
Use of Microorganisms as Nutritional and Functional Feedstuffs for Nursery Pigs and Broilers

Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Bacteria

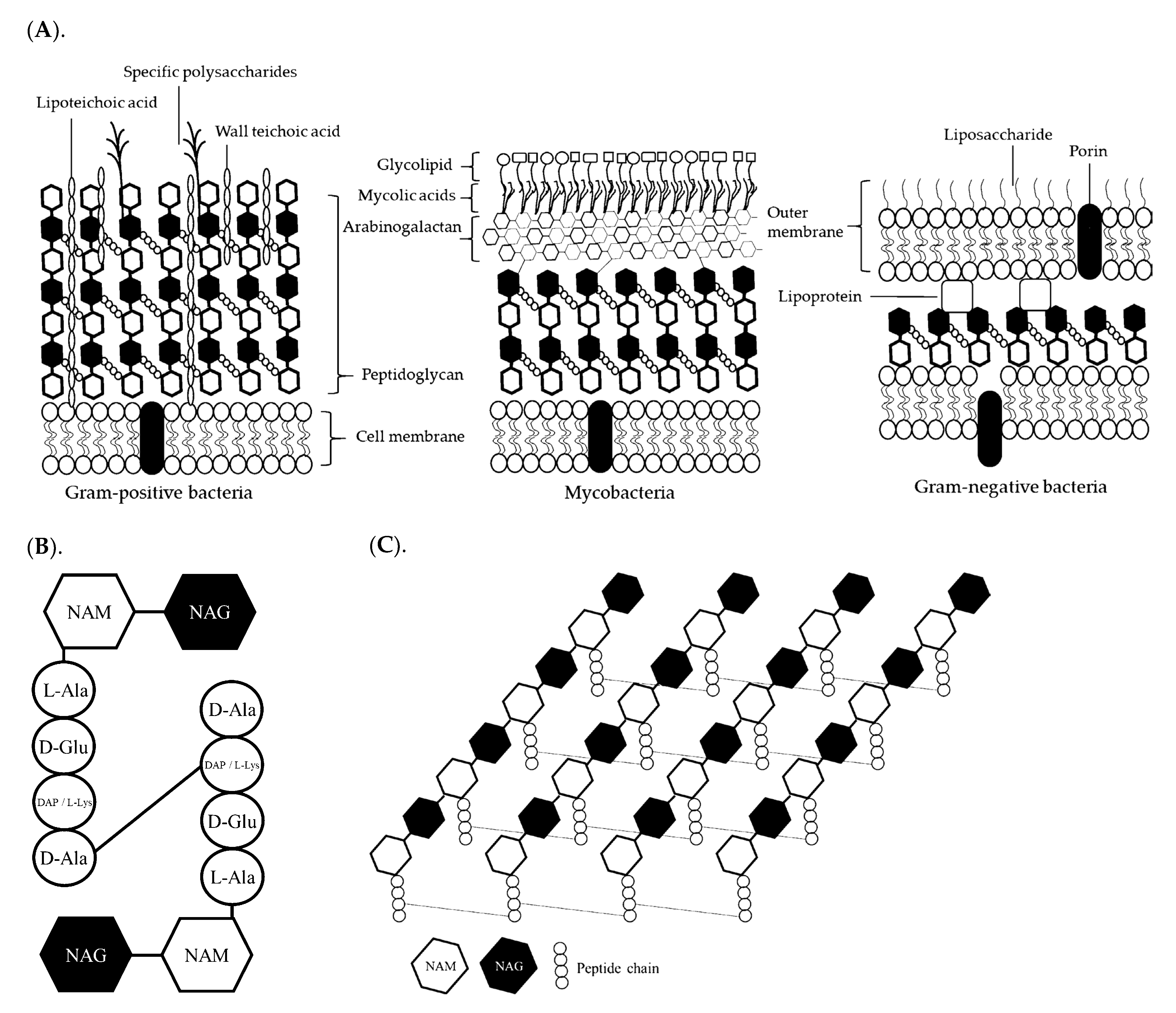{kind=link}
{kind=link}
{kind=link}
| Bacteria | Yeast | Microalgae | ||
|---|---|---|---|---|
| DM 1, % | 90 to 95 | 93 | 94 | |
| CP, % DM basis | 50 to 80 | 12 to 53 | 10 to 70 | |
| Lipid, % DM basis | 7 to 15 | 1 to 40 | 3 to 71 | |
| Total Fiber, % DM basis | 3 to 6 | 2 to 40 | 10 to 66 | |
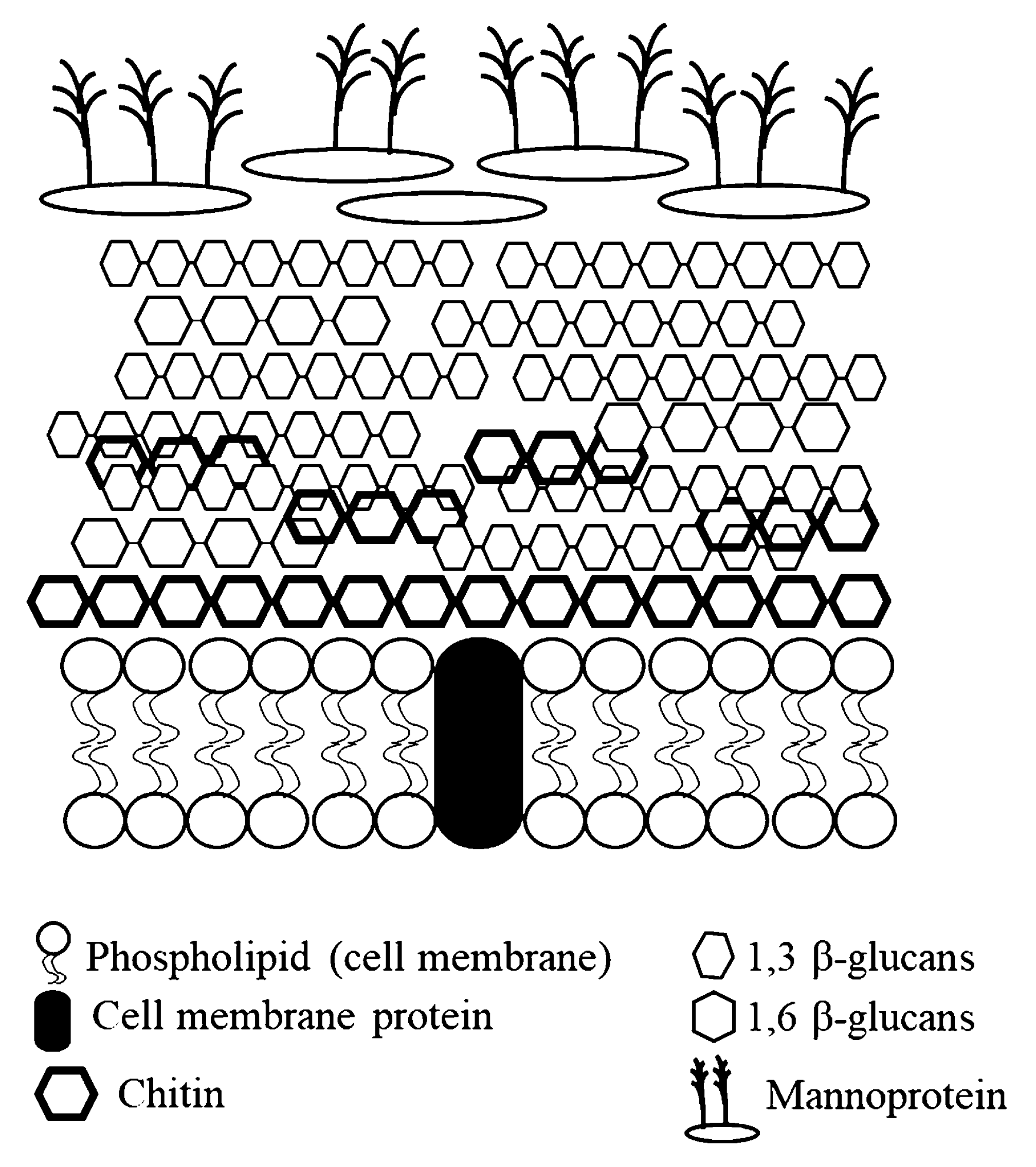
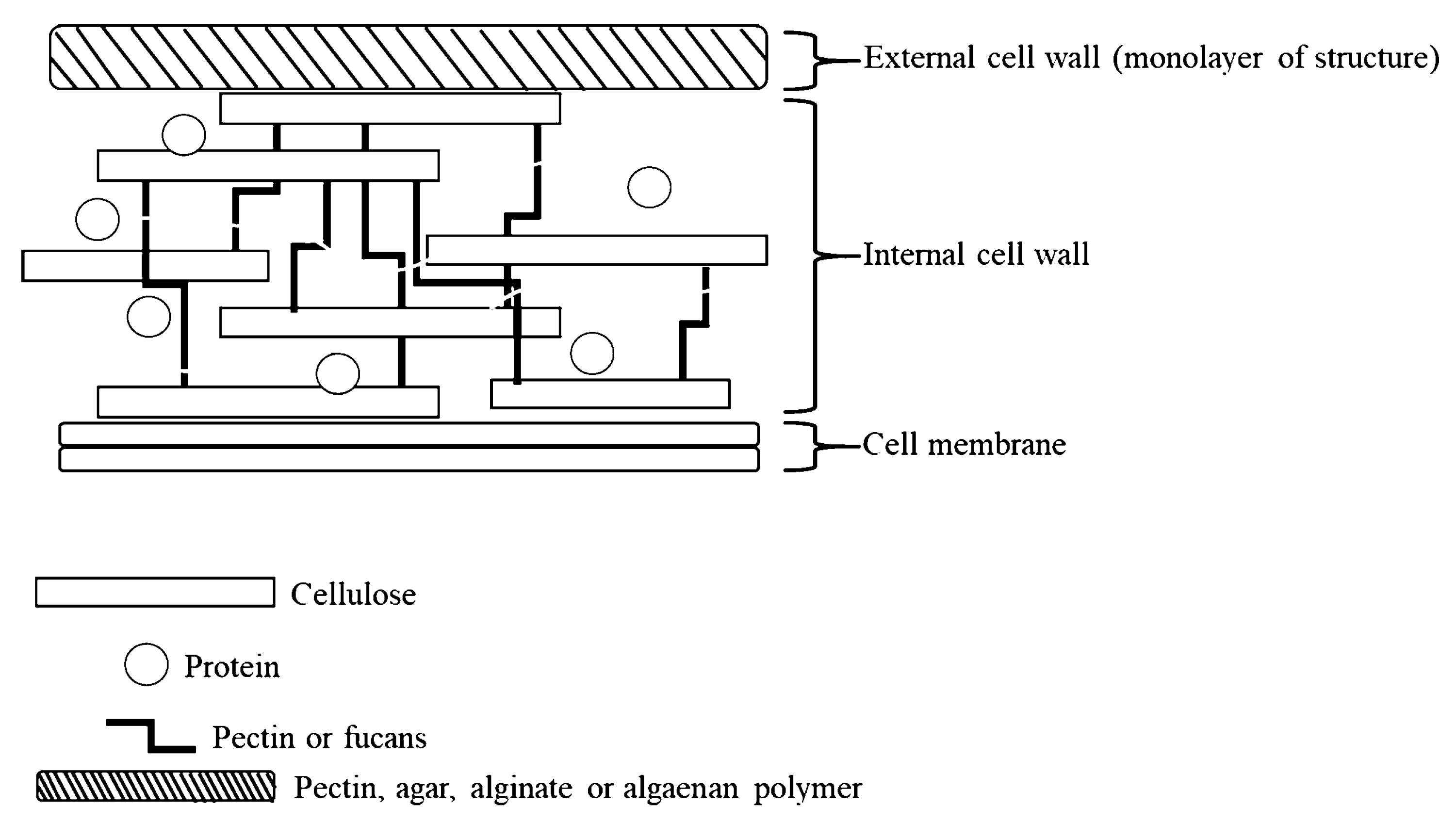
| Cell wall contents | Gram + 2 | Gram − 2 | Mannoprotein (35 to 40%); 1,3 β-glucan (50 to 55%); 1,6 β-glucan (5 to 10%); Chitin (up to 3%) | Polysaccharide (1 to 12%) Soluble protein (up to 4.5%) |
| 20 to 80 nm; PN 3 (40 to 60%); Teichoic acid (up to 40%); Arabinogalactan (10 to 20%) | 8 to 10 nm; PN (10 to 20%); LPS 4; Lipoprotein | |||
| References | [30,31] | [23,32,33] | [34,35,36] | |

1.2. Yeasts
1.3. Microalgae
2. Microorganisms as Functional Feed Additives
2.1. Introduction of Functional Feed Additives
2.2. Mechanism and Application
2.2.1. Bacteria
2.2.2. Yeasts
2.2.3. Microalgae
3. Single Cell Protein (SCP)
3.1. Introduction of SCP
3.2. Application of SCP
3.2.1. Bacteria
3.2.2. Yeasts
3.2.3. Microalgae
4. Single-Cell Oil (SCO)
4.1. Introduction of SCO
4.2. Application of SCO
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pomar, C.; Remus, A. Precision Pig Feeding: A Breakthrough toward Sustainability. Anim. Front. 2019, 9, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.-J.; Lee, C.-H.; Kim, S.W. Aspergillus Oryzae GB-107 Fermentation Improves Nutritional Quality of Food Soybeans and Feed Soybean Meals. J. Med. Food 2004, 7, 430–435. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Duarte, M.E.; Jang, K.B.; Kim, S.W. Soy Protein Concentrate Replacing Animal Protein Supplements and Its Impacts on Intestinal Immune Status, Intestinal Oxidative Stress Status, Nutrient Digestibility, Mucosa-Associated Microbiota, and Growth Performance of Nursery Pigs. J. Anim. Sci. 2022, 100, 1–76. [Google Scholar] [CrossRef]
- Kim, S.W. Bio-Fermentation Technology to Improve Efficiency of Swine Nutrition. Asian-Australas. J. Anim. Sci. 2010, 23, 825–832. [Google Scholar] [CrossRef]
- Babel, W.; Pöhland, H.-D.; Soyez, K. Single Cell Proteins. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2000; pp. 437–452. ISBN 9781482279047. [Google Scholar]
- Giec, A.; Skupin, J. Single Cell Protein as Food and Feed. Food/Nahrung 1988, 32, 219–229. [Google Scholar] [CrossRef]
- Ugalde, U.O.; Castrillo, J.I. Single Cell Proteins from Fungi and Yeasts. In Applied Mycology and Biotechnology; Elsevier: Amsterdam, The Netherlands, 2002; Volume 2, pp. 123–149. [Google Scholar]
- Rounsevell, M.D.A.; Ewert, F.; Reginster, I.; Leemans, R.; Carter, T.R. Future Scenarios of European Agricultural Land Use. Agric. Ecosyst. Environ. 2005, 107, 117–135. [Google Scholar] [CrossRef]
- Kim, S.W.; Easter, R.A. Nutritional Value of Fish Meals in the Diet for Young Pigs. J. Anim. Sci. 2001, 79, 1829. [Google Scholar] [CrossRef]
- Weaver, A.C.; Campbell, J.M.; Crenshaw, J.D.; Polo, J.; Kim, S.W. Efficacy of Dietary Spray Dried Plasma Protein to Mitigate the Negative Effects on Performance of Pigs Fed Diets with Corn Naturally Contaminated with Multiple Mycotoxins. J. Anim. Sci. 2014, 92, 3878–3886. [Google Scholar] [CrossRef]
- U.S. Department of Agriculture. Agricultural Statistics 2019; Bernan Press: Washington, DC, USA, 2019; ISBN 978-1-64143-466-9.
- Kim, S.W.; Less, J.F.; Wang, L.; Yan, T.; Kiron, V.; Kaushik, S.J.; Lei, X.G. Meeting Global Feed Protein Demand: Challenge, Opportunity, and Strategy. Annu. Rev. Anim. Biosci. 2019, 7, 221–243. [Google Scholar] [CrossRef]
- Kim, S.W.; Hansen, J.A. Diet Formulation and Feeding Programs. In Sustainable Swine Nutrition; Wiley: Oxford, UK, 2013; pp. 215–227. ISBN 9780813805344. [Google Scholar]
- Bharathiraja, B.; Sridharan, S.; Sowmya, V.; Yuvaraj, D.; Praveenkumar, R. Microbial Oil—A Plausible Alternate Resource for Food and Fuel Application. Bioresour. Technol. 2017, 233, 423–432. [Google Scholar] [CrossRef]
- Kwak, W.S.; Kang, J.S. Effect of Feeding Food Waste-Broiler Litter and Bakery by-Product Mixture to Pigs. Bioresour. Technol. 2006, 97, 243–249. [Google Scholar] [CrossRef]
- Almeida, F.N.; Petersen, G.I.; Stein, H.H. Digestibility of Amino Acids in Corn, Corn Coproducts, and Bakery Meal Fed to Growing Pigs. J. Anim. Sci. 2011, 89, 4109–4115. [Google Scholar] [CrossRef]
- Rojas, O.J.; Liu, Y.; Stein, H.H. Phosphorus Digestibility and Concentration of Digestible and Metabolizable Energy in Corn, Corn Coproducts, and Bakery Meal Fed to Growing Pigs. J. Anim. Sci. 2013, 91, 5326–5335. [Google Scholar] [CrossRef] [Green Version]
- Luciano, A.; Espinosa, C.D.; Pinotti, L.; Stein, H.H. Standardized Total Tract Digestibility of Phosphorus in Bakery Meal Fed to Pigs and Effects of Bakery Meal on Growth Performance of Weanling Pigs. Anim. Feed Sci. Technol. 2022, 284, 115148. [Google Scholar] [CrossRef]
- Guo, J.Y.; Phillips, C.E.; Coffey, M.T.; Kim, S.W. Efficacy of a Supplemental Candy Coproduct as an Alternative Carbohydrate Source to Lactose on Growth Performance of Newly Weaned Pigs in a Commercial Farm Condition. J. Anim. Sci. 2015, 93, 5304–5312. [Google Scholar] [CrossRef] [Green Version]
- Neumann, C.; Velten, S.; Liebert, F. N Balance Studies Emphasize the Superior Protein Quality of Pig Diets at High Inclusion Level of Algae Meal (Spirulina platensis) or Insect Meal (Hermetia illucens) When Adequate Amino Acid Dupplementation Is Ensured. Animals 2018, 8, 172. [Google Scholar] [CrossRef] [Green Version]
- Allegretti, G.; Talamini, E.; Schmidt, V.; Bogorni, P.C.; Ortega, E. Insect as Feed: An Emergy Assessment of Insect Meal as a Sustainable Protein Source for the Brazilian Poultry Industry. J. Clean. Prod. 2018, 171, 403–412. [Google Scholar] [CrossRef]
- DiGiacomo, K.; Leury, B.J. Review: Insect Meal: A Future Source of Protein Feed for Pigs? Animal 2019, 13, 3022–3030. [Google Scholar] [CrossRef] [Green Version]
- Hogg, S. Essential Microbiology, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2013; ISBN 978-1-119-97890-9. [Google Scholar]
- Johannsen, L. Biological Properties of Bacterial Peptidoglycan. Apmis 1993, 101, 337–344. [Google Scholar] [CrossRef]
- Pazos, M.; Peters, K. Peptidoglycan. In Subcellular Biochemistry; Springer: Berlin/Heidelberg, Germany, 2019; pp. 127–168. [Google Scholar]
- Van Amersfoort, E.S.; Kuiper, J. Receptors, Mediators, and Mechanisms Involved in Bacterial Sepsis and Septic Shock. In Endotoxins; CRC Press: Boca Raton, FL, USA, 2007; pp. 403–426. ISBN 9781420020595. [Google Scholar]
- Duarte, M.E.; Tyus, J.; Kim, S.W. Synbiotic Effects of Enzyme and Probiotics on Intestinal Health and Growth of Newly Weaned Pigs Challenged with Enterotoxigenic F18+Escherichia Coli. Front. Vet. Sci. 2020, 7, 7–573. [Google Scholar] [CrossRef]
- Bandara, T. Alternative Feed Ingredients in Aquaculture: Opportunities and Challenges. J. Entomol. Zool. Stud. 2018, 6, 3087–3094. [Google Scholar]
- Sun, Y.; Duarte, M.E.; Kim, S.W. Dietary Inclusion of Multispecies Probiotics to Reduce the Severity of Post-Weaning Diarrhea Caused by Escherichia Coli F18+ in Pigs. Anim. Nutr. 2021, 7, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G Protein-Coupled Receptors GPR41 and GPR43 Are Activated by Propionate and Other Short Chain Carboxylic Acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukaszczyk, M.; Pradhan, B.; Remaut, H. Bacterial Cell Walls and Membranes; Kuhn, A., Ed.; Subcellular Biochemistry; Springer International Publishing: Cham, Switzerland, 2019; Volume 92, ISBN 978-3-030-18767-5. [Google Scholar]
- Morphology, G. Yeast Cell Architecture and Functions. In Yeast; Wiley: Weinheim, Germany, 2012; pp. 5–24. ISBN 9783527332526. [Google Scholar]
- Pacheco, M.T.B.; Caballero-Cordoba, G.M.; Sgarbieri, V.C. Composition and Nutritive Value of Yeast Biomass and Yeast Protein Concentrates. J. Nutr. Sci. Vitaminol. 1997, 43, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, W.; Richmond, A. Handbook of Microalgal Culture; Richmond, A., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2003; ISBN 9780470995280. [Google Scholar]
- de Jesus Raposo, M.; de Morais, A.; de Morais, R. Emergent Sources of Prebiotics: Seaweeds and Microalgae. Mar. Drugs 2016, 14, 27. [Google Scholar] [CrossRef]
- Kay, R.A.; Barton, L.L. Microalgae as Food and Supplement. Crit. Rev. Food Sci. Nutr. 1991, 30, 555–573. [Google Scholar] [CrossRef]
- Koch, A.L. The Bacteria: Their Origin, Structure, Function and Antibiosis; Springer: Dordrecht, The Netherlands, 2006; ISBN 978-1-4020-6625-2. [Google Scholar]
- Kang, S.-S.; Sim, J.-R.; Yun, C.-H.; Han, S.H. Lipoteichoic Acids as a Major Virulence Factor Causing Inflammatory Responses via Toll-like Receptor. Arch. Pharm. Res. 2016, 39, 1519–1529. [Google Scholar] [CrossRef]
- Stewart, G.G. Energy Metabolism by the Yeast Cell. In Brewing and Distilling Yeasts; Springer International Publishing: Cham, Switzerland, 2017; pp. 77–107. [Google Scholar]
- Stewart, G.G. The Structure and Function of the Yeast Cell Wall, Plasma Membrane and Periplasm. In Brewing and Distilling Yeasts; Springer International Publishing: Cham, Switzerland, 2017; pp. 55–75. ISBN 9783319691244. [Google Scholar]
- Shurson, G.C. Yeast and Yeast Derivatives in Feed Additives and Ingredients: Sources, Characteristics, Animal Responses, and Quantification Methods. Anim. Feed Sci. Technol. 2018, 235, 60–76. [Google Scholar] [CrossRef]
- Kim, S.W.; Holanda, D.M.; Gao, X.; Park, I.; Yiannikouris, A. Efficacy of a Yeast Cell Wall Extract to Mitigate the Effect of Naturally Co-Occurring Mycotoxins Contaminating Feed Ingredients Fed to Young Pigs: Impact on Gut Health, Microbiome, and Growth. Toxins 2019, 11, 633. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.B.; Piao, X.S.; Kim, S.W.; Wang, L.; Liu, P.; Yoon, I.; Zhen, Y.G. Effects of Yeast Culture Supplementation on Growth Performance, Intestinal Health, and Immune Response of Nursery Pigs. J. Anim. Sci. 2009, 87, 2614–2624. [Google Scholar] [CrossRef] [Green Version]
- LeMieux, F.M.; Southern, L.L.; Bidner, T.D. Effect of Mannan Oligosaccharides on Growth Performance of Weanling Pigs1. J. Anim. Sci. 2003, 81, 2482–2487. [Google Scholar] [CrossRef]
- Lipke, P.N.; Ovalle, R. Cell Wall Architecture in Yeast: New Structure and New Challenges. J. Bacteriol. 1998, 180, 3735–3740. [Google Scholar] [CrossRef] [Green Version]
- Kogan, G.; Pajtinka, M.; Babincova, M.; Miadokova, E.; Rauko, P.; Slamenova, D.; Korolenko, T.A. Yeast Cell Wall Polysaccharides as Antioxidants and Antimutagens: Can They Fight Cancer? Neoplasma 2008, 55, 387–393. [Google Scholar]
- Baudelet, P.-H.; Ricochon, G.; Linder, M.; Muniglia, L. A New Insight into Cell Walls of Chlorophyta. Algal Res. 2017, 25, 333–371. [Google Scholar] [CrossRef]
- Da Silva, J.C.; Lombardi, A.T. Chlorophylls in Microalgae: Occurrence, Distribution, and Biosynthesis. In Pigments from Microalgae Handbook; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–18. ISBN 9783030509705. [Google Scholar]
- Bold, H.C.; Wynne, M.J. Introduction to the Algae: Structure and Reproduction, 2nd ed.Prentice-Hall: Englewood Cliffs, NJ, USA, 1985; ISBN 9780134777467. [Google Scholar]
- Metting, F.B. Biodiversity and Application of Microalgae. J. Ind. Microbiol. Biotechnol. 1996, 17, 477–489. [Google Scholar] [CrossRef]
- Austic, R.E.; Mustafa, A.; Jung, B.; Gatrell, S.; Lei, X.G. Potential and Limitation of a New Defatted Diatom Microalgal Biomass in Replacing Soybean Meal and Corn in Diets for Broiler Chickens. J. Agric. Food Chem. 2013, 61, 7341–7348. [Google Scholar] [CrossRef]
- Kim, J.; Magnuson, A.; Tao, L.; Barcus, M.; Lei, X.G. Potential of Combining Flaxseed Oil and Microalgal Biomass in Producing Eggs-Enriched with n − 3 Fatty Acids for Meeting Human Needs. Algal Res. 2016, 17, 31–37. [Google Scholar] [CrossRef]
- Yi, Z.; Xu, M.; Di, X.; Brynjolfsson, S.; Fu, W. Exploring Valuable Lipids in Diatoms. Front. Mar. Sci. 2017, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Velazquez-Lucio, J.; Rodríguez-Jasso, R.M.; Colla, L.M.; Sáenz-Galindo, A.; Cervantes-Cisneros, D.E.; Aguilar, C.N.; Fernandes, B.D.; Ruiz, H.A. Microalgal Biomass Pretreatment for Bioethanol Production: A Review. Biofuel Res. J. 2018, 5, 780–791. [Google Scholar] [CrossRef]
- Pôjo, V.; Tavares, T.; Malcata, F.X. Processing Methodologies of Wet Microalga Biomass toward Oil Separation: An Overview. Molecules 2021, 26, 641. [Google Scholar] [CrossRef]
- Kihlberg, R. The Microbe as a Source of Food. Annu. Rev. Microbiol. 1972, 26, 427–466. [Google Scholar] [CrossRef] [PubMed]
- Bozell, J.J.; Petersen, G.R. Technology Development for the Production of Biobased Products from Biorefinery Carbohydrates—the US Department of Energy’s “Top 10” Revisited. Green Chem. 2010, 12, 539. [Google Scholar] [CrossRef]
- Babson, D.M.; Held, M.; Schmidt-Dannert, C. Designer Microbial Ecosystems—Toward Biosynthesis with Engineered Microbial Consortia. In Natural Products; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2014; pp. 23–38. [Google Scholar]
- Namioka, S.; Sasaki, T.; Maede, Y. Immunopotentiation of the Small Intestine of Weaning Piglets by Peptidoglycan Derived from Bifidobacterium Thermophilum. Bifidobact. Microflora 1991, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kogan, G.; Kocher, A. Role of Yeast Cell Wall Polysaccharides in Pig Nutrition and Health Protection. Livest. Sci. 2007, 109, 161–165. [Google Scholar] [CrossRef]
- Puri, M.; Thyagarajan, T.; Gupta, A.; Barrow, C.J. Omega-3 Fatty Acids Produced from Microalgae. In Hb25_Springer Handbook of Marine Biotechnology; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1043–1057. [Google Scholar]
- Salzman, N.H. Microbiota–Immune System Interaction: An Uneasy Alliance. Curr. Opin. Microbiol. 2011, 14, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Lallès, J.-P.; Bosi, P.; Smidt, H.; Stokes, C.R. Nutritional Management of Gut Health in Pigs around Weaning. Proc. Nutr. Soc. 2007, 66, 260–268. [Google Scholar] [CrossRef]
- Duarte, M.E.; Kim, S.W. Intestinal Microbiota and Its Interaction to Intestinal Health in Nursery Pigs. Anim. Nutr. 2022, 8, 169–184. [Google Scholar] [CrossRef]
- Furbeyre, H.; van Milgen, J.; Mener, T.; Gloaguen, M.; Labussière, E. Effects of Dietary Supplementation with Freshwater Microalgae on Growth Performance, Nutrient Digestibility and Gut Health in Weaned Piglets. Animal 2017, 11, 183–192. [Google Scholar] [CrossRef]
- Iakhno, S.; Umu, Ö.C.O.; Håkenåsen, I.M.; Åkesson, C.P.; Mydland, L.T.; Press, C.M.; Sørum, H.; Øverland, M. Effect of Cyberlindnera Jadinii Yeast as a Protein Source on Intestinal Microbiota and Butyrate Levels in Post-Weaning Piglets. Anim. Microbiome 2020, 2, 13. [Google Scholar] [CrossRef]
- Broom, L.J.; Miller, H.M.; Kerr, K.G.; Knapp, J.S. Effects of Zinc Oxide and Enterococcus Faecium SF68 Dietary Supplementation on the Performance, Intestinal Microbiota and Immune Status of Weaned Piglets. Res. Vet. Sci. 2006, 80, 45–54. [Google Scholar] [CrossRef]
- Monger, X.C.; Gilbert, A.-A.; Saucier, L.; Vincent, A.T. Antibiotic Resistance: From Pig to Meat. Antibiotics 2021, 10, 1209. [Google Scholar] [CrossRef] [PubMed]
- Butaye, P.; Devriese, L.A.; Haesebrouck, F. Antimicrobial Growth Promoters Used in Animal Feed: Effects of Less Well Known Antibiotics on Gram-Positive Bacteria. Clin. Microbiol. Rev. 2003, 16, 175–188. [Google Scholar] [CrossRef]
- Kenny, M.; Smidt, H.; Mengheri, E.; Miller, B. Probiotics—Do They Have a Role in the Pig Industry? Animal 2011, 5, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Teame, T.; Wang, A.; Xie, M.; Zhang, Z.; Yang, Y.; Ding, Q.; Gao, C.; Olsen, R.E.; Ran, C.; Zhou, Z. Paraprobiotics and Postbiotics of Probiotic Lactobacilli, Their Positive Effects on the Host and Action Mechanisms: A Review. Front. Nutr. 2020, 7, 344. [Google Scholar] [CrossRef]
- De Marzi, M.C.; Todone, M.; Ganem, M.B.; Wang, Q.; Mariuzza, R.A.; Fernández, M.M.; Malchiodi, E.L. Peptidoglycan Recognition Protein-Peptidoglycan Complexes Increase Monocyte/Macrophage Activation and Enhance the Inflammatory Response. Immunology 2015, 145, 429–442. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Kang, S.S.; Woo, S.J.; Park, O.J.; Ahn, K.B.; Song, K.D.; Lee, H.K.; Yun, C.H.; Han, S.H. Lipoteichoic Acid of Probiotic Lactobacillus Plantarum Attenuates Poly I: C-Induced IL-8 Production in Porcine Intestinal Epithelial Cells. Front. Microbiol. 2017, 8, 1827. [Google Scholar] [CrossRef]
- Irazoki, O.; Hernandez, S.B.; Cava, F. Peptidoglycan Muropeptides: Release, Perception, and Functions as Signaling Molecules. Front. Microbiol. 2019, 10, 500. [Google Scholar] [CrossRef] [Green Version]
- Sorbara, M.T.; Philpott, D.J. Peptidoglycan: A Critical Activator of the Mammalian Immune System during Infection and Homeostasis. Immunol. Rev. 2011, 243, 40–60. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.; Guidato, P.M.; Peters, K.; Megger, D.A.; Sitek, B.; Classen, B.; Heise, E.M.; Bufe, A. Allergy-Protective Arabinogalactan Modulates Human Dendritic Cells via C-Type Lectins and Inhibition of NF-ΚB. J. Immunol. 2016, 196, 1626–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, A.; Flamminio, G.; Folghera, S.; Peroni, L.; Foresti, I.; Balsari, A.; Turano, A. Expression of Activation Markers on Peripheral-Blood Lymphocytes Following Oral Administration of Bacillus Subtilis Spores. Int. J. Immunopharmacol. 1993, 15, 87–92. [Google Scholar] [CrossRef]
- Poulsen, A.-S.R.; de Jonge, N.; Nielsen, J.L.; Højberg, O.; Lauridsen, C.; Cutting, S.M.; Canibe, N. Impact of Bacillus Spp. Spores and Gentamicin on the Gastrointestinal Microbiota of Suckling and Newly Weaned Piglets. PLoS ONE 2018, 13, e0207382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suda, Y.; Villena, J.; Takahashi, Y.; Hosoya, S.; Tomosada, Y.; Tsukida, K.; Shimazu, T.; Aso, H.; Tohno, M.; Ishida, M.; et al. Immunobiotic Lactobacillus Jensenii as Immune-Health Promoting Factor to Improve Growth Performance and Productivity in Post-Weaning Pigs. BMC Immunol. 2014, 15, 24. [Google Scholar] [CrossRef]
- Taras, D.; Vahjen, W.; Macha, M.; Simon, O. Response of Performance Characteristics and Fecal Consistency to Long-Lasting Dietary Supplementation with the Probiotic Strain Bacillus Cereus Var. Toyoi to Sows and Piglets. Arch. Anim. Nutr. 2005, 59, 405–417. [Google Scholar] [CrossRef]
- He, Y.; Jinno, C.; Kim, K.; Wu, Z.; Tan, B.; Li, X.; Whelan, R.; Liu, Y. Dietary Bacillus Spp. Enhanced Growth and Disease Resistance of Weaned Pigs by Modulating Intestinal Microbiota and Systemic Immunity. J. Anim. Sci. Biotechnol. 2020, 11, 101. [Google Scholar] [CrossRef]
- Pessione, E. Lactic Acid Bacteria Contribution to Gut Microbiota Complexity: Lights and Shadows. Front. Cell. Infect. Microbiol. 2012, 2, 86. [Google Scholar] [CrossRef] [Green Version]
- Taras, D.; Vahjen, W.; Macha, M.; Simon, O. Performance, Diarrhea Incidence, and Occurrence of Escherichia Coli Virulence Genes during Long-Term Administration of a Probiotic Enterococcus Faecium Strain to Sows and Piglets. J. Anim. Sci. 2006, 84, 608–617. [Google Scholar] [CrossRef]
- Pollmann, M.; Nordhoff, M.; Pospischil, A.; Tedin, K.; Wieler, L.H. Effects of a Probiotic Strain of Enterococcus Faecium on the Rate of Natural Chlamydia Infection in Swine. Infect. Immun. 2005, 73, 4346–4353. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, J.N.; Laguna, J.S.; Millán, C.; Casabuena, O.; Gracia, M.I. Effects of a Bacillus-Based Probiotic and Dietary Energy Content on the Performance and Nutrient Digestibility of Wean to Finish Pigs. Anim. Feed Sci. Technol. 2016, 221, 54–61. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Fernández, B.; Viñas, M.; Lizardo, R.; Brufau, J.; Owusu-Asiedu, A.; Walsh, M.C.; Awati, A. Effect of Bacillus Spp. Direct-Fed Microbial on Slurry Characteristics and Gaseous Emissions in Growing Pigs Fed with High Fibre-Based Diets. Animal 2017, 11, 209–218. [Google Scholar] [CrossRef]
- Lee, S.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lokhande, A.; Kim, E.K.; Kwon, I.K.; Kim, Y.H.; Chae, B.J. Effects of Dietary Supplementation with Bacillus Subtilis LS 1–2 Fermentation Biomass on Growth Performance, Nutrient Digestibility, Cecal Microbiota and Intestinal Morphology of Weanling Pig. Anim. Feed Sci. Technol. 2014, 188, 102–110. [Google Scholar] [CrossRef]
- Sonia, T.A.; Ji, H.; Hong-Seok, M.; Chul-Ju, Y. Evaluation of Lactobacillus and Bacillus-Based Probiotics as Alternatives to Antibiotics in Enteric Microbial Challenged Weaned Piglets. Afr. J. Microbiol. Res. 2014, 8, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Guo, Y.; Guo, S.; Tan, J. Effects of Clostridium Butyricum and Enterococcus Faecium on Growth Performance, Lipid Metabolism, and Cecal Microbiota of Broiler Chickens. Appl. Microbiol. Biotechnol. 2013, 97, 6477–6488. [Google Scholar] [CrossRef] [PubMed]
- Mohana Devi, S.; Kim, I. Effect of Medium Chain Fatty Acids (MCFA) and Probiotic (Enterococcus faecium) Supplementation on the Growth Performance, Digestibility and Blood Profiles in Weanling Pigs. Vet. Med. 2014, 59, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Duarte, M.E.; Kim, S.W. Postbiotic Effects of Lactobacillus Fermentate on Intestinal Health, Mucosa-Associated Microbiota, and Growth Efficiency of Nursery Pigs Challenged with F18+ Escherichia coli. J. Anim. Sci. 2022, 100, skac210. [Google Scholar] [CrossRef] [PubMed]
- Veizaj-Delia, E.; Piu, T.; Lekaj, P.; Tafaj, M. Using Combined Probiotic to Improve Growth Performance of Weaned Piglets on Extensive Farm Conditions. Livest. Sci. 2010, 134, 249–251. [Google Scholar] [CrossRef]
- Huang, C.; Qiao, S.; Li, D.; Piao, X.; Ren, J. Effects of Lactobacilli on the Performance, Diarrhea Incidence, VFA Concentration and Gastrointestinal Microbial Flora of Weaning Pigs. Asian-Australas. J. Anim. Sci. 2004, 17, 401–409. [Google Scholar] [CrossRef]
- de Graaff, P.; Govers, C.; Wichers, H.J.; Debets, R. Consumption of β-Glucans to Spice up T Cell Treatment of Tumors: A Review. Expert Opin. Biol. Ther. 2018, 18, 1023–1040. [Google Scholar] [CrossRef]
- Gallois, M.; Rothkötter, H.J.; Bailey, M.; Stokes, C.R.; Oswald, I.P. Natural Alternatives to In-Feed Antibiotics in Pig Production: Can Immunomodulators Play a Role? Animal 2009, 3, 1644–1661. [Google Scholar] [CrossRef] [Green Version]
- Akira, S.; Uematsu, S.; Takeuchi, O. Pathogen Recognition and Innate Immunity. Cell 2006, 124, 783–801. [Google Scholar] [CrossRef] [Green Version]
- Li, T.-H.; Liu, L.; Hou, Y.-Y.; Shen, S.-N.; Wang, T.-T. C-Type Lectin Receptor-Mediated Immune Recognition and Response of the Microbiota in the Gut. Gastroenterol. Rep. 2019, 7, 312–321. [Google Scholar] [CrossRef] [Green Version]
- Anwar, M.I.; Muhammad, F.; Awais, M.M.; Akhtar, M. A Review of β-Glucans as a Growth Promoter and Antibiotic Alternative against Enteric Pathogens in Poultry. World’s Poult. Sci. J. 2017, 73, 651–661. [Google Scholar] [CrossRef]
- Li, J.; Li, D.F.; Xing, J.J.; Cheng, Z.B.; Lai, C.H. Effects of β-Glucan Extracted from Saccharomyces Cerevisiae on Growth Performance, and Immunological and Somatotropic Responses of Pigs Challenged with Escherichia Coli Lipopolysaccharide. J. Anim. Sci. 2006, 84, 2374–2381. [Google Scholar] [CrossRef]
- Price, K.L.; Totty, H.R.; Lee, H.B.; Utt, M.D.; Fitzner, G.E.; Yoon, I.; Ponder, M.A.; Escobar, J. Use of Saccharomyces Cerevisiae Fermentation Product on Growth Performance and Microbiota of Weaned Pigs during Salmonella Infection. J. Anim. Sci. 2010, 88, 3896–3908. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Brandherm, M.; Freeland, M.; Newton, B.; Cook, D.; Yoon, I. Effects of Yeast Culture Supplementation to Gestation and Lactation Diets on Growth of Nursing Piglets. Asian-Australas. J. Anim. Sci. 2008, 21, 1011–1014. [Google Scholar] [CrossRef]
- Kim, S.W.; Brandherm, M.; Newton, B.; Cook, D.R.; Yoon, I.; Fitzner, G. Effect of Supplementing Saccharomyces Cerevisiae Fermentation Product in Sow Diets on Reproductive Performance in a Commercial Environment. Can. J. Anim. Sci. 2010, 90, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.B.; Carroll, J.A.; Yoon, I.; Mateo, R.D.; Kim, S.W. Effects of Supplementing Saccharomyces Cerevisiae Fermentation Product in Sow Diets on Performance of Sows and Nursing Piglets. J. Anim. Sci. 2011, 89, 2462–2471. [Google Scholar] [CrossRef]
- Lu, H.; Wilcock, P.; Adeola, O.; Ajuwon, K.M. Effect of Live Yeast Supplementation to Gestating Sows and Nursery Piglets on Postweaning Growth Performance and Nutrient Digestibility. J. Anim. Sci. 2019, 97, 2534–2540. [Google Scholar] [CrossRef]
- Davis, M.E.; Maxwell, C.V.; Brown, D.C.; de Rodas, B.Z.; Johnson, Z.B.; Kegley, E.B.; Hellwig, D.H.; Dvorak, R.A. Effect of Dietary Mannan Oligosaccharides and(or) Pharmacological Additions of Copper Sulfate on Growth Performance and Immunocompetence of Weanling and Growing/Finishing Pigs. J. Anim. Sci. 2002, 80, 2887–2894. [Google Scholar] [CrossRef] [Green Version]
- Onwurah, F. Effect of Baker’s Yeast (Saccharomyces cerevisiae) Inclusion in Feed and in Drinking Water on Performance of Broiler Birds. Br. J. Appl. Sci. Technol. 2014, 4, 144–151. [Google Scholar] [CrossRef]
- Zhang, A.W.; Lee, B.D.; Lee, S.K.; Lee, K.W.; An, G.H.; Song, K.B.; Lee, C.H. Effects of Yeast (Saccharomyces cerevisiae) Cell Components on Growth Performance, Meat Quality, and Ileal Mucosa Development of Broiler Chicks. Poult. Sci. 2005, 84, 1015–1021. [Google Scholar] [CrossRef]
- Sauerwein, H.; Schmitz, S.; Hiss, S. Effects of a Dietary Application of a Yeast Cell Wall Extract on Innate and Acquired Immunity, on Oxidative Status and Growth Performance in Weanling Piglets and on the Ileal Epithelium in Fattened Pigs. J. Anim. Physiol. Anim. Nutr. 2007, 91, 369–380. [Google Scholar] [CrossRef]
- Mathew, A.G.; Chattin, S.E.; Robbins, C.M.; Golden, D.A. Effects of a Direct-Fed Yeast Culture on Enteric Microbial Populations, Fermentation Acids, and Performance of Weanling Pigs. J. Anim. Sci. 1998, 76, 2138. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Lee, S.I.; Kim, I.H. Effect of Dietary Spirulina (Arthrospira) Platensis on the Growth Performance, Antioxidant Enzyme Activity, Nutrient Digestibility, Cecal Microflora, Excreta Noxious Gas Emission, and Breast Meat Quality of Broiler Chickens. Poult. Sci. 2018, 97, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.A.; Morlacchini, M.; Keegan, J.D.; Fusconi, G. Dietary Supplementation of Finishing Pigs with the Docosahexaenoic Acid-Rich Microalgae, Aurantiochytrium Limacinum: Effects on Performance, Carcass Characteristics and Tissue Fatty Acid Profile. Asian-Australas. J. Anim. Sci. 2018, 31, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Waldenstedt, L.; Inborr, J.; Hansson, I.; Elwinger, K. Effects of Astaxanthin-Rich Algal Meal (Haematococcus pluvalis) on Growth Performance, Caecal Campylobacter and Clostridial Counts and Tissue Astaxanthin Concentration of Broiler Chickens. Anim. Feed Sci. Technol. 2003, 108, 119–132. [Google Scholar] [CrossRef]
- Kibria, S.; Kim, I.H. Impacts of Dietary Microalgae (Schizochytrium JB5) on Growth Performance, Blood Profiles, Apparent Total Tract Digestibility, and Ileal Nutrient Digestibility in Weaning Pigs. J. Sci. Food Agric. 2019, 99, 6084–6088. [Google Scholar] [CrossRef]
- Raja, R.; Hemaiswarya, S.; Kumar, N.A.; Sridhar, S.; Rengasamy, R. A Perspective on the Biotechnological Potential of Microalgae. Crit. Rev. Microbiol. 2008, 34, 77–88. [Google Scholar] [CrossRef]
- Mason, R.P.; Libby, P.; Bhatt, D.L. Emerging Mechanisms of Cardiovascular Protection for the Omega-3 Fatty Acid Eicosapentaenoic Acid. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1135–1147. [Google Scholar] [CrossRef]
- Sardi, L.; Martelli, G.; Lambertini, L.; Parisini, P.; Mordenti, A. Effects of a Dietary Supplement of DHA-Rich Marine Algae on Italian Heavy Pig Production Parameters. Livest. Sci. 2006, 103, 95–103. [Google Scholar] [CrossRef]
- Baňoch, T.; Svoboda, M.; Kuta, J.; Saláková, A.; Fajt, Z. The Effect of Iodine from Iodine-Enriched Alga Chlorella Spp. on the Pork Iodine Content and Meat Quality in Finisher Pigs. Acta Vet. Brno 2012, 81, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Pourkarimi, S.; Hallajisani, A.; Alizadehdakhel, A.; Nouralishahi, A.; Golzary, A. Factors Affecting Production of Beta-Carotene from Dunaliella Salina Microalgae. Biocatal. Agric. Biotechnol. 2020, 29, 101771. [Google Scholar] [CrossRef]
- Brito, A.d.F.; Silva, A.S.; de Oliveira, C.V.C.; de Souza, A.A.; Ferreira, P.B.; de Souza, I.L.L.; da Cunha Araujo, L.C.; da Silva Félix, G.; de Souza Sampaio, R.; Tavares, R.L.; et al. Spirulina Platensis Prevents Oxidative Stress and Inflammation Promoted by Strength Training in Rats: Dose-Response Relation Study. Sci. Rep. 2020, 10, 6382. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Kiros, T.G.; Derakhshani, H.; Pinloche, E.; D’Inca, R.; Marshall, J.; Auclair, E.; Khafipour, E.; Van Kessel, A. Effect of Live Yeast Saccharomyces Cerevisiae (Actisaf Sc 47) Supplementation on the Performance and Hindgut Microbiota Composition of Weanling Pigs. Sci. Rep. 2018, 8, 5315. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The Role of Short-Chain Fatty Acids in the Interplay between Diet, Gut Microbiota, and Host Energy Metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Jiao, A.; Diao, H.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Luo, Y.; Luo, J.; Wang, Q.; Wang, H.; et al. Infusion of Short Chain Fatty Acids in the Ileum Improves the Carcass Traits, Meat Quality and Lipid Metabolism of Growing Pigs. Anim. Nutr. 2021, 7, 94–100. [Google Scholar] [CrossRef]
- Lum, K.K.; Kim, J.; Lei, X.G. Dual Potential of Microalgae as a Sustainable Biofuel Feedstock and Animal Feed. J. Anim. Sci. Biotechnol. 2013, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Taliercio, E.; Kim, S.W. Epitopes from Two Soybean Glycinin Subunits Are Antigenic in Pigs. J. Sci. Food Agric. 2013, 93, 2927–2932. [Google Scholar] [CrossRef]
- Bosi, P.; Han, I.K.; Jung, H.J.; Heo, K.N.; Perini, S.; Castellazzi, A.M.; Casini, L.; Creston, D.; Gremokolini, C. Effect of Different Spray Dried Plasmas on Growth, Ileal Digestibility, Nutrient Deposition, Immunity and Health of Early-Weaned Pigs Challenged with E. Coli K88. Asian-Australas. J. Anim. Sci. 2001, 14, 1138–1143. [Google Scholar] [CrossRef]
- Olsen, R.L.; Hasan, M.R. A Limited Supply of Fishmeal: Impact on Future Increases in Global Aquaculture Production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Nasseri, A.T.; Rasoul-Ami, S.; Morowvat, M.H.; Ghasemi, Y. Single Cell Protein: Production and Process. Am. J. Food Technol. 2011, 6, 103–116. [Google Scholar] [CrossRef]
- Becker, P.M. Single Cell Proteins in Diets for Weanling Pigs; Animal Sciences Group: Wageningen, The Netherlands, 2014. [Google Scholar]
- Gatrell, S.; Lum, K.; Kim, J.; Lei, X.G. Nonruminant Nutrition Symposium: Potential of Defatted Microalgae from the Biofuel Industry as an Ingredient to Replace Corn and Soybean Meal in Swine and Poultry Diets. J. Anim. Sci. 2014, 92, 1306–1314. [Google Scholar] [CrossRef] [PubMed]
- Øverland, M.; Skrede, A.; Matre, T. Bacterial Protein Grown on Natural Gas as Feed for Pigs. Acta Agric. Scand. Sect. A Anim. Sci. 2001, 51, 97–106. [Google Scholar] [CrossRef]
- An, B.-K.; Choi, Y.-I.; Kang, C.-W.; Lee, K.-W. Effects of Dietary Corynebacterium Ammoniagenes-Derived Single Cell Protein on Growth Performance, Blood and Tibia Bone Characteristics, and Meat Quality of Broiler Chickens. J. Anim. Feed Sci. 2018, 27, 140–147. [Google Scholar] [CrossRef]
- Cruz, A.; Sterten, H.; Steinhoff, F.S.; Mydland, L.T.; Øverland, M. Cyberlindnera Jadinii Yeast as a Protein Source for Broiler Chickens: Effects on Growth Performance and Digestive Function from Hatching to 30 Days of Age. Poult. Sci. 2020, 99, 3168–3178. [Google Scholar] [CrossRef]
- Ekmay, R.; Gatrell, S.; Lum, K.; Kim, J.; Lei, X.G. Nutritional and Metabolic Impacts of a Defatted Green Marine Microalgal (Desmodesmus sp.) Biomass in Diets for Weanling Pigs and Broiler Chickens. J. Agric. Food Chem. 2014, 62, 9783–9791. [Google Scholar] [CrossRef]
- Øverland, M.; Kjos, N.P.; Olsen, E.; Skrede, A. Changes in Fatty Acid Composition and Improved Sensory Quality of Backfat and Meat of Pigs Fed Bacterial Protein Meal. Meat Sci. 2005, 71, 719–729. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Mollah, M. Autolyzed Saccharomyces Cerevisiae as a Single Cell Protein for Broiler Diet. Bangladesh J. Anim. Sci. 2019, 48, 1–8. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012; ISBN 978-0-309-22423-9. [Google Scholar]
- Zhang, H.Y.; Piao, X.S.; Li, P.; Yi, J.Q.; Zhang, Q.; Li, Q.Y.; Liu, J.D.; Wang, G.Q. Effects of Single Cell Protein Replacing Fish Meal in Diet on Growth Performance, Nutrient Digestibility and Intestinal Morphology in Weaned Pigs. Asian-Australas. J. Anim. Sci. 2013, 26, 1320–1328. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Y.-C.; Duarte, M.E.; Kim, S.W. Nutritional and Functional Values of Lysed Corynebacterium Glutamicum Cell Mass for Intestinal Health and Growth of Nursery Pigs. J. Anim. Sci. 2021, 99, skab331. [Google Scholar] [CrossRef]
- D’Mello, J.P.F.; Acamovic, T. Evaluation of Methanol-grown Bacteria as a Source of Protein and Energy for Young Chicks. Br. Poult. Sci. 1976, 17, 393–401. [Google Scholar] [CrossRef]
- Braude, R.; Rhodes, D.N. Pruteen, a New Source of Protein for Growing Pigs. II. Feeding Trial: Growth Rate, Feed Utilization and Carcass and Meat Quality. Livest. Prod. Sci. 1977, 4, 91–100. [Google Scholar] [CrossRef]
- Schøyen, H.F.; Svihus, B.; Storebakken, T.; Skrede, A. Bacterial Protein Meal Produced on Natural Gas Replacing Soybean Meal or Fish Meal in Broiler Chicken Diets. Arch. Anim. Nutr. 2007, 61, 276–291. [Google Scholar] [CrossRef]
- Øverland, M.; Tauson, A.-H.; Shearer, K.; Skrede, A. Evaluation of Methane-Utilising Bacteria Products as Feed Ingredients for Monogastric Animals. Arch. Anim. Nutr. 2010, 64, 171–189. [Google Scholar] [CrossRef]
- Skrede, A.; Berge, G.; Storebakken, T.; Herstad, O.; Aarstad, K.; Sundstøl, F. Digestibility of Bacterial Protein Grown on Natural Gas in Mink, Pigs, Chicken and Atlantic Salmon. Anim. Feed Sci. Technol. 1998, 76, 103–116. [Google Scholar] [CrossRef]
- Tlusty, M.; Rhyne, A.; Szczebak, J.T.; Bourque, B.; Bowen, J.L.; Burr, G.; Marx, C.J.; Feinberg, L. A Transdisciplinary Approach to the Initial Validation of a Single Cell Protein as an Alternative Protein Source for Use in Aquafeeds. PeerJ 2017, 5, e3170. [Google Scholar] [CrossRef] [Green Version]
- Hellwing, A.L.F.; Tauson, A.-H.; Skrede, A.; Kjos, N.P.; Ahlstrøm, Ø. Bacterial Protein Meal in Diets for Pigs and Minks: Comparative Studies on Protein Turnover Rate and Urinary Excretion of Purine Base Derivatives. Arch. Anim. Nutr. 2007, 61, 425–443. [Google Scholar] [CrossRef]
- Chen, Y.; Chi, S.; Zhang, S.; Dong, X.; Yang, Q.; Liu, H.; Zhang, W.; Deng, J.; Tan, B.; Xie, S. Replacement of Fish Meal with Methanotroph (Methylococcus capsulatus, Bath) Bacteria Meal in the Diets of Pacific White Shrimp (Litopenaeus vannamei). Aquaculture 2021, 541, 736801. [Google Scholar] [CrossRef]
- Zhang, Q.; Liang, H.; Longshaw, M.; Wang, J.; Ge, X.; Zhu, J.; Li, S.; Ren, M. Effects of Replacing Fishmeal with Methanotroph (Methylococcus capsulatus, Bath) Bacteria Meal (FeedKind®) on Growth and Intestinal Health Status of Juvenile Largemouth Bass (Micropterus salmoides). Fish Shellfish Immunol. 2022, 122, 298–305. [Google Scholar] [CrossRef]
- Plavnik, I.; Bornstein, S.; Hurwitz, S. Evaluation of Methanol-grown Bacteria and Hydrocarbon-grown Yeast as Sources of Protein for Poultry: Studies with Young Chicks. Br. Poult. Sci. 1981, 22, 123–140. [Google Scholar] [CrossRef]
- Sahm, H.; Eggeling, L. New Ubiquitous Translocators: Amino Acid Export by Corynebacterium Glutamicum and Escherichia Coli. Arch. Microbiol. 2003, 180, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Leuchtenberger, W.; Huthmacher, K.; Drauz, K. Biotechnological Production of Amino Acids and Derivatives: Current Status and Prospects. Appl. Microbiol. Biotechnol. 2005, 69, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, A.; Zeng, X.; Hou, C.; Liu, H.; Qiao, S. Lactobacillus Reuteri I5007 Modulates Tight Junction Protein Expression in IPEC-J2 Cells with LPS Stimulation and in Newborn Piglets under Normal Conditions. BMC Microbiol. 2015, 15, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.P.; Kim, J.D.; Kim, J.E.; Kim, I.H. Amino Acid Digestibility of Single Cell Protein from Corynebacterium Ammoniagenes in Growing Pigs. Anim. Feed Sci. Technol. 2013, 180, 111–114. [Google Scholar] [CrossRef]
- Whittemore, C.T.; Moffat, I.W.; Taylor, A.G. Evaluation by Digestibility, Growth and Slaughter of Microbial Cells as a Source of Protein for Young Pigs. J. Sci. Food Agric. 1976, 27, 1163–1170. [Google Scholar] [CrossRef]
- Yáñez, E.; Ballester, D.; Fernández, N.; Gattás, V.; Monckeberg, F. Chemical Composition of Candida Utilis and the Biological Quality of the Yeast Protein. J. Sci. Food Agric. 1972, 23, 581–586. [Google Scholar] [CrossRef]
- Lagos, L.V.; Stein, H.H. Torula Yeast Has Greater Digestibility of Amino Acids and Phosphorus, but Not Energy, Compared with a Commercial Source of Fish Meal Fed to Weanling Pigs. J. Anim. Sci. 2020, 98, skz375. [Google Scholar] [CrossRef]
- Reed, G.; Nagodawithana, T.W. Yeast Technology; Van Nostrand Reinhold: New York, NY, USA, 1991. [Google Scholar]
- Chand, N.; Khan, R.U. Replacement of Soybean Meal with Yeast Single Cell Protein in Broiler Ration: The Effect on Performance Traits. Pak. J. Zool. 2014, 46, 1753–1758. [Google Scholar]
- Lagos, L.; Bekkelund, A.K.; Skugor, A.; Ånestad, R.; Åkesson, C.P.; Press, C.M.; Øverland, M. Cyberlindnera Jadinii Yeast as a Protein Source for Weaned Piglets—Impact on Immune Response and Gut Microbiota. Front. Immunol. 2020, 11, 1924. [Google Scholar] [CrossRef]
- Cruz, A.; Håkenåsen, I.M.; Skugor, A.; Mydland, L.T.; Åkesson, C.P.; Hellestveit, S.S.; Sørby, R.; Press, C.M.; Øverland, M. Candida Utilis Yeast as a Protein Source for Weaned Piglets: Effects on Growth Performance and Digestive Function. Livest. Sci. 2019, 226, 31–39. [Google Scholar] [CrossRef]
- van Heugten, E.; Funderburke, D.W.; Dorton, K.L. Growth Performance, Nutrient Digestibility, and Fecal Microflora in Weanling Pigs Fed Live Yeast. J. Anim. Sci. 2003, 81, 1004–1012. [Google Scholar] [CrossRef]
- Kim, B.G.; Liu, Y.; Stein, H.H. Energy Concentration and Phosphorus Digestibility in Yeast Products Produced from the Ethanol Industry, and in Brewers’ Yeast, Fish Meal, and Soybean Meal Fed to Growing Pigs1. J. Anim. Sci. 2014, 92, 5476–5484. [Google Scholar] [CrossRef]
- Valdivie, M.; Compte, X.; Fundora, O. The Utilization of Torula Yeast in Diets for White Leghorn Birds during Growth and Laying Periods. Anim. Feed Sci. Technol. 1982, 7, 185–190. [Google Scholar] [CrossRef]
- Espinosa, C.D.; Lagos, L.V.; Stein, H.H. Effect of Torula Yeast on Growth Performance, Diarrhea Incidence, and Blood Characteristics in Weanling Pigs. J. Anim. Sci. 2020, 98, skaa307. [Google Scholar] [CrossRef]
- Czech, A.; Smolczyk, A.; Ognik, K.; Kiesz, M. Nutritional Value of Yarrowia Lipolytica Yeast and Its Effect on Growth Performance Indicators in Piglets. Ann. Anim. Sci. 2016, 16, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Lipstein, B.; Hurwitz, S. The Nutritional Value of Algae for Poultry. Dried Chlorella in Broiler Diets. Br. Poult. Sci. 1980, 21, 9–21. [Google Scholar] [CrossRef]
- Coelho, D.; Pestana, J.; Almeida, J.M.; Alfaia, C.M.; Fontes, C.M.G.A.; Moreira, O.; Prates, J.A.M. A High Dietary Incorporation Level of Chlorella Vulgaris Improves the Nutritional Value of Pork Fat without Impairing the Performance of Finishing Pigs. Animals 2020, 10, 2384. [Google Scholar] [CrossRef]
- Alfaia, C.M.; Pestana, J.M.; Rodrigues, M.; Coelho, D.; Aires, M.J.; Ribeiro, D.M.; Major, V.T.; Martins, C.F.; Santos, H.; Lopes, P.A.; et al. Influence of Dietary Chlorella Vulgaris and Carbohydrate-Active Enzymes on Growth Performance, Meat Quality and Lipid Composition of Broiler Chickens. Poult. Sci. 2021, 100, 926–937. [Google Scholar] [CrossRef]
- Gatrell, S.K.; Derksen, T.J.; O’Neil, E.V.; Lei, X.G. A New Type of Defatted Green Microalgae Exerts Dose-Dependent Nutritional, Metabolic, and Environmental Impacts in Broiler Chicks. J. Appl. Poult. Res. 2017, 26, 358–366. [Google Scholar] [CrossRef]
- Czech, A.; Smolczyk, A.; Ognik, K.; Wlazło, Ł.; Nowakowicz-Dębek, B.; Kiesz, M. Effect of Dietary Supplementation with Yarrowia Lipolytica or Saccharomyces Cerevisiae Yeast and Probiotic Additives on Haematological Parameters and the Gut Microbiota in Piglets. Res. Vet. Sci. 2018, 119, 221–227. [Google Scholar] [CrossRef]
- Foltz, K.L.; Smith, D.L.; Moritz, J.S. Porcine Feed Intake of Corn–Soybean Based Diets Supplemented with Oil-Extracted Microalgae and Subsequent Performance. Prof. Anim. Sci. 2016, 32, 849–853. [Google Scholar] [CrossRef]
- Manor, M.L.; Kim, J.; Derksen, T.J.; Schwartz, R.L.; Roneker, C.A.; Bhatnagar, R.S.; Lei, X.G. Defatted Microalgae Serve as a Dual Dietary Source of Highly Bioavailable Iron and Protein in an Anemic Pig Model. Algal Res. 2017, 26, 409–414. [Google Scholar] [CrossRef]
- Madeira, M.S.; Cardoso, C.; Lopes, P.A.; Coelho, D.; Afonso, C.; Bandarra, N.M.; Prates, J.A.M. Microalgae as Feed Ingredients for Livestock Production and Meat Quality: A Review. Livest. Sci. 2017, 205, 111–121. [Google Scholar] [CrossRef]
- Pestana, J.M.; Puerta, B.; Santos, H.; Madeira, M.S.; Alfaia, C.M.; Lopes, P.A.; Pinto, R.M.A.; Lemos, J.P.C.; Fontes, C.M.G.A.; Lordelo, M.M.; et al. Impact of Dietary Incorporation of Spirulina (Arthrospira platensis) and Exogenous Enzymes on Broiler Performance, Carcass Traits, and Meat Quality. Poult. Sci. 2020, 99, 2519–2532. [Google Scholar] [CrossRef] [PubMed]
- Toyomizu, M.; Sato, K.; Taroda, H.; Kato, T.; Akiba, Y. Effects of Dietary Spirulina on Meat Colour in Muscle of Broiler Chickens. Br. Poult. Sci. 2001, 42, 197–202. [Google Scholar] [CrossRef]
- Grinstead, G.S.; Tokach, M.D.; Dritz, S.S.; Goodband, R.D.; Nelssen, J.L. Effects of Spirulina Platensis on Growth Performance of Weanling Pigs. Anim. Feed Sci. Technol. 2000, 83, 237–247. [Google Scholar] [CrossRef]
- El-Bahr, S.; Shousha, S.; Shehab, A.; Khattab, W.; Ahmed-Farid, O.; Sabike, I.; El-Garhy, O.; Albokhadaim, I.; Albosadah, K. Effect of Dietary Microalgae on Growth Performance, Profiles of Amino and Fatty Acids, Antioxidant Status, and Meat Quality of Broiler Chickens. Animals 2020, 10, 761. [Google Scholar] [CrossRef]
- Gatrell, S.K.; Magnuson, A.D.; Barcus, M.; Lei, X.G. Graded Levels of a Defatted Green Microalgae Inclusion in Diets for Broiler Chicks Led to Moderate Up-Regulation of Protein Synthesis Pathway in the Muscle and Liver. Algal Res. 2018, 29, 290–296. [Google Scholar] [CrossRef]
- Sun, T.; Wang, K.; Wyman, B.; Sudibyo, H.; Liu, G.; Beal, C.; Manning, S.; Johnson, Z.I.; Aydemir, T.B.; Tester, J.W.; et al. Supplemental Dietary Full-Fatted and Defatted Desmodesmus Sp. Exerted Similar Effects on Growth Performance, Gut Health, and Excreta Hydrothermal Liquefaction of Broiler Chicks. Algal Res. 2021, 54, 102205. [Google Scholar] [CrossRef]
- Martins, C.F.; Pestana Assunção, J.; Ribeiro Santos, D.M.; Madeira, M.S.M.D.S.; Alfaia, C.M.R.P.M.; Lopes, P.A.A.B.; Coelho, D.F.M.; Cardoso Lemos, J.P.; Almeida, A.M.; Mestre Prates, J.A.; et al. Effect of Dietary Inclusion of Spirulina on Production Performance, Nutrient Digestibility and Meat Quality Traits in Post-weaning Piglets. J. Anim. Physiol. Anim. Nutr. 2021, 105, 247–259. [Google Scholar] [CrossRef]
- Schøyen, H.F.; Frøyland, J.R.K.; Sahlström, S.; Knutsen, S.H.; Skrede, A. Effects of Autolysis and Hydrolysis of Bacterial Protein Meal Grown on Natural Gas on Chemical Characterization and Amino Acid Digestibility. Aquaculture 2005, 248, 27–33. [Google Scholar] [CrossRef]
- Øverland, M.; Skrede, A. Yeast Derived from Lignocellulosic Biomass as a Sustainable Feed Resource for Use in Aquaculture. J. Sci. Food Agric. 2017, 97, 733–742. [Google Scholar] [CrossRef]
- Øverland, M.; Schøyen, H.F.; Skrede, A. Growth Performance and Carcase Quality in Broiler Chickens Fed on Bacterial Protein Grown on Natural Gas. Br. Poult. Sci. 2010, 51, 686–695. [Google Scholar] [CrossRef]
- Øverland, M.; Mydland, L.T.; Skrede, A. Marine Macroalgae as Sources of Protein and Bioactive Compounds in Feed for Monogastric Animals. J. Sci. Food Agric. 2019, 99, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Ritala, A.; Häkkinen, S.T.; Toivari, M.; Wiebe, M.G. Single Cell Protein—State-of-the-Art, Industrial Landscape and Patents 2001–2016. Front. Microbiol. 2017, 8, 2009. [Google Scholar] [CrossRef] [Green Version]
- Mielke, T. World Markets for Vegetable Oils and Animal Fats. In Biokerosene; Springer: Berlin/Heidelberg, Germany, 2018; pp. 147–188. ISBN 9783662530658. [Google Scholar]
- Cera, K.R.; Mahan, D.C.; Reinhart, G.A. Weekly Digestibilities of Diets Supplemented with Corn Oil, Lard or Tallow by Weanling Swine. J. Anim. Sci. 1988, 66, 1430. [Google Scholar] [CrossRef]
- Cera, K.R.; Mahan, D.C.; Reinhart, G.A. Apparent Fat Digestibilities and Performance Responses of Postweaning Swine Fed Diets Supplemented with Coconut Oil, Corn Oil or Tallow. J. Anim. Sci. 1989, 67, 2040. [Google Scholar] [CrossRef]
- USDA. Oil Crops Yearbook; Economic Research Service, U.S. Department of Agriculture: Washington, DC, USA, 2020.
- European Parliament, C. of the E.U. Regulation 1069/2009. Regul. No 1069/2009 European Parliament and of the Council of 21 October 2009 Lay down Health rules as regards Anim. by-products Derived Products not Intended for Human Consumption repealing Regulation no. 1774/2002. Off. J. Eur. Union 2009, 52, 109.
- Park, S.W.; Seo, S.H.; Chang, M.B.; Shin, I.S.; Paik, I.K. Evaluation of Soybean Oil as a Lipid Source for Pig Diets. Asian-Australasian J. Anim. Sci. 2009, 22, 1311–1319. [Google Scholar] [CrossRef]
- Shurson, G.C.; Kerr, B.J.; Hanson, A.R. Evaluating the Quality of Feed Fats and Oils and Their Effects on Pig Growth Performance. J. Anim. Sci. Biotechnol. 2015, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Darvishi, F.; Salmani, N.; Hosseini, B. Biovalorization of Vegetable Oil Refinery Wastewater into Value-added Compounds by Yarrowia Lipolytica. J. Chem. Technol. Biotechnol. 2019, 94, 2961–2968. [Google Scholar] [CrossRef]
- Abril, R.; Garrett, J.; Zeller, S.G.; Sander, W.J.; Mast, R.W. Safety Assessment of DHA-Rich Microalgae from Schizochytrium Sp. Part V: Target Animal Safety/Toxicity Study in Growing Swine. Regul. Toxicol. Pharmacol. 2003, 37, 73–82. [Google Scholar] [CrossRef]
- Alvarez, H.M.; Kalscheuer, R.; Steinbüchel, A. Accumulation and Mobilization of Storage Lipids by Rhodococcus Opacus PD630 and Rhodococcus Ruber NCIMB 40126. Appl. Microbiol. Biotechnol. 2000, 54, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Galán, B.; Santos-merino, M.; Nogales, J. Health Consequences of Microbial Interactions with Hydrocarbons, Oils, and Lipids. Goldfine, H., Ed.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-15146-1. [Google Scholar]
- Thies, F.; Peterson, L.D.; Powell, J.R.; Nebe-von-Caron, G.; Hurst, T.L.; Matthews, K.R.; Newsholme, E.A.; Calder, P.C. Manipulation of the Type of Fat Consumed by Growing Pigs Affects Plasma and Mononuclear Cell Fatty Acid Compositions and Lymphocyte and Phagocyte Functions. J. Anim. Sci. 1999, 77, 137. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.F.; Kim, J.H.; Han, I.K.; Jung, H.J.; Kwon, C.H. Effects of Various Fat Sources and Lecithin on the Growth Performances and Nutrient Utilization in Pigs Weaned at 21 Days of Age. Asian-Australas. J. Anim. Sci. 1998, 11, 176–184. [Google Scholar] [CrossRef]
- Lindblom, S.C.; Dozier, W.A.; Shurson, G.C.; Kerr, B.J. Digestibility of Energy and Lipids and Oxidative Stress in Nursery Pigs Fed Commercially Available Lipids. J. Anim. Sci. 2017, 95, 239. [Google Scholar] [CrossRef]
- Lauridsen, C.; Bruun Christensen, T.; Halekoh, U.; Krogh Jensen, S. Alternative Fat Sources to Animal Fat for Pigs. Lipid Technol. 2007, 19, 156–159. [Google Scholar] [CrossRef] [Green Version]
- van Heugten, E.; Coffey, M.T.; Spears, J.W. Effects of Immune Challenge, Dietary Energy Density, and Source of Energy on Performance and Immunity in Weanling Pigs. J. Anim. Sci. 1996, 74, 2431. [Google Scholar] [CrossRef]
- Gatlin, L.A.; See, M.T.; Larick, D.K.; Lin, X.; Odle, J. Conjugated Linoleic Acid in Combination with Supplemental Dietary Fat Alters Pork Fat Quality. J. Nutr. 2002, 132, 3105–3112. [Google Scholar] [CrossRef] [Green Version]
- Sousa, R.V.; Fialho, E.T.; Lima, J.A.F.; Alvarez-Leite, J.I.; Cortez, W.C.; Ferreira, M.S.S. Effect of Different Oils in Diets for Finishing Pigs: Performance, Carcass Traits and Fatty Acid Profile of the Meat. Anim. Prod. Sci. 2010, 50, 863. [Google Scholar] [CrossRef]
- Pickova, J.; Mørkøre, T. Alternate Oils in Fish Feeds. Eur. J. Lipid Sci. Technol. 2007, 109, 256–263. [Google Scholar] [CrossRef]
- Blomqvist, J.; Pickova, J.; Tilami, S.K.; Sampels, S.; Mikkelsen, N.; Brandenburg, J.; Sandgren, M.; Passoth, V. Oleaginous Yeast as a Component in Fish Feed. Sci. Rep. 2018, 8, 15945. [Google Scholar] [CrossRef] [Green Version]
- Hatlen, B.; Berge, G.M.; Odom, J.M.; Mundheim, H.; Ruyter, B. Growth Performance, Feed Utilisation and Fatty Acid Deposition in Atlantic Salmon, Salmo Salar L., Fed Graded Levels of High-Lipid/High-EPA Yarrowia Lipolytica Biomass. Aquaculture 2012, 364–365, 39–47. [Google Scholar] [CrossRef]
- Eryalçin, K.M.; Ganuza, E.; Atalah, E.; Cruz, M.C.H. Nannochloropsis Gaditana and Crypthecodinium Cohnii, Two Microalgae as Alternative Sources of Essential Fatty Acids in Early Weaning for Gilthead Seabream. Hidrobiologica 2015, 25, 193–202. [Google Scholar]
- Santigosa, E.; Constant, D.; Prudence, D.; Wahli, T.; Verlhac-Trichet, V. A Novel Marine Algal Oil Containing Both EPA and DHA Is an Effective Source of Omega-3 Fatty Acids for Rainbow Trout Oncorhynchus Mykiss. J. World Aquac. Soc. 2020, 51, 649–665. [Google Scholar] [CrossRef]
- Barclay, W.; Zeller, S. Nutritional Enhancement of N-3 and n-6 Fatty Acids in Rotifers and Artemia Nauplii by Feeding Spray-Dried Schizochytrium sp. J. World Aquac. Soc. 1996, 27, 314–322. [Google Scholar] [CrossRef]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijck, P.W.M.; Wyss, M. Yarrowia Lipolytica: Safety Assessment of an Oleaginous Yeast with a Great Industrial Potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Khot, M.; Kamat, S.; Zinjarde, S.; Pant, A.; Chopade, B.; RaviKumar, A. Single Cell Oil of Oleaginous Fungi from the Tropical Mangrove Wetlands as a Potential Feedstock for Biodiesel. Microb. Cell Fact. 2012, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- den Haan, R.; van Rensburg, E.; Rose, S.H.; Görgens, J.F.; van Zyl, W.H. Progress and Challenges in the Engineering of Non-Cellulolytic Microorganisms for Consolidated Bioprocessing. Curr. Opin. Biotechnol. 2015, 33, 32–38. [Google Scholar] [CrossRef]
- Cheng, Y.-C.; Duarte, M.E.; Kim, S.W. Effects of Yarrowia Lipolytica Supplementation on Growth Performance, Intestinal Health and Apparent Ileal Digestibility of Diets Fed to Nursery Pigs. Anim. Biosci. 2022, 35, 605–613. [Google Scholar] [CrossRef]
- Berge, G.M.; Hatlen, B.; Odom, J.M.; Ruyter, B. Physical Treatment of High EPA Yarrowia Lipolytica Biomass Increases the Availability of N-3 Highly Unsaturated Fatty Acids When Fed to Atlantic Salmon. Aquac. Nutr. 2013, 19, 110–121. [Google Scholar] [CrossRef]
- Sun, T.; Tolba, S.A.; Magnuson, A.D.; Lei, X.G. Excessive Aurantiochytrium Acetophilum Docosahexaenoic Acid Supplementation Decreases Growth Performance and Breast Muscle Mass of Broiler Chickens. Algal Res. 2022, 63, 102648. [Google Scholar] [CrossRef]
- Lee, A.; You, L.; Oh, S.-Y.; Li, Z.; Code, A.; Zhu, C.; Fisher-Heffernan, R.; Regnault, T.; De Lange, C.; Huber, L.-A.; et al. Health Benefits of Supplementing Nursery Pig Diets with Microalgae or Fish Oil. Animals 2019, 9, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Tonnac, A.; Labussière, E.; Vincent, A.; Mourot, J. Effect of α -Linolenic Acid and DHA Intake on Lipogenesis and Gene Expression Involved in Fatty Acid Metabolism in Growing-Finishing Pigs. Br. J. Nutr. 2016, 116, 7–18. [Google Scholar] [CrossRef] [Green Version]
- De Tonnac, A.; Guillevic, M.; Mourot, J. Fatty Acid Composition of Several Muscles and Adipose Tissues of Pigs Fed N-3 PUFA Rich Diets. Meat Sci. 2018, 140, 1–8. [Google Scholar] [CrossRef]
- Patil, V.; Reitan, K.; Mortensen, L.; Källqvist, T.; Olsen, Y.; Vogt, G.; Gislerød, H. Microalgae as a Source of Polyunsaturated Fatty Acids for Aquaculture. Curr. Top. Plant Biol. 2005, 6, 57–65. [Google Scholar]
- Harel, M.; Koven, W.; Lein, I.; Bar, Y.; Behrens, P.; Stubblefield, J.; Zohar, Y.; Place, A.R. Advanced DHA, EPA and ArA Enrichment Materials for Marine Aquaculture Using Single Cell Heterotrophs. Aquaculture 2002, 213, 347–362. [Google Scholar] [CrossRef]
- Atalah, E.; Cruz, C.M.H.; Izquierdo, M.S.; Rosenlund, G.; Caballero, M.J.; Valencia, A.; Robaina, L. Two Microalgae Crypthecodinium Cohnii and Phaeodactylum tricornutum as Alternative Source of Essential Fatty Acids in Starter Feeds for Seabream (Sparus aurata). Aquaculture 2007, 270, 178–185. [Google Scholar] [CrossRef]


| Micro-organism | Species | Feedstuff | Product | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | |||||||
| Bacteria | Bacillus sp. | probiotic | 1.0 × 1010 CFU/g | 0.01 | 11.1 | 2.94 | 8.28 | Nursery pigs | [80] |
| Bacteria | Bacillus sp. | probiotic | 1.0 × 109 CFU/kg | 0.05 | 18.83 | 20.5 | −0.91 | Nursery pigs | [81] |
| 0.05 | 2.44 | −0.80 | 1.21 | ||||||
| Bacteria | Bacillus sp. | probiotic | 3.2 × 109 CFU/kg | 0.04 | 2.65 | 0.81 | 1.94 | Growing-finishing pigs | [85] |
| Bacteria | Bacillus sp. | probiotic | 6.0 × 108 CFU/g | 0.05 | 9.80 | −0.44 | 10.4 | Growing pigs | [86] |
| 0.03 | 4.93 | −0.15 | 5.11 | ||||||
| Bacteria | Bacillus sp. | probiotic | 2.4 × 108 CFU/g | 0.45 | 9.12 | 4.87 | 4.17 | Nursery pigs | [87] |
| 0.30 | 6.92 | 4.06 | 2.78 | ||||||
| 0.15 | 2.20 | 1.01 | 1.24 | ||||||
| Bacteria | Bacillus sp. | probiotic | 3.2 × 109 CFU/kg | 0.04 | 2.41 | −10.6 | 6.63 | Nursery pigs | [88] |
| Bacteria | Clostridium butyricum | probiotic | 1.0 × 109 CFU/kg | 0.05 | 2.54 | 1.76 | 0.00 | Broilers | [89] |
| Bacteria | Enterococcus faecium | probiotic | 2.0 × 109 CFU/kg | 0.05 | 4.93 | 5.14 | 0.00 | Broilers | [89] |
| Bacteria | Enterococcus faecium | probiotic | 2.0 × 109 CFU/kg | 0.01 | 5.50 | −0.77 | 6.01 | Nursery pigs | [90] |
| Bacteria | Lactobacillus sp. | postbiotic | 6.0 × 1010 CFU/g and medium | 0.20 | 26.1 | 20.0 | 9.52 | Nursery pigs | [91] |
| Bacteria | Lactobacillus sp. | probiotic | 1.0 × 1010 CFU/g | 0.50 | 4.56 | 36.2 | 1.10 | Nursery pigs | [88] |
| Bacteria | Lactobacillus sp. | probiotic | 5.0 × 109 CFU/kg | 0.10 | 6.06 | −7.20 | 12.5 | Nursery pigs | [92] |
| 0.15 | 3.56 | −7.12 | 10.3 | ||||||
| 0.20 | −1.89 | −10.2 | 8.44 | ||||||
| Bacteria | Lactobacillus sp. | probiotic | 2.4 × 105 CFU/g | 0.10 | 16.0 | 10.5 | 5.16 | Nursery pigs | [93] |
| 0.50 | 1.91 | 0.84 | 1.03 | ||||||
| 0.75 | 3.35 | 1.75 | 1.63 | ||||||
| 1.00 | 4.16 | 1.16 | 2.90 | ||||||
| Microorganism | Species | Feedstuff | Product | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | |||||||
| Yeast | Saccharomyces cerevisiae | Postbiotic | Glucans extracted | 0.05 | 10.6 | 5.16 | 5.38 | Broilers | [106] |
| 0.10 | 18.7 | 13.3 | 4.93 | ||||||
| 0.15 | 10.5 | 10.0 | 1.35 | ||||||
| 0.20 | 10.6 | 4.55 | 10.8 | ||||||
| Yeast | Saccharomyces cerevisiae | Prebiotic | Cell wall extract | 0.30 | 4.71 | −0.26 | 6.25 | Broilers | [107] |
| Yeast | Saccharomyces cerevisiae | Prebiotic | Cell wall extract | 0.03 | −15.7 | 0.85 | 5.88 | Nursery pigs | [108] |
| Yeast | Saccharomyces cerevisiae | Probiotic | 1.0 × 109 CFU/g | 0.10 | 4.48 | 20.1 | −15.4 | Nursery pigs | [109] |
| Yeast | Saccharomyces cerevisiae | Postbiotic | yeast culture | 0.50 | 6.31 | 9.48 | 0.00 | Nursery pigs | [43] |
| 1.00 | −6.80 | −1.07 | −2.79 | ||||||
| 2.00 | −8.01 | −5.47 | 0.93 | ||||||
| Microalgae | Arthrospira platensis | Postbiotic | Spray dried algae, Setalg (Pleubian, France) | 1.00 | −0.25 | 4.21 | −3.28 | Nursery pigs | [65] |
| Microalgae | Arthrospira platensis | Postbiotic | Spirulina powder, NeoEnBiz Co. (Bucheon, Republic of Korea) | 0.25 | 1.79 | 1.12 | 0.67 | Broilers | [110] |
| 0.50 | 1.91 | 0.84 | 1.03 | ||||||
| 0.75 | 3.35 | 1.75 | 1.63 | ||||||
| 1.00 | 4.16 | 1.16 | 2.90 | ||||||
| Microalgae | Aurantiochytrium limacinum | Postbiotic | ALL-G Rich, Alltech Inc. (Lexingtong, KE, USA) | 1.00 | 1.12 | −0.38 | 1.22 | Finishing pigs | [111] |
| Microalgae | Chlorella sp. | Postbiotic | Spray dried algae, Setalg | 1.00 | 0.51 | −1.40 | 1.64 | Nursery pigs | [65] |
| Microalgae | Haematococcus pluvialis | Postbiotic | Novasta, AstaCarotene AB (Stockholm, Sweden) | 0.04 | −2.00 | −2.20 | 0.00 | Broilers | [112] |
| 0.18 | −1.77 | −3.64 | 1.90 | ||||||
| 0.90 | −0.74 | −2.43 | 1.27 | ||||||
| Microalgae | Schizochytrium sp. | Postbiotic | JB5, JINIS Co., Ltd. (Jeonju, Republic of Korea) | 0.50 | −1.33 | 2.23 | −3.36 | Nursery pigs | [113] |
| 1.00 | 1.56 | 0.48 | 1.12 | ||||||
| Conventional Protein Supplement | Bacteria | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish Meal | Soybean Meal | Blood Plasma | Corynebacterium glutamicum | Methylophilus methylotrophus | Methylococcus capsulatus | ||||||||
| CP, % | 63.0 | 48.0 | 78.0 | 76.8 | 79.9 | 68.0 | |||||||
| Essential AA, % | Arg | 3.80 | 83% | 3.50 | 117% | 4.40 | 64% | 4.09 | 61% | 3.61 | 80% | 4.56 | 123% |
| His | 1.40 | 30% | 1.30 | 43% | 2.50 | 36% | 1.55 | 23% | 1.54 | 34% | 1.54 | 42% | |
| Ile | 2.60 | 57% | 2.10 | 70% | 2.70 | 39% | 3.35 | 50% | 3.32 | 73% | 3.01 | 81% | |
| Leu | 4.50 | 98% | 3.60 | 120% | 0.40 | 6% | 5.38 | 80% | 5.45 | 120% | 5.06 | 137% | |
| Lys | 4.60 | - | 3.00 | - | 6.90 | - | 6.74 | - | 4.54 | - | 3.70 | - | |
| Met | 1.70 | 37% | 0.70 | 23% | 0.80 | 12% | 1.26 | 19% | 1.83 | 40% | 1.72 | 46% | |
| Phe | 2.50 | 54% | 2.40 | 80% | 4.30 | 62% | 2.78 | 41% | 4.22 | 93% | 2.70 | 73% | |
| Thr | 2.60 | 57% | 1.90 | 63% | 4.50 | 65% | 3.32 | 49% | 3.97 | 87% | 2.87 | 77% | |
| Trp | 0.60 | 13% | 0.70 | 23% | 1.40 | 20% | 0.56 | 8% | 0.77 | 17% | 2.21 | 60% | |
| Val | 3.10 | 67% | 2.20 | 73% | 5.10 | 74% | 4.61 | 68% | 4.91 | 108% | 3.94 | 106% | |
| Non-essential AA, % | Ala | 3.90 | 85% | 2.10 | 70% | 4.00 | 58% | 6.26 | 93% | 6.04 | 133% | 4.64 | 125% |
| Asp | 5.40 | 117% | 5.40 | 180% | 7.40 | 107% | 6.68 | 99% | 7.52 | 166% | 5.66 | 153% | |
| Cys | 0.60 | 13% | 0.70 | 23% | 2.60 | 38% | 0.35 | 5% | 0.57 | 13% | 0.45 | 12% | |
| Glu | 7.90 | 172% | 8.50 | 283% | 10.90 | 158% | 8.86 | 131% | 10.50 | 231% | 7.35 | 198% | |
| Gly | 4.70 | 102% | 2.00 | 67% | 2.80 | 41% | 3.33 | 49% | 5.69 | 125% | 3.34 | 90% | |
| Pro | 2.90 | 63% | 2.50 | 83% | 4.30 | 62% | 2.32 | 34% | NA | NA | 2.57 | 69% | |
| Ser | 2.40 | 52% | 2.40 | 80% | 4.20 | 61% | 2.34 | 35% | 2.60 | 57% | 2.37 | 64% | |
| Tyr | 1.90 | 41% | 1.60 | 53% | 3.90 | 57% | 1.81 | 27% | 3.48 | 77% | 2.43 | 66% | |
| Reference | [137] | [137] | [137] | [138,139] | [140,141] | [135,142,143] | |||||||
| Micro-organism | Species | Product | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | ||||||
| Bacteria | Corynebacterium ammoniagenes | Protide, CJ | 1.00 | 3.69 | 1.82 | 2.08 | Broilers | [132] |
| 3.00 | −1.02 | −1.18 | 0.00 | |||||
| 5.00 | −3.07 | −1.50 | −1.56 | |||||
| Bacteria | Corynebacterium glutamicum | Bacteria and medium | 2.50 | −4.32 | −1.89 | −2.08 | Nursery pigs | [138] |
| 5.00 | −8.38 | 1.89 | −10.4 | |||||
| Bacteria | Corynebacterium glutamicum | Lysed bacteria | 0.70 | −1.27 | 0.00 | −3.64 | Nursery pigs | [139] |
| 1.40 | −1.27 | 5.31 | −9.09 | |||||
| 2.10 | 11.4 | 8.78 | 0.00 | |||||
| Bacteria | Methylococcus capsulatus | BP, Dansk Bioprotein | 10.7 | 5.59 | 1.31 | 3.44 | Growing pigs | [131] |
| 12.0 | 8.48 | 7.82 | −1.26 | Nursery pigs | ||||
| 8.00 | 5.65 | 2.42 | −2.52 | |||||
| 4.00 | −0.22 | −4.55 | 3.77 | |||||
| Bacteria | Methylococcus capsulatus | BBP, Norferm AS | 6.00 | 2.68 | −3.08 | 5.88 | Broilers | [142] |
| 4.00 | 5.01 | −0.57 | 5.29 | |||||
| 2.00 | 3.98 | 0.81 | 3.53 | |||||
| Bacteria | Methylophilus methylotrophus | Bacteria and medium | 10.0 | −1.44 | 0.20 | 1.78 | Nursery pigs | [154] |
| 20.0 | 9.41 | -0.08 | 1.78 | |||||
| Bacteria | Methylophilus methylotrophus | Bacteria and medium | 9.60 | 4.01 | 2.23 | 9.60 | Broilers | [140] |
| 19.2 | −14.2 | −6.95 | −8.09 | |||||
| Bacteria | Methylophilus methylotrophus | Bacteria and medium | 3.65 | 0.15 | −4.00 | −4.00 | Broilers | [149] |
| 6.35 | −1.00 | −4.00 | −3.00 | |||||
| 9.00 | −8.00 | −8.00 | −1.00 | |||||
| 13.6 | −13.0 | −13.0 | 0.00 | |||||
| Yeast | Microalgae | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Torula Yeast | Saccharomyces cerevisiae | Yarrowia lipolytica | Desmodesmus sp. | Chlorella sp. | Nannochloropsis oceanica | ||||||||
| CP, % | 49.1 | 44.2 | 43.5 | 31.2 | 47.7 | 38.2 | |||||||
| Essential AA, % | Arg | 2.39 | 72% | 2.29 | 73% | 1.81 | 55% | 1.50 | 94% | 3.88 | 123% | 1.99 | 88% |
| His | 0.89 | 27% | 1.05 | 34% | 0.95 | 29% | 0.50 | 31% | 0.92 | 29% | 0.64 | 28% | |
| Ile | 2.16 | 66% | 1.92 | 62% | 1.99 | 61% | 1.10 | 69% | 1.87 | 60% | 1.50 | 66% | |
| Leu | 3.16 | 96% | 2.99 | 96% | 3.10 | 94% | 2.30 | 144% | 3.58 | 114% | 2.90 | 128% | |
| Lys | 3.30 | - | 3.11 | - | 3.28 | - | 1.60 | - | 3.15 | 100% | 2.27 | - | |
| Met | 0.58 | 18% | 0.73 | 23% | 0.72 | 22% | 0.50 | 31% | 0.84 | 27% | 0.57 | 25% | |
| Phe | 1.92 | 58% | 1.82 | 58% | 1.54 | 47% | 1.30 | 81% | 2.12 | 67% | 1.57 | 69% | |
| Thr | 2.10 | 64% | 2.19 | 70% | 2.01 | 61% | 1.30 | 81% | 2.63 | 84% | 1.54 | 68% | |
| Trp | 0.59 | 18% | 0.57 | 18% | 0.65 | 20% | 0.40 | 25% | 0.24 | 7% | 0.49 | 22% | |
| Val | 2.49 | 76% | 2.24 | 72% | 2.39 | 73% | 1.60 | 100% | 3.44 | 109% | 2.13 | 94% | |
| Non-essential AA, % | Ala | 3.03 | 92% | 2.68 | 86% | 3.63 | 111% | 2.30 | 144% | 1.39 | 44% | 2.22 | 98% |
| Asp | 3.98 | 121% | 4.49 | 144% | 3.58 | 109% | 2.70 | 169% | 0.03 | 1% | 2.80 | 123% | |
| Cys | 0.46 | 14% | 0.54 | 17% | 0.44 | 13% | 0.30 | 19% | 0.42 | 13% | 0.30 | 13% | |
| Glu | 6.77 | 205% | 6.57 | 211% | 6.07 | 185% | 2.90 | 181% | 2.04 | 65% | 3.34 | 147% | |
| Gly | 1.94 | 59% | 1.75 | 56% | 1.96 | 60% | 1.70 | 106% | 2.43 | 77% | 1.92 | 85% | |
| Pro | 1.55 | 47% | 2.10 | 68% | 1.72 | 52% | 2.70 | 169% | 0.94 | 30% | 4.00 | 176% | |
| Ser | 1.78 | 54% | 2.32 | 75% | 1.82 | 55% | 1.10 | 69% | 0.78 | 25% | 1.21 | 53% | |
| Tyr | 1.48 | 45% | 1.56 | 50% | 1.50 | 46% | 1.00 | 63% | 1.77 | 56% | 1.20 | 53% | |
| References | [137,156,163,164] | [137] | [165] | [134] | [166,167,168] | [169] | |||||||
| Micro-organism | Species | Product | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | ||||||
| Yeast | Saccharomyces cerevisiae | Autolyzed yeast | 1.25 | −1.05 | 5.56 | 1.89 | Broilers | [136] |
| 2.50 | −8.99 | −0.20 | −0.47 | |||||
| 5.00 | −12.6 | 2.69 | −7.55 | |||||
| Yeast | Saccharomyces cerevisiae | Whole yeast | 0.50 | 4.32 | −0.33 | 5.68 | Broilers | [107] |
| Yeast extract | 0.30 | 3.74 | 2.45 | 2.84 | ||||
| Yeast | Saccharomyces cerevisiae | Yeast and medium | 3.00 | −3.61 | 1.60 | −2.27 | Nursery pigs | [170] |
| Yeast | Torula yeast | Extracted yeast | 4.00 | −1.00 | −3.00 | −3.00 | Broilers | [149] |
| 7.00 | −3.00 | −3.00 | −1.00 | |||||
| 10.0 | −4.00 | −3.00 | 1.00 | |||||
| 15.0 | −6.00 | −5.00 | 1.00 | |||||
| Yeast | Torula yeast | Yeast and medium | 20.0 | −4.87 | −0.77 | −4.44 | Broilers | [163] |
| 4.75 | 1.38 | −3.72 | 5.31 | |||||
| Yeast | Torula yeast | SylPro, Arbiom Inc | 10.8 | 8.76 | 2.29 | 6.76 | Nursery pigs | [164] |
| 9.00 | −3.75 | −2.93 | 2.38 | |||||
| 16.0 | −3.00 | −7.32 | 7.77 | |||||
| 23.0 | −7.87 | −12.4 | 2.69 | |||||
| Yeast | Yarrowia lipolytica | Yeast and medium | 3.00 | 12.4 | −1.25 | 11.9 | Nursery pigs | [165] |
| 6.00 | −1.81 | −1.63 | −0.63 | |||||
| Yeast | Yarrowia lipolytica | Yeast and medium | 3.00 | 2.27 | −1.20 | 2.14 | Nursery pigs | [170] |
| Micro-organism | Species | Product | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | ||||||
| Microalgae | Arthrospira platensis | Spirulina, Sopropeche | 15.0 | −11.5 | −2.42 | −10.1 | Broilers | [174] |
| Microalgae | Arthrospira platensis | Spirulina powder, Sopropeche | 10.0 | −12.4 | −3.71 | −9.46 | Nursery pigs | [180] |
| Microalgae | Chlorella sp. | Pure, whole | 7.50 | 0.21 | 2.32 | −2.50 | Broilers | [166] |
| 15.0 | −2.07 | −0.51 | −1.87 | |||||
| Microalgae | Chlorella sp. | Allmicroalgae, Natural Products | 5.00 | 12.6 | 4.30 | 7.43 | Finishing pigs | [167] |
| Microalgae | Chlorella sp. | Allmicroalgae, Natural Products | 10.0 | 5.44 | −3.48 | 3.14 | Broilers | [168] |
| Microalgae | Desmodesmus sp. | DGM, Cellana | 10.0 | −11.4 | −9.73 | −1.64 | Nursery pigs | [134] |
| 15.0 | 5.21 | 8.56 | 16.42 | Broilers | ||||
| Microalgae | Desmodesmus sp. | Pure, whole | 5.00 | 35.36 | −0.24 | 12.24 | Broilers | [179] |
| Pure, defatted | 5.00 | 20.91 | −2.37 | 5.44 | ||||
| Microalgae | Nannochloropsis oceanica | DGA, Cellana | 2.00 | 0.84 | −1.67 | 1.54 | Broilers | [169] |
| 4.00 | −2.39 | 0.83 | −3.08 | |||||
| 8.00 | 1.97 | 3.33 | 0.00 | |||||
| 16.0 | −10.4 | −3.33 | −6.15 | |||||
| Microalgae | Staurosira sp. | Pure, defatted | 7.50 | 1.20 | 6.21 | −4.78 | Broilers | [51] |
| Poultry Fat | Soybean Oil | Yarrowia lipolytica | Schizochytrium sp. | Crypthecodinium cohnii | |
|---|---|---|---|---|---|
| ME, kcal/kg | 8364 | 8574 | - | - | - |
| Total saturated, % | 28.7 | 14.2 | 19.4 | 36.5 | - |
| Total unsaturated, % | 64.8 | 81.0 | 80.6 | 62.4 | - |
| FA, % | - | ||||
| C 14:0 | 0.9 | 0.1 | 0.3 | 11.0 | 16.0 |
| C 16:0 | 21.6 | 10.3 | 10.7 | 38.5 | 25.0 |
| C 16:1 | 5.7 | 0.2 | 1.5 | 18.5 | 0.4 |
| C 18:0 | 6.0 | 3.8 | 6.6 | 1.10 | - |
| C 18:1 | 37.4 | 22.8 | 8.8 | 3.15 | 16.0 |
| C 18:2 | 19.5 | 51.0 | 22.9 | - | 0.5 |
| C 18:3 | 1.0 | 6.8 | 2.3 | - | 0.4 |
| C 20:0 | - | - | 0.7 | - | - |
| C 20:1 | 1.1 | 0.2 | 0.2 | 0.60 | - |
| C 20:4 | 0.1 | 0.0 | 4.0 | - | - |
| C 20:5 | 0.0 | 0.0 | 30.2 | 1.65 | 0.1 |
| C 22:1 | 0.0 | 0.0 | 0.9 | 0.10 | - |
| C 22:5 | 0.0 | 0.0 | 0.9 | 12.9 | - |
| C 22:6 | 0.0 | 0.0 | - | 24.0 | 39.0 |
| References | [137] | [137] | [206] | [208,209] | [207] |
| Microorganism | Species | Level, % | Improvement, % | Animal Model | Reference | ||
|---|---|---|---|---|---|---|---|
| ADG | ADFI | FE | |||||
| Yeast | Yarrowia lipolytica | 1.50 | 15.2 | −5.08 | 20.0 | Nursery pigs | [213] |
| 3.00 | 4.64 | −3.23 | 7.27 | ||||
| Microalgae | Aurantiochytrium acetophilum | 1.00 | −2.65 | 6.42 | −8.18 | Broilers | [215] |
| 2.00 | −4.05 | 1.83 | −6.21 | ||||
| 4.00 | −12.7 | 1.83 | −14.7 | ||||
| Microalgae | Schizochytrium sp. | 3.12 | −0.23 | −3.69 | 4.35 | Nursery pigs | [216] |
| Microalgae | Schizochytrium sp. | 3.60 | −1.92 | −0.95 | −0.86 | Growing-finishing pigs | [217] |
| Microalgae | Schizochytrium sp. | 3.70 | −0.93 | −3.03 | 3.45 | Growing-finishing pigs | [218] |
| Microalgae | Schizochytrium sp. | 0.25 | 4.17 | 0.65 | 3.38 | Growing-finishing pigs | [116] |
| 0.50 | 4.17 | 1.18 | 3.80 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Y.-C.; Kim, S.W. Use of Microorganisms as Nutritional and Functional Feedstuffs for Nursery Pigs and Broilers. Animals 2022, 12, 3141. https://doi.org/10.3390/ani12223141
Cheng Y-C, Kim SW. Use of Microorganisms as Nutritional and Functional Feedstuffs for Nursery Pigs and Broilers. Animals. 2022; 12(22):3141. https://doi.org/10.3390/ani12223141
Chicago/Turabian StyleCheng, Yi-Chi, and Sung Woo Kim. 2022. "Use of Microorganisms as Nutritional and Functional Feedstuffs for Nursery Pigs and Broilers" Animals 12, no. 22: 3141. https://doi.org/10.3390/ani12223141