
Direct–Maternal Genetic Parameters for Litter Size and Body Weight of Piglets of a New Black Breed for the Taiwan Black Hog Market

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
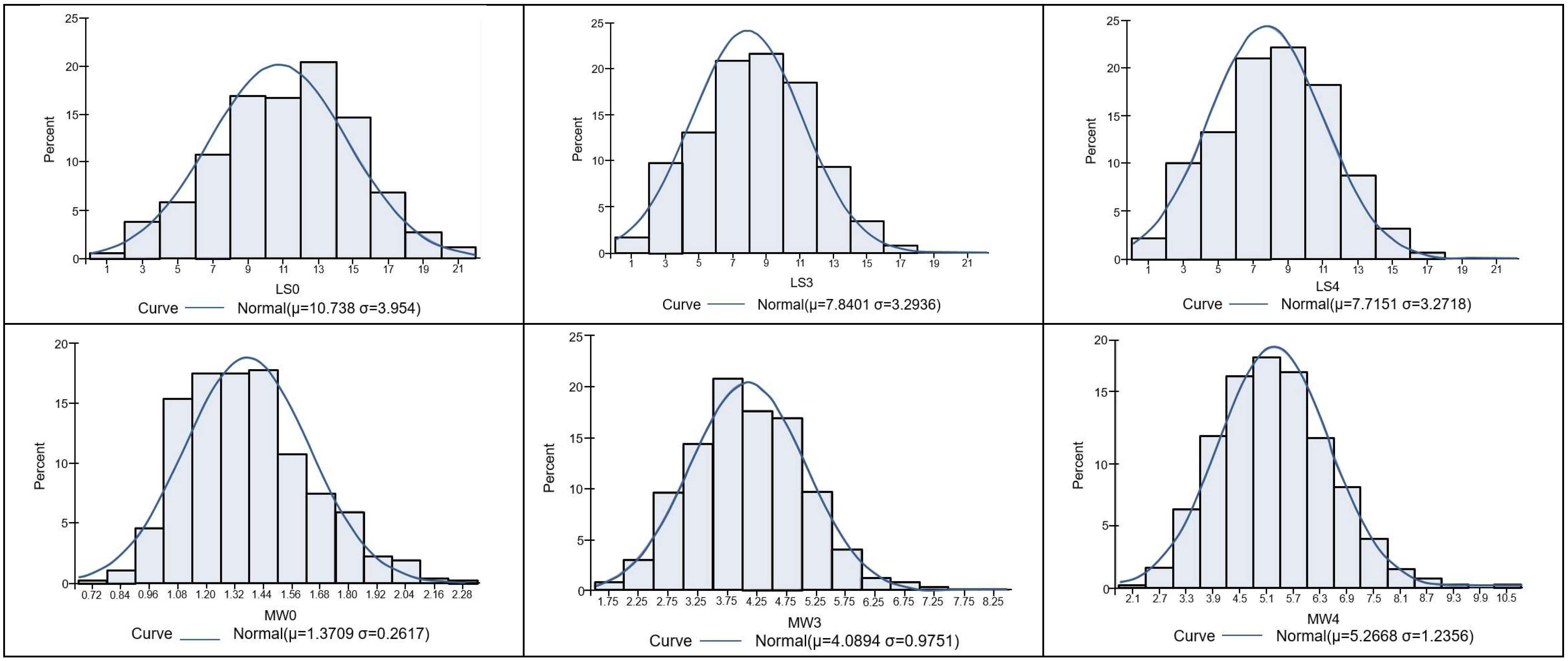
3.1. Descriptive Statistics
3.2. Genetic Parameter Estimates
4. Discussion
4.1. Descriptive Statistics
4.2. Genetic Parameter Estimates
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koch, R.M.; Clark, R.T. Genetic and environmental relationships among economic characters in beef cattle III. Evaluating maternal environment. J. Anim. Sci. 1955, 14, 979–996. [Google Scholar] [CrossRef]
- Meyer, K. Variance components due to direct and maternal effects for growth traits of Australian beef cattle. Liv. Prod. Sci. 1992, 31, 179–204. [Google Scholar] [CrossRef]
- Khombe, C.T.; Hayes, J.F.; Cue, R.I.; Wade, K.M. Estimation of direct additive and maternal additive genetic effects for weaning weight in Mashona cattle of Zimbabwe using an individual animal model. Anim. Sci. 1995, 60, 41–48. [Google Scholar] [CrossRef]
- Robinson, D.L. Estimation and interpretation of direct and maternal genetic parameters for weights of Australian Angus cattle. Livest. Prod. Sci. 1996, 45, 1–11. [Google Scholar] [CrossRef]
- Roehe, R.; Kennedy, B.W. The influence of maternal effects on accuracy of evaluation of litter size in swine. J. Anim. Sci. 1993, 71, 2353–2364. [Google Scholar] [CrossRef] [PubMed]
- Southwood, O.I.; Kennedy, B.W. Estimation of direct and maternal genetic variance for litter size in Canadian, Yorkshire and Landrace pigs using an animal model. J. Anim. Sci. 1990, 68, 1841–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, J.T.; Crump, R.E. Genetic parameter estimates for reproduction traits in purebred Landrace pigs. In Proceedings of the 4th World Congress on Genetics Applied to Livestock Production, Edinburgh, UK, 23–27 July 1990. [Google Scholar]
- Perez-Enciso, M.; Gianola, D. Estimates of genetic parameters for litter size in six strains of Iberian pigs. Livest. Prod. Sci. 1992, 32, 283–293. [Google Scholar] [CrossRef]
- Eaglen, S.A.E.; Bijma, P. Genetic parameters of direct and maternal effects for calving ease in Dutch Holstein-Friesian cattle. J. Dairy Sci. 2009, 92, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- SAS Institute Inc. The SAS System for Windows; Release 9.4.; SAS Institute: Cary, NC, USA, 2015. [Google Scholar]
- Groeneveld, E.; Kovac, M.; Mielenz, N. VCE User’s Guide and Reference Manual Version 6.0. Institute of Farm Animal Genetics, Neustadt, Germany. 2010. Available online: ftp://ftp.tzv.fal.de/pub/latest_vce/doc/vce6-manual-3.1-A4.pdf (accessed on 31 August 2015).
- Dickerson, G.E. Composition of hog carcasses as influenced by heritable differences in rate and economy of gain. Iowa Agric. Exp. Stn. Res. Bull. 1947, 28, 489–524. [Google Scholar]
- Willham, R.L. The role of maternal effects in animal breeding: III. Biometrical aspects of maternal effects in animals. J. Anim. Sci. 1972, 35, 1288–1293. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, Z.; Yang, Y.; Zhao, J.; Chen, Q.; Liao, R.; Chen, Z.; Zhang, X.; Xue, M.; Yang, H.; et al. Identification of pleiotropic genes and gene sets underlying growth and immunity traits: A case study on Meishan pigs. Animal 2016, 10, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Tan, C.; Cai, G.; Li, Y.; Wu, D.; Wu, Z. Estimates of variance components and heritability using different animal models for growth, backfat, litter size, and healthy birth ratio in Large White pigs. Can. J. Anim. Sci. 2020, 100, 330–336. [Google Scholar] [CrossRef]
- Arango, J.; Misztal, I.; Tsuruta, S.; Culbertson, M.; Holl, J.W.; Herring, W. Genetic study of individual preweaning mortality and birth weight in Large White piglets using threshold–linear models. Livest. Prod. Sci. 2006, 101, 208–218. [Google Scholar] [CrossRef]
- Fix, J.S.; Cassady, J.P.; Holl, J.W.; Herring, W.O.; Culbertson, M.S.; See, M.T. Effect of piglet birth weight on survival and quality of commercial market swine. Livest. Sci. 2010, 132, 98–106. [Google Scholar] [CrossRef]
- Krahn, G.T. Comparison of Piglet Birth Weight Classes, Parity of the Dam, Number Born Alive and the Relationship with Litter Variation and Piglet Survival until Weaning. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2015. [Google Scholar] [CrossRef] [Green Version]
- Feldpausch, J.A.; Jourquin, J.; Bergstrom, J.R.; Bargen, J.L.; Bokenkroger, C.D.; Davis, D.L.; Gonzalez, J.M.; Nelssen, J.L.; Puls, C.L.; Trout, W.E.; et al. Birth weight threshold for identifying piglets at risk for preweaning mortality. Transl. Anim. Sci. 2019, 3, 633–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bidanel, J.P. Biology and genetics of reproduction. In The Genetics of the Pig, 2nd ed.; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Wallingford, UK, 2011; pp. 218–241. [Google Scholar] [CrossRef] [Green Version]
- Banville, M.; Riquet, J.; Bahon, D.; Sourdioux, M.; Canario, L. Genetic parameters for litter size, piglet growth and sow’s early growth and body composition in the Chinese–European line Tai Zumu. J. Anim. Breed. Genet. 2015, 132, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Estany, J.; Sorensen, D. Estimation of genetic parameters for litter size in Danish Landrace and Yorkshire pigs. Anim. Sci. 1995, 60, 315–324. [Google Scholar] [CrossRef]
- Roehe, R.; Kennedy, B.W. Estimation of genetic parameters for litter size in Canadian Yorkshire and Landrace swine with each parity of farrowing treated as a different trait. J. Anim. Sci. 1995, 73, 2959–2970. [Google Scholar] [CrossRef] [Green Version]
- Su, G.; Lund, M.S.; Sorensen, D. Selection for litter size at day five to improve litter size at weaning and piglet survival rate. J. Anim. Sci. 2007, 85, 1385–1392. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.; Chang, H.-K.; Lee, S.-S.; Choi, T.-J. Genetic analysis of major production and reproduction traits of Korean Duroc, Landrace and Yorkshire pigs. Animals 2021, 11, 1321. [Google Scholar] [CrossRef] [PubMed]
- Cheverud, J.M. A comparison of genetic and phenotypic correlations. Evolution 1988, 42, 958–968. [Google Scholar] [CrossRef]
- Huby, M.; Canario, L.; Maignel, L.; Tribout, T.; Gogué, J.; Bidanel, J.P. Genetic correlations between litter size and weights, piglet weight variability and piglet survival from birth to weaning in Large White pigs. In Proceedings of the 54th Annual Meeting of the European Association for Animal Production, Rome, Italy, 31 August–3 September 2003. [Google Scholar]
- Kaufmann, D.; Hofer, A.; Bidanel, J.P.; Künzi, N. Genetic parameters for individual birth and weaning weight and for litter size of Large White pigs. J. Anim. Breed. Genet. 2000, 117, 121–128. [Google Scholar] [CrossRef]
- Alves, K.; Schenkel, F.S.; Brito, L.F.; Robinson, J.A.B. Estimation of direct and maternal genetic parameters for individual birth weight and probe weight using cross-fostering information. Can. J. Anim. Sci. 2018, 98, 548–556. [Google Scholar] [CrossRef]
- Alves, K.; Schenkel, F.S.; Brito, L.F.; Robinson, A. Estimation of direct and maternal genetic parameters for individual birth weight, weaning weight, and probe weight in Yorkshire and Landrace pigs. J. Anim. Sci. 2018, 96, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Roehe, R.; Shrestha, N.P.; Mekkawy, W.; Baxter, E.M.; Knap, P.W.; Smurthwaite, K.M.; Jarvis, S.; Lawrence, A.B.; Edwards, S.A. Genetic parameters of piglet survival and birth weight from a two-generation crossbreeding experiment under outdoor conditions designed to disentangle direct and maternal effects. J. Anim. Sci. 2010, 88, 1276–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knol, E.F.; Ducro, B.J.; Van Arendonk, J.A.M.; Van Der Lende, T. Direct, maternal and nurse sow genetic effects on farrowing-, pre-weaning- and total piglet survival. Livest. Prod. Sci. 2002, 73, 153–164. [Google Scholar] [CrossRef]
- Wilson, A.J.; Coltman, D.W.; Pemberton, J.M.; Overall, A.D.J.; Byrne, K.A.; Kruuk, L.E.B. Maternal genetic effects set the potential for evolution in a free-living vertebrate population. J. Evol. Biol. 2005, 18, 405–414. [Google Scholar] [CrossRef]
- Ghafouri-Kesbi, F.; Baneh, H. Genetic parameters for direct and maternal effects on growth traits of sheep. Archiv. Tierzucht. 2012, 55, 603–611. [Google Scholar] [CrossRef]
- Dufrasne, M.; Wavreille, J.; Piedboeuf, M.; Gengler, N. Genetic parameters for individual birth weight, weaning weight and final weight of crossbred pigs from Pietrain boars. In Proceedings of the 10th World Congress on Genetics Applied to Livestock Production, Vancouver, BC, Canada, 17–22 January 2014. [Google Scholar]
- Ahlschwede, W.T.; Robison, O.W. Maternal effects on weights and backfat of swine. J. Anim. Sci. 1971, 33, 1206–1211. [Google Scholar] [CrossRef]
- Kushwaha, B.P.; Mandal, A.; Arora, A.L.; Kumar, R.; Kumar, S.; Notter, D.R. Direct and maternal (co)variance components and heritability estimates for body weights in Chokla sheep. J. Anim. Breed. Genet. 2009, 126, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Garrick, D.J.; Pollak, E.J.; Quaas, R.L.; Vleck, L.D.V. Variance heterogeneity in direct and maternal weight traits by sex and percent purebred for Simmental-sired calves. J. Anim. Sci. 1989, 67, 2515–2528. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Item | Litter size at, head | Average piglet weight at, kg | ||||
| Birth | 3-week | 4-week | Birth | 3-week | 4-week | |
| Minimum | 1 | 1 | 1 | 0.74 | 1.81 | 2.24 |
| Maximum | 21 | 17 | 17 | 2.31 | 8.31 | 10.51 |
| Mean | 10.74 | 7.84 | 7.72 | 1.37 | 4.09 | 5.27 |
| SD | 3.95 | 3.29 | 3.27 | 0.26 | 0.98 | 1.24 |
| CV | 36.8% | 42.0% | 42.4% | 19.0% | 24.0% | 23.5% |
| Skewness | −0.113 | 0.027 | 0.011 | 0.522 | 0.380 | 0.400 |
| Kurtosis | −0.347 | −0.447 | −0.467 | −0.006 | 0.424 | 0.278 |
| Litter size at | Birth | 3-week old | 4-week old |
| Birth | 0.236 ± 0.045 (0.230 ± 0.047) | 0.797 ± 0.069 | 0.790 ± 0.072 |
| 3-week old | 0.662 ± 0.029 | 0.253 ± 0.049 (0.250 ± 0.053) | 0.998 ± 0.000 |
| 4-week old | 0.643 ± 0.029 | 0.988 ± 0.006 | 0.247 ± 0.049 (0.247 ± 0.052) |
| Body weight at | Birth | 3-week old | 4-week old |
| Birth | 0.220 ± 0.037 (0.221 ± 0.049) | 0.567 ± 0.136 | 0.426 ± 0.144 |
| 3-week old | 0.486 ± 0.033 | 0.148 ± 0.044 (0.146 ± 0.046) | 0.976 ± 0.018 |
| 4-week old | 0.410 ± 0.035 | 0.881 ± 0.018 | 0.194 ± 0.044 (0.198 ± 0.051) |
| Trait | h2d | h2m | rdm | h2r | h2T |
|---|---|---|---|---|---|
| LS0 | 0.108 ± 0.079 | 0.237 * ± 0.100 | −0.757 ± 0.460 | 0.045 (0.066) | 0.103 (0.114) |
| LS3 | 0.181 * ± 0.083 | 0.225 ± 0.113 | −0.814 * ± 0.316 | 0.047 (0.038) | 0.077 (0.076) |
| LS4 | 0.174 * ± 0.073 | 0.222 ± 0.134 | −0.791 * ± 0.340 | 0.052 (0.040) | 0.085 (0.079) |
| MW0 | 0.080 ± 0.084 | 0.252 * ± 0.085 | −0.745 ± 0.593 | 0.047 (0.065) | 0.121 (0.125) |
| MW3 | 0.085 ± 0.079 | 0.115 ± 0.090 | −0.484 ± 0.551 | 0.071 (0.089) | 0.104 (0.116) |
| MW4 | 0.078 ± 0.069 | 0.125 ± 0.069 | −0.022 ± 0.675 | 0.137 (0.140) | 0.199 (0.195) |
| Item | Direct Genetic Effects | Maternal Genetic Effects | ||||
|---|---|---|---|---|---|---|
| LS0d | LS3d | LS4d | LS0m | LS3m | LS4m | |
| LS0d | 0.139 * ± 0.049 | 0.939 * ± 0.071 | 0.924 * ± 0.080 | −0.696 * ± 0.165 | −0.897 * ± 0.105 | −0.895 * ± 0.109 |
| LS3d | 0.170 * ± 0.036 | 0.999 * ± 0.001 | −0.801 * ± 0.154 | −0.830 * ± 0.109 | −0.831 * ± 0.112 | |
| LS4d | 0.162 * ± 0.036 | −0.808 * ± 0.143 | −0.816 * ± 0.110 | −0.817 * ± 0.111 | ||
| LS0m | 0.216 * ± 0.051 | 0.862 * ± 0.095 | 0.867 * ± 0.095 | |||
| LS3m | 0.244 * ± 0.063 | 1.000 * ± 0.000 | ||||
| LS4m | 0.238 * ± 0.061 | |||||
| Trait | Direct Genetic | Maternal Genetic | ||||
|---|---|---|---|---|---|---|
| MW0d | MW3d | MW4d | MW0m | MW3m | MW4m | |
| MW0d | 0.105 ± 0.066 | 0.926 * ± 0.184 | 0.874 * ± 0.260 | −0.660 * ± 0.379 | −0.682 * ± 0.221 | −0.402 ± 0.298 |
| MW3d | 0.105 * ± 0.043 | 0.989 * ± 0.029 | −0.500 ± 0.432 | −0.431 ± 0.400 | −0.145 ± 0.454 | |
| MW4d | 0.095 * ± 0.043 | −0.370 ± 0.533 | −0.351 ± 0.463 | −0.084 ± 0.481 | ||
| MW0m | 0.220 * ± 0.074 | 0.565 * ± 0.226 | 0.322 ± 0.261 | |||
| MW3m | 0.098 * ± 0.055 | 0.937 * ± 0.051 | ||||
| MW4m | 0.118 * ± 0.053 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-L.; Lin, M.-Y.; Wang, H.-S.; Hsu, C.-B.; Lin, C.-Y.; Chang, S.-C.; Shen, P.-C.; Chang, H.-L. Direct–Maternal Genetic Parameters for Litter Size and Body Weight of Piglets of a New Black Breed for the Taiwan Black Hog Market. Animals 2022, 12, 3295. https://doi.org/10.3390/ani12233295
Lee H-L, Lin M-Y, Wang H-S, Hsu C-B, Lin C-Y, Chang S-C, Shen P-C, Chang H-L. Direct–Maternal Genetic Parameters for Litter Size and Body Weight of Piglets of a New Black Breed for the Taiwan Black Hog Market. Animals. 2022; 12(23):3295. https://doi.org/10.3390/ani12233295
Chicago/Turabian StyleLee, Hsiu-Lan, Mu-Yao Lin, Han-Sheng Wang, Chin-Bin Hsu, Cheng-Yung Lin, Shen-Chang Chang, Perng-Chih Shen, and Hsiu-Luan Chang. 2022. "Direct–Maternal Genetic Parameters for Litter Size and Body Weight of Piglets of a New Black Breed for the Taiwan Black Hog Market" Animals 12, no. 23: 3295. https://doi.org/10.3390/ani12233295