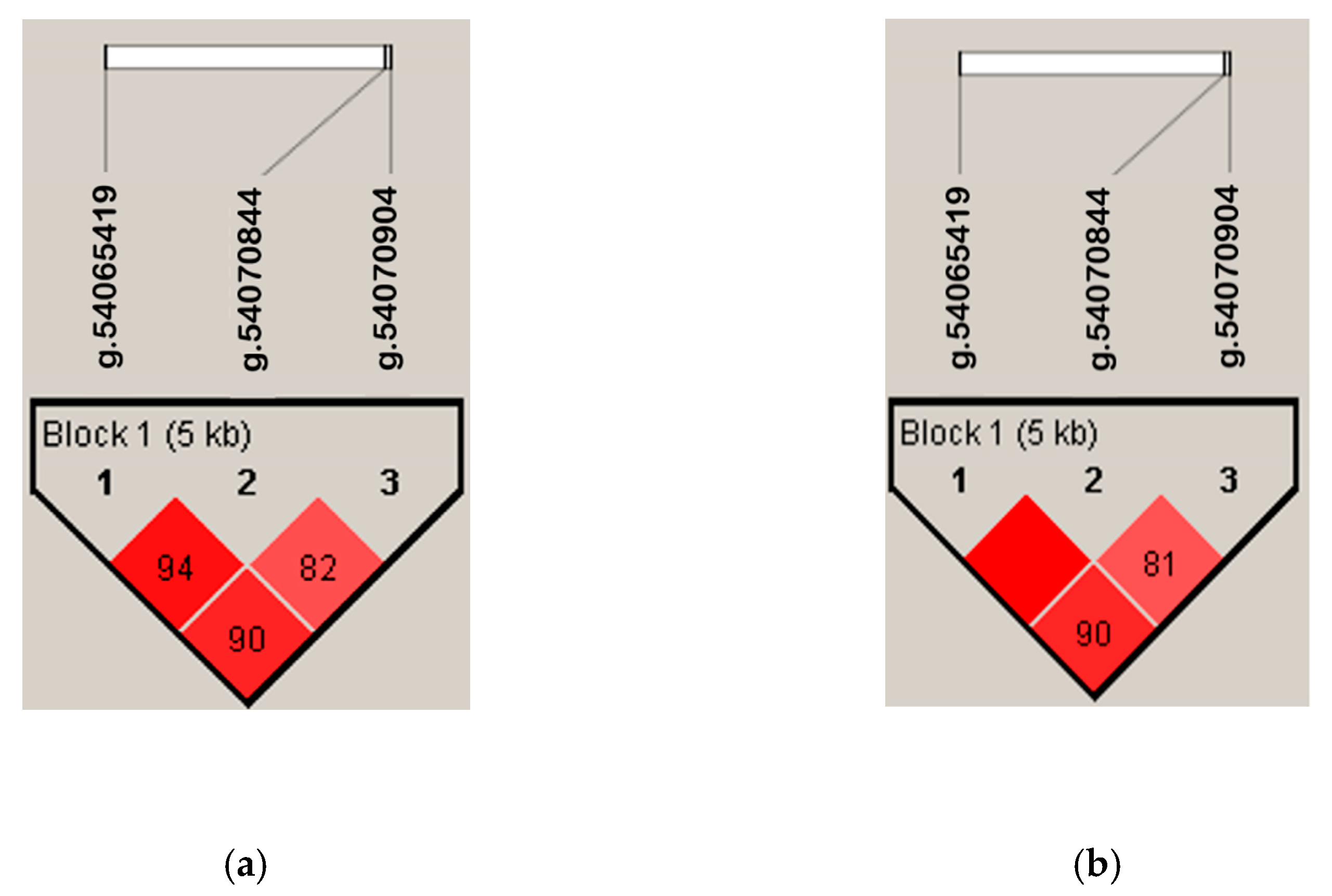
3.2. SNP Identification, Linkage Disequilibrium, and Haplotypes
3.3. Genotype Association of Duck Plumage Color
3.4. Association of Duck Reproductive Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g.54065419 | g.54070844 | g.54070904 | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenotype | C/C | C/T | T/T | p-Value | C/C | C/T | T/T | p-Value | C/C | C/T | T/T | p-Value |
| ——————————————————————————————————Mean——————————————————————————————————— | ||||||||||||
| Age at sexual maturity (days) | 119.6 ± 15.7 b | 115.1 ± 14.0 b | 135.3 ± 12.9 a | 7.0 × 10−3 * | 120.5 ± 14.5 | 124.2 ± 14.9 | 113.6 ± 17.4 | 0.11 | 124.3 ± 14.8 | 116.0 ± 15.0 | 116.2 ± 16.5 | 0.48 |
| Body weight at sexual maturity (g) | 1808.0 ± 242.2 | 1804.0 ± 203.7 | 1876.0 ± 210.9 | 0.38 | 1805.0 ± 234.9 | 1825.0 ± 207.6 | 1828.0 ± 251.4 | 0.38 | 1823.0 ± 229.2 | 1800.0 ± 213.8 | 1773.0 ± 213.1 | 0.17 |
| Egg weight at sexual maturity (g) | 48.8 ± 9.8 | 42.9 ± 8.8 | 50.9 ± 7.0 | 0.02 | 47.6 ± 9.6 | 48.2 ± 8.9 | 44.9 ± 9.0 | 0.36 | 47.1 ± 9.1 | 45.5 ± 9.9 | 52.3 ± 6.9 | 0.13 |
| Egg weight at peak production (g) | 67.4 ± 7.0 | 65.5 ± 6.7 | 65.1 ± 5.9 | 0.84 | 66.3 ± 6.7 | 66.3 ± 7.2 | 67.3 ± 6.5 | 0.97 | 66.3 ± 7.0 | 66.2 ± 6.4 | 68.0 ± 5.2 | 0.85 |
| Egg production at 120 days | 2.9 ± 4.3 | 2.6 ± 4.1 | 1.3 ± 3.2 | 0.18 | 2.6 ± 4.1 | 2.8 ± 4.2 | 2.8 ± 4.7 | 0.79 | 2.7 ± 4.3 | 2.7 ± 3.8 | 3.1 ± 5.3 | 0.92 |
| Egg production at 150 days | 22.4 ± 11.5 a | 23.0 ± 10.0 a | 11.2 ± 10.6 b | 4.0 × 10−3 * | 22.4 ± 10.8 | 22.6 ± 10.9 | 20.3 ± 11.5 | 0.57 | 22.4 + 11.0 | 22.6 + 10.7 | 20.3 + 9.7 | 0.81 |
| Egg production at 180 days | 48.54 ± 15.1 | 50.8 ± 11.2 | 42.5 ± 11.9 | 0.21 | 49.2 ± 13.2 | 49.1 ± 13.9 | 45.9 ± 13.4 | 0.41 | 49.2 + 13.6 | 49.5 + 13.7 | 45.9 + 10.5 | 0.39 |
| Egg production at 210 days | 74.7 ± 17.7 | 78.6 ± 11.6 | 68.1 ± 11.8 | 0.14 | 76.4 ± 14.6 | 76.1 ± 15.9 | 71.7 ± 15.9 | 0.28 | 76.3 + 15.1 | 75.3 + 16.1 | 73.3 + 12.4 | 0.69 |
| Egg production at 240 days | 100.5 ± 20.0 | 110.7 ± 12.1 | 86.2 ± 11.4 | 0.02 | 103.7 ± 16.2 | 103.3 ± 16.9 | 94.9 ± 20.3 | 0.03 | 103.3 + 16.7 | 102.7 + 16.8 | 93.0 + 22.5 | 0.16 |
| Egg production at 270 days | 126.9 ± 22.1 | 132.9 ± 12.5 | 118.0 ± 10.2 | 0.16 | 129.6 ± 17.8 | 129.7 ± 17.7 | 118.6 ± 24.3 | 0.07 | 129.3 + 18.1 | 129.3 + 17.6 | 114.0 + 30.0 | 0.19 |
| Egg production at 300 days | 153.6 ± 23.3 | 161.3 ± 12.4 | 143.3 ± 9.8 | 0.18 | 157.2 ± 18.7 a | 157.0 ± 18.6 a | 145.3 ± 24.9 b | 6.0 × 10−3 * | 156.9 + 19.0 | 156.2 + 18.7 | 139.9 + 29.1 | 0.07 |
| Egg production at 330 days | 180.4 ± 25.2 | 189.6 ± 12.5 | 171.6 ± 9.7 | 0.11 | 185.0 ± 20.0 | 184.4 ± 20.0 | 170.9 ± 26.0 | 0.04 | 184.7 + 20.1 | 183.2 + 20.8 | 163.2 + 27.8 | 0.03 |
| Egg production at 360 days | 206.7 ± 28.0 | 217.8 ± 13.1 | 199.3 ± 9.0 | 0.02 | 212.0 ± 22.2 a | 211.4 ± 21.6 a | 196.7 ± 28.7 b | 7.0 × 10−3 * | 211.7 + 21.9 a | 210.1 + 23.2 a | 187.2 + 30.4 b | 1.0 × 10−3 ** |
| Egg production at 390 days | 231.8 ± 30.9 ab | 244.3 ± 14.0 a | 225.3 ± 11.5 b | 5.0 × 10−3 * | 238.1 ± 24.8 a | 237.7 ± 23.1 a | 220.0 ± 32.1 b | 1.0 × 10−3 ** | 237.8 + 24.3 a | 236.8 + 24.6 a | 208.7 + 35.8 b | 1.0 × 10−3 ** |
| Egg production at 420 days | 255.7 ± 34.4 ab | 270.6 ± 15.4 a | 250.7 ± 14.4 b | 7.0 × 10−4 ** | 263.2 ± 28.0 a | 262.8 ± 25.1 a | 242.0 ± 37.5 b | 7.0 × 10−3 * | 262.0 + 27.4 a | 260.8 + 26.5 a | 230.0 + 43.1 b | 1.0 × 10−3 ** |
| Shell strength (kgf/cm2) | 4.8 ± 1.0 | 4.8 ± 0.9 | 4.4 ± 1.3 | 0.87 | 4.8 ± 1.0 | 4.6 ± 1.0 | 4.8 ± 0.8 | 0.41 | 4.5 ± 1.0 | 4.9 ± 1.0 | 5.2 ± 0.3 | 0.09 |
| Shape index | 1.34 ± 0.06 | 1.33 ± 0.07 | 1.33 ± 0.05 | 0.22 | 1.34 ± 0.07 | 1.34 ± 0.06 | 1.35 ± 0.03 | 0.93 | 1.34 ± 0.07 | 1.35 ± 0.05 | 1.34 ± 0.02 | 0.93 |
| Haugh unit (HU) | 79.9 ± 10.9 | 82.4 ± 11.7 | 80.0 ± 12.5 | 0.32 | 82.3 ± 9.7 | 79.64 ± 13.1 | 79.2 ± 8.8 | 0.03 | 81.8 ± 11.1 | 79.4 ± 12.2 | 83.5 ± 5.3 | 0.19 |
| Yolk color index | 4.2 ± 0.6 | 4.2 ± 0.6 | 4.5 ± 0.7 | 0.20 | 4.2 ± 0.6 | 4.3 ± 0.6 | 3.9 ± 0.8 | 0.48 | 4.2 ± 0.6 | 4.2 ± 0.6 | 4.2 ± 0.6 | 0.80 |
| Parameter | r-Value | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ASM | BWSM | EWSM | EP120 | EP150 | EP180 | EP210 | EP240 | EP270 | EP300 | EP330 | EP360 | EP390 | EP420 | HU | YCI | |
| ASM | 1.00 | |||||||||||||||
| BWSM | 0.39 * | 1.00 | ||||||||||||||
| EWSM | 0.35 ** | 0.28 ** | 1.00 | |||||||||||||
| EP120 | −0.64 ** | −0.29 ** | −0.31 ** | 1.00 | ||||||||||||
| EP150 | −0.78 ** | −0.35 ** | −0.38 ** | 0.75 ** | 1.00 | |||||||||||
| EP180 | −0.75 ** | −0.34 ** | −0.38 ** | 0.66 ** | 0.94 ** | 1.00 | ||||||||||
| EP210 | −0.69 ** | −0.31 ** | −0.36 ** | 0.58 ** | 0.86 ** | 0.95 ** | 1.00 | |||||||||
| EP240 | −0.61 ** | −0.30 ** | −0.34 ** | 0.52 ** | 0.76 ** | 0.86 ** | 0.96 ** | 1.00 | ||||||||
| EP270 | −0.55 ** | −0.29 ** | −0.31 ** | 0.46 ** | 0.68 ** | 0.77 ** | 0.88 ** | 0.96 ** | 1.00 | |||||||
| EP300 | −0.50 ** | −0.27 ** | −0.30 ** | 0.43 ** | 0.62 ** | 0.71 ** | 0.82 ** | 0.91 ** | 0.97 ** | 1.00 | ||||||
| EP330 | −0.46 ** | −0.25 ** | −0.29 ** | 0.40 ** | 0.58 ** | 0.66 ** | 0.77 ** | 0.86 ** | 0.92 ** | 0.97 ** | 1.00 | |||||
| EP360 | −0.42 ** | −0.22 ** | −0.28 ** | 0.37 ** | 0.54 ** | 0.62 ** | 0.72 ** | 0.80 ** | 0.86 ** | 0.92 ** | 0.98 ** | 1.00 | ||||
| EP390 | −0.38 ** | −0.18 ** | −0.25 ** | 0.34 ** | 0.49 ** | 0.56 ** | 0.66 ** | 0.74 ** | 0.80 ** | 0.85 ** | 0.92 ** | 0.98 ** | 1.00 | |||
| EP420 | −0.34 ** | −0.16 ** | −0.24 ** | 0.31 ** | 0.44 ** | 0.52 ** | 0.61 ** | 0.68 ** | 0.74 ** | 0.80 ** | 0.87 ** | 0.93 ** | 0.98 ** | 1.00 | ||
| HU | −0.006 | 0.011 | −0.003 | 0.024 | 0.042 | 0.009 | −0.010 | −0.025 | −0.049 | −0.065 | −0.065 | −0.065 | 0.075 * | 0.082 * | 1.00 | |
| YCI | 0.075 * | 0.057 | −0.011 | −0.049 | −0.042 | −0.047 | −0.058 | −0.077* | −0.056 | −0.056 | −0.061 | −0.060 | −0.064 | −0.072 * | −0.143 ** | 1.00 |
References
- Berri, C. 2-Breeding and quality of poultry. In Poultry Meat Processing and Quality; Mead, G.C., Ed.; Woodhead Publishing: Sawston, UK, 2004; pp. 21–37. [Google Scholar]
- Moran, E.T. 3-Husbandry techniques. In Poultry Meat Processing and Quality; Mead, G.C., Ed.; Woodhead Publishing: Sawston, UK, 2004; pp. 38–64. [Google Scholar]
- CFDIA. Several Real Facts about China Down Industry. China Feather and Down Industrial Association, Beijing. 2015. Available online: http://www.cfd.com.cn/en/content/details_117_1262.html (accessed on 5 January 2016).
- CFDIA. China Feather and Down Industry Exports in First Three Quarters of 2015, China Feather and Down Industrial Association, Beijing. 2015. Available online: http://www.cfd.com.cn/en/content/details_108_1332.html (accessed on 5 January 2016).
- Sonaiya, E.B.; Swan, S.E.J. Small scale poultry production. FAO Anim. Prod. Health 2004, 1–224. [Google Scholar]
- Jennings, C.; Cale, W. USA Down & Feather Labeling Standards-Apparel (Down Jackets, Vests, Boots, etc). In Down Clothing Does not Require the Complex Law Tag Label Needed for Bedding Products. Fill can be Listed Using the Sub-Label pe. 2015. Available online: https://alliedfeather.com/wp-content/uploads/2021/08/IDFL_Standards_-_USA_Jackets_Labeling.pdf (accessed on 5 January 2016).
- Gong, Y.; Yang, Q.; Li, S.; Feng, Y.; Gao, C.; Tu, G.; Peng, X. Grey plumage colouration in the duck is genetically determined by the alleles on two different, interacting loci. Anim. Genet. 2010, 41, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Gunnarsson, U.; Kerje, S.; Bed’Hom, B.; Sahlqvist, A.-S.; Ekwall, O.; Tixier-Boichard, M.; Kämpe, O.; Andersson, L. The Dark brown plumage color in chickens is caused by an 8.3-kb deletion upstream of SOX10. Pigment Cell Melanoma Res. 2011, 24, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Domyan, E.T.; Guernsey, M.W.; Kronenberg, Z.; Krishnan, S.; Boissy, R.E.; Vickrey, A.I.; Rodgers, C.; Cassidy, P.; Leachman, S.A.; Fondon, J.W.; et al. Epistatic and Combinatorial Effects of Pigmentary Gene Mutations in the Domestic Pigeon. Curr. Biol. 2014, 24, 459–464. [Google Scholar] [CrossRef] [Green Version]
- Bondurand, N.; Moal, F.D.-L.; Stanchina, L.; Collot, N.; Baral, V.; Marlin, S.; Attie-Bitach, T.; Giurgea, I.; Skopinski, L.; Reardon, W.; et al. Deletions at the SOX10 Gene Locus Cause Waardenburg Syndrome Types 2 and 4. Am. J. Hum. Genet. 2007, 81, 1169–1185. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Chen, H.; Jiang, W.; Hu, Z.; Mei, L.; Xue, J.; He, C.; Liu, Y.; Xia, K.; Feng, Y. Novel mutations in the SOX10 gene in the first two Chinese cases of type IV Waardenburg syndrome. Biochem. Biophys. Res. Commun. 2011, 408, 620–624. [Google Scholar] [CrossRef]
- Tachibana, M.; Kobayashi, Y.; Matsushima, Y. Mouse Models for Four Types of Waardenburg Syndrome. Pigment. Cell Res. 2003, 16, 448–454. [Google Scholar] [CrossRef]
- Antonellis, A.; Bennett, B.; Menheniott, T.R.; Prasad, A.B.; Lee-Lin, S.-Q.; Green, E.D.; Paisley, D.; Kelsh, R.; Pavan, W.; Ward, A. Deletion of long-range sequences at Sox10 compromises developmental expression in a mouse model of Waardenburg-Shah (WS4) syndrome. Hum. Mol. Genet. 2006, 15, 259–271. [Google Scholar] [CrossRef]
- Aoki, Y.; Saint-Germain, N.; Gyda, M.; Magner-Fink, E.; Lee, Y.-H.; Credidio, C.; Saint-Jeannet, J.-P. Sox10 regulates the development of neural crest-derived melanocytes in Xenopus. Dev. Biol. 2003, 259, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, A.; Hochedlinger, K. The Sox Family of Transcription Factors: Versatile Regulators of Stem and Progenitor Cell Fate. Cell Stem Cell 2013, 12, 15–30. [Google Scholar] [CrossRef] [Green Version]
- De La Rocha, A.M.A.; Sampron, N.; Alonso, M.M.; Matheu, A. Role of SOX family of transcription factors in central nervous system tumors. Am. J. Cancer Res. 2014, 4, 312–324. [Google Scholar]
- Kellerer, S.; Schreiner, S.; Stolt, C.C.; Scholz, S.; Bosl, M.R.; Wegner, M. Replacement of the Sox10 transcription factor by Sox8 reveals incomplete functional equivalence. Development 2006, 133, 2875–2886. [Google Scholar] [CrossRef]
- Dravis, C.; Spike, B.T.; Harrell, J.C.; Johns, C.; Trejo, C.L.; Southard-Smith, E.M.; Perou, C.M.; Wahl, G.M. Sox10 Regulates Stem/Progenitor and Mesenchymal Cell States in Mammary Epithelial Cells. Cell Rep. 2015, 12, 2035–2048. [Google Scholar] [CrossRef] [Green Version]
- Pingault, V.; Bodereau, V.; Baral, V.; Marcos, S.; Watanabe, Y.; Chaoui, A.; Fouveaut, C.; Leroy, C.; Vérier-Mine, O.; Francannet, C.; et al. Loss-of-Function Mutations in SOX10 Cause Kallmann Syndrome with Deafness. Am. J. Hum. Genet. 2013, 92, 707–724. [Google Scholar] [CrossRef] [Green Version]
- Polanco, J.C.; Wilhelm, D.; Davidson, T.-L.; Knight, D.; Koopman, P. Sox10 gain-of-function causes XX sex reversal in mice: Implications for human 22q-linked disorders of sex development. Hum. Mol. Genet. 2009, 19, 506–516. [Google Scholar] [CrossRef] [Green Version]
- Mertelmeyer, S.; Weider, M.; Baroti, T.; Reiprich, S.; Fröb, F.; Stolt, C.C.; Wagner, K.-U.; Wegner, M. The transcription factor Sox10 is an essential determinant of branching morphogenesis and involution in the mouse mammary gland. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef]
- Sadaf, T.; Rashid, M.; Hussain, A.; Mahmud, A.; Bukhari, S.M.; Noor, R.; Mustafa, G.; Saleem, M.; Ali, A.; Ashraf, S.; et al. Interspecific variations in external and internal egg quality among various captive avian species from Punjab, Pakistan. Braz. J. Biol. 2022, 82, 1–6. [Google Scholar] [CrossRef]
- Purcell, S. PLINK (1.07). Documentation. Book. 2010, pp. 1–293. Available online: http://zzz.bwh.harvard.edu/plink/dist/plink-doc-1.07.pdf (accessed on 28 August 2010).
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Roulin, A.; Ducrest, A.-L. Genetics of colouration in birds. Semin. Cell Dev. Biol. 2013, 24, 594–608. [Google Scholar] [CrossRef]
- Brewer, M.H.; Ma, K.H.; Beecham, G.W.; Gopinath, C.; Baas, F.; Choi, B.-O.; Reilly, M.M.; Shy, M.E.; Züchner, S.; Svaren, J.; et al. Haplotype-specific modulation of a SOX10/CREB response element at the Charcot–Marie–Tooth disease type 4C locus SH3TC2. Hum. Mol. Genet. 2014, 23, 5171–5187. [Google Scholar] [CrossRef] [Green Version]
- Inoue, K.; Khajavi, M.; Ohyama, T.; Hirabayashi, S.-I.; Wilson, J.H.; Reggin, J.D.; Mancias, P.; Butler, I.J.; Wilkinson, M.F.; Wegner, M.; et al. Molecular mechanism for distinct neurological phenotypes conveyed by allelic truncating mutations. Nat. Genet. 2004, 36, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Cherry, P.; Morris, T.R. Domestic Duck Production: Science and Practice; CABI Publishing: Wallingford, UK, 2008. [Google Scholar]
- Wright, D.; Rubin, C.; Schutz, K.; Kerje, S.; Kindmark, A.; Brandström, H.; Andersson, L.; Pizzari, T.; Jensen, P. Onset of Sexual Maturity in Female Chickens is Genetically Linked to Loci Associated with Fecundity and a Sexual Ornament. Reprod. Domest. Anim. 2011, 47, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.V.; Panaccione, A.; Nonaka, D.; Prasad, M.L.; Boyd, K.L.; Brown, B.; Guo, Y.; Sewell, A.; Yarbrough, W.G. Diagnostic SOX10 gene signatures in salivary adenoid cystic and breast basal-like carcinomas. Br. J. Cancer 2013, 109, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-R.; Kim, T.-H.; Choi, K.-C. Functions and physiological roles of two types of estrogen receptors, ERα and ERβ, identified by estrogen receptor knockout mouse. Lab. Anim. Res. 2012, 28, 71. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.E.; Fuller, P. The importance of ERβ signalling in the ovary. J. Endocrinol. 2010, 205, 15–23. [Google Scholar] [CrossRef]
- González-Morán, M.G. Changes in the cellular localization of estrogen receptor alpha in the growing and regressing ovaries of Gallus domesticus during development. Biochem. Biophys. Res. Commun. 2014, 447, 197–204. [Google Scholar] [CrossRef]
- Cankaya, S.; Ocak, N.; Sungu, M. Canonical Correlation Analysis for Estimation of Relationships between Sexual Maturity and Egg Production Traits upon Availability of Nutrients in Pullets. Asian-Australasian J. Anim. Sci. 2008, 21, 1576–1584. [Google Scholar] [CrossRef]
- Field, A. Discovering Statistics Using SPSS ISM (London, England) Introducing Statistical Methods Series, 3rd ed.; SAGE Publications Ltd.: Thousand Oaks, CA, USA, 2009; Volume 2. [Google Scholar]
- Sauna, Z.E.; Kimchi-Sarfaty, C. Understanding the contribution of synonymous mutations to human disease. Nat. Rev. Genet. 2011, 12, 683–691. [Google Scholar] [CrossRef]
- Kimchi-Sarfaty, C.; Oh, J.M.; Kim, I.-W.; Sauna, Z.E.; Calcagno, A.M.; Ambudkar, S.V.; Gottesman, M.M. A “Silent” Polymorphism in the MDR1 Gene Changes Substrate Specificity. Science 2007, 315, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Komar, A.A. Silent SNPs: Impact on gene function and phenotype. Pharmacogenomics 2007, 8, 1075–1080. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.-M.; Huang, J.; Chen, S.-Y.; Liu, Y.-P. Mutations of TYR and MITF Genes are Associated with Plumage Colour Phenotypes in Geese. Asian-Australasian J. Anim. Sci. 2014, 27, 778–783. [Google Scholar] [CrossRef]




| Primer Name | Primer Sequence (5′ to 3′) | Product Size (bp) | Tm (°C) |
|---|---|---|---|
| S10-E1-F | ATTAGTAAAAACCAAGCCTC | 242 | 53 |
| S10-E1-R | CTTGTTACTTCCATTGACCC | ||
| S10-E2-F | ACCACCACTGCCTCTCGCC | 225 | 62 |
| S10-E2-R | TCTTGTTACTTCCATTGACCCG | ||
| S10-E3-F1 | TTTCTCACACACCTGCCC | 330 | 59 |
| S10-E3-R1 | TCACCTGAGGAGTGTTCTG | ||
| S10-E3-F2 | GCTGCTGAACGAAAGCGACA | 289 | 63 |
| S10-E3-R2 | GGTTCACAAAGACCCAGGACTC | ||
| S10-E4-F | CTCCAAAGCCCAGGTGAA | 245 | 59 |
| S10-E4-R | ATGGCAGTGTAAAGAGGACG |
| SOX10 SNP | Mutation | Location | Amino Acid Change |
|---|---|---|---|
| g. 54065311 | G>A | Exon 2 | Ser31Ser |
| g. 54065419 | C>T | Exon 2 | Arg67Arg |
| g. 54070844 | C>T | Exon 3 | His162His |
| g. 54070853 | C>T | Exon 3 | Tyr165Tyr |
| g. 54070904 * | C>T | Exon 3 | Gly162Gly |
| g. 54070940 * | C>T | Exon 3 | Gly191Gly |
| g. 54071020 * | C>T | Exon 3 | Pro195Leu |
| g. 54071499 * | A>C | Exon 3 | Gln222Lys |
| g. 54071948 | C>T | Exon 4 | Pro248Leu |
| g. 54072026 * | G>T | Exon 4 | Leu398Val |
| g. 54072059 * | A>C | Exon 4 | Pro423Thr |
| Parameter | Means of Haplotypes of Reproductive Traits and Egg Quality [g. 54065419 | g. 54070844 | g. 54070904] | p-Value | ||||
|---|---|---|---|---|---|---|
| CTT (H1) | CCT (H2) | CTC (H3) | TCC (H4) | CCC (H5) | ||
| Haplotype Count (Frequency) | 157 (0.1221) | 21 (0.0162) | 150 (0.1163) | 391 (0.3043) | 554 (0.4310) | |
| Age at sexual maturity | 121.38 ± 1.79 | 116.50 ± 3.61 | 120.58 ± 1.71 | 121.49 ± 0.86 | 121.36 ± 0.64 | 0.93 |
| Egg weight at sexual maturity | 48.54 ± 1.11 | 50.50 ± 2.25 | 45.32 ± 0.89 | 46.25 ± 0.57 | 46.8 ± 0.41 | 0.18 |
| Egg production at 150 days | 22.08 ± 1.22 | 26.83 ± 2.66 | 21.87 ± 1.24 | 22.12 ± 0.70 | 22.66 ± 0.48 | 0.80 |
| Egg production at 240 days | 99.24 ± 2.32 | 105.17 ± 7.81 | 100.20 ± 2.05 | 99.73 ± 1.22 | 102.66 ± 0.75 | 0.18 |
| Egg production at 270 days | 124.68 ± 2.70 ab | 130.83 ± 4.56 a | 125.63 ± 2.23 ab | 123.91 ± 1.42 b | 128.35 ± 0.82 ab | 0.03 |
| Egg production at 300 days | 150.42 ± 2.7 AB | 158.33 ± 4.75 A | 152.87 ± 2.36 AB | 149.01 ± 1.67 B | 155.12 ± 0.90 AB | 0.01 |
| Egg production at 330 days | 175.99 ± 3.00 AB | 182.50 ± 7.94 A | 180.26 ± 2.52 AB | 174.69 ± 1.91 B | 181.57 ± 1.01 AB | 0.01 |
| Egg production at 360 days | 201.77 ± 3.35 AB | 210.33 ± 8.95 A | 207.24 ± 2.69 AB | 199.49 ± 2.17 B | 206.72 ± 1.18 AB | 0.01 |
| Egg production at 390 days | 225.84 ± 3.69 ab | 239.00 ± 8.97 a | 232.60 ± 2.89 ab | 223.45 ± 2.47 b | 231.36 ± 1.35 ab | 0.02 |
| Egg production at 420 days | 248.28 ± 4.08 ab | 263.67 ± 8.67 a | 256.41 ± 3.22 ab | 247.19 ± 2.77 b | 255.05 ± 1.54 ab | 0.02 |
| Haugh unit (HU) | 76.86 ± 1.78 | 78.12 ± 2.28 | 79.34 ± 1.50 | 80.90 ± 0.86 | 80.94 ± 0.56 | 0.17 |
| SNPs | Genotype | Genotypic Frequency | MAF | χ2 | p-Value | ||
|---|---|---|---|---|---|---|---|
| White | Grey | Black | |||||
| C/C | 0.12 | 0.14 | 0.15 | ||||
| g.54065419 | C/T | 0.16 | 0.26 | 0.08 | 0.34 | 50.58 | 0.000 ** |
| T/T | 0.03 | 0.05 | 0.02 | ||||
| C/C | 0.30 | 0.27 | 0.16 | ||||
| g.54070844 | C/T | 0.08 | 0.09 | 0.10 | 0.15 | 18.88 | 0.001 ** |
| T/T | 0.003 | 0.004 | 0.004 | ||||
| C/C | 0.18 | 0.24 | 0.12 | ||||
| g.54070904 | C/T | 0.12 | 0.18 | 0.11 | 0.25 | 5.17 | 0.083 |
| T/T | 0.01 | 0.02 | 0.02 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarjana, T.A.; Zhang, G. Association between Synonymous SNPs of SOX10 and Plumage Color and Reproductive Traits of Ducks. Animals 2022, 12, 3345. https://doi.org/10.3390/ani12233345
Sarjana TA, Zhang G. Association between Synonymous SNPs of SOX10 and Plumage Color and Reproductive Traits of Ducks. Animals. 2022; 12(23):3345. https://doi.org/10.3390/ani12233345
Chicago/Turabian StyleSarjana, Teysar Adi, and Gongyan Zhang. 2022. "Association between Synonymous SNPs of SOX10 and Plumage Color and Reproductive Traits of Ducks" Animals 12, no. 23: 3345. https://doi.org/10.3390/ani12233345