
Gruel Creep Feeding Accelerates Growth and Alters Intestinal Health of Young Pigs
 , , , , , and
, , , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Cytokine Analysis
2.3. Histology
2.4. Gas Chromatography
2.5. Statistics
3. Results
3.1. ADG and Feed Intake
3.2. Intestinal Measures
3.3. VFA
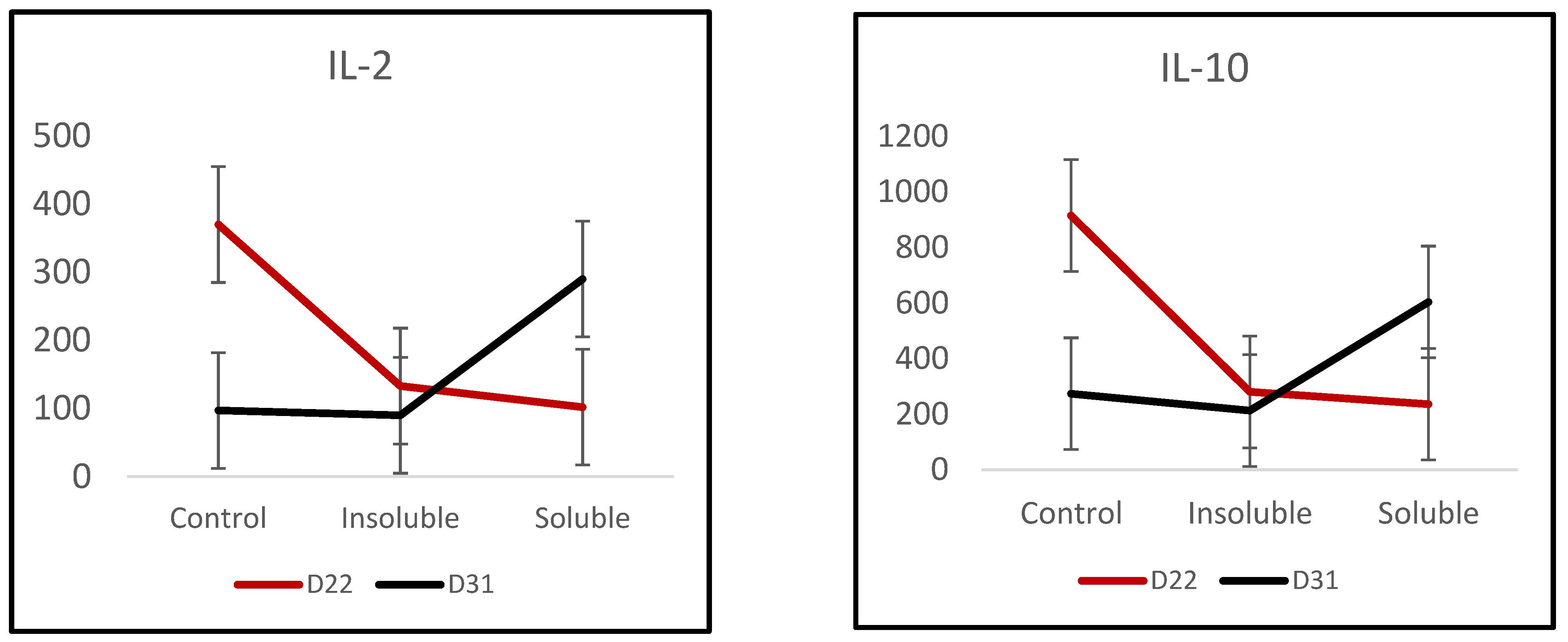
3.4. Cytokines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Beers-Schreurs, H.M.G.; Nabuurs, M.J.A.; Vellenga, L.; Valk, H.J.K.-V.D.; Wensing, T.; Breukink, H.J. Weaning and the Weanling Diet Influence the Villous Height and Crypt Depth in the Small Intestine of Pigs and Alter the Concentrations of Short-Chain Fatty Acids in the Large Intestine and Blood. J. Nutr. 1998, 128, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Cera, K.R.; Mahan, D.C.; Cross, R.F.; Reinhart, G.A.; Whitmoyer, R.E. Effect of age, weaning and postweaning diet on small intestinal growth and jejunal morphology in young swine. J. Anim. Sci. 1988, 66, 574–584. [Google Scholar] [CrossRef] [PubMed]
- De Greeff, A.; Resink, J.W.; van Hees, H.M.; Ruuls, L.; Klaassen, G.J.; Rouwers, S.M.; Stockhofe-Zurwieden, N. Supplementation of piglets with nutrient-dense complex milk replacer improves intestinal development and microbial fermentation. J. Anim. Sci. 2016, 94, 1012–1019. [Google Scholar] [CrossRef]
- Kobek-Kjeldager, C.; Moustsen, V.A.; Theil, P.K.; Pedersen, L.J. Effect of large litter size and within-litter differences in piglet weight on the use of milk replacer in litters from hyper-prolific sows under two housing conditions. Appl. Anim. Behav. Sci. 2020, 230, 9. [Google Scholar] [CrossRef]
- Willard, N.C.; Moest, N.K.; Vande Pol, K.D.; Olivio, A.; Shull, C.M.; McKilligan, D.; Ellis, M. 3 Effect of Litter Size and Provision of Supplementary Liquid Milk Replacer During Lactation on Piglet Pre-Weaning Performance. J. Anim. Sci. 2022, 100, 2–3. [Google Scholar] [CrossRef]
- Bruininx, E.M.; Binnendijk, G.P.; van der Peet-Schwering, C.M.; Schrama, J.W.; den Hartog, L.A.; Everts, H.; Beynen, A.C. Effect of creep feed consumption on individual feed intake characteristics and performance of group-housed weanling pigs. J. Anim. Sci. 2002, 80, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Santi-Devi, U. The Impact of Weaning Stress on Gut Health and the Mechanistic Aspects of Several Feed Additives Contributing to Improved Gut Health Function in Weanling Piglets—A Review. Animals 2021, 11, 2418. [Google Scholar] [CrossRef]
- Lee, S.I.; Kim, I.H. Creep feeding improves growth performance of suckling piglets. Rev. Bras. De Zootec. Braz. J. Anim. Sci. 2018, 47. [Google Scholar] [CrossRef]
- Byrgesen, N.; Madsen, J.G.; Larsen, C.; Kjeldsen, N.J.; Cilieborg, M.S.; Amdi, C. The Effect of Feeding Liquid or Dry Creep Feed on Growth Performance, Feed Disappearance, Enzyme Activity and Number of Eaters in Suckling Piglets. Animals 2021, 11, 3144. [Google Scholar] [CrossRef]
- Middelkoop, A.; Costermans, N.; Kemp, B.; Bolhuis, J.E. Feed intake of the sow and playful creep feeding of piglets influence piglet behaviour and performance before and after weaning. Sci. Rep. 2019, 9, 13. [Google Scholar] [CrossRef] [Green Version]
- Sulabo, R.C.; Tokach, M.D.; DeRouchey, J.M.; Dritz, S.S.; Goodband, R.D.; Nelssen, J.L. Effects of creep feeder design and feed accessibility on preweaning pig performance and the proportion of pigs consuming creep feed. J. Swine. Health Prod. 2010, 18, 174–181. [Google Scholar] [CrossRef]
- Heo, P.S.; Kim, D.H.; Jang, J.C.; Hong, J.S.; Kim, Y.Y. Effects of different creep feed types on pre-weaning and post-weaning performance and gut development. Asian Australas. J. Anim. Sci. 2018, 31, 1956–1962. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Duarte, M.E.; Sevarolli Loftus, A.; Kim, S.W. Intestinal Health of Pigs Upon Weaning: Challenges and Nutritional Intervention. Front. Vet. Sci. 2021, 8, 628258. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.A.; Parker, K.; Calvert, C.C.; Mills, D.A. Diet shapes the gut microbiome of pigs during nursing and weaning. Microbiome 2015, 3, 10. [Google Scholar] [CrossRef]
- Van Hees, H.M.J.; Davids, M.; Maes, D.; Millet, S.; Possemiers, S.; den Hartog, L.A.; van Kempen, T.; Janssens, G.P.J. Dietary fibre enrichment of supplemental feed modulates the development of the intestinal tract in suckling piglets. J. Anim. Sci. Biotechnol. 2019, 10, 11. [Google Scholar] [CrossRef]
- Zhang, C.J.; Yu, M.; Yang, Y.X.; Mu, C.L.; Su, Y.; Zhu, W.Y. Effect of early antibiotic intervention on specific bacterial communities and immune parameters in the small intestine of growing pigs fed different protein level diets. Animal 2020, 14, 2042–2053. [Google Scholar] [CrossRef]
- Kuller, W.I.; van Beers-Schreurs, H.M.; Soede, N.M.; Taverne, M.A.; Kemp, B.; Verheijden, J.H. Addition of chromic oxide to creep feed as a fecal marker for selection of creep feed-eating suckling pigs. Am. J. Vet. Res. 2007, 68, 748–752. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Duarte, M.E.; da Silva, S.N.; Kim, S.W. Supplemental Effects of Functional Oils on the Modulation of Mucosa-Associated Microbiota, Intestinal Health, and Growth Performance of Nursery Pigs. Animals 2021, 11, 1591. [Google Scholar] [CrossRef]
- Hess, H.A.; Corl, B.A.; Lin, X.; Jacobi, S.K.; Harrell, R.J.; Blikslager, A.T.; Odle, J. Enrichment of intestinal mucosal phospholipids with arachidonic and eicosapentaenoic acids fed to suckling piglets is dose and time dependent. J. Nutr. 2008, 138, 2164–2171. [Google Scholar] [CrossRef]
- Weigand, E.; Young, J.W.; McGilliard, A.D. Volatile fatty acid metabolism by rumen mucosa from cattle fed hay or grain. J. Dairy Sci. 1975, 58, 1294–1300. [Google Scholar] [CrossRef]
- Fraser, D.; Feddes, J.J.R.; Pajor, E.A. The relationship between creep feeding-behavior of piglets and adaptation to weaning—Effect of diet quality. Can. J. Anim. Sci. 1994, 74, 1–6. [Google Scholar] [CrossRef]
- Cabrera, R.A.; Usry, J.L.; Arrellano, C.; Nogueira, E.T.; Kutschenko, M.; Moeser, A.J.; Odle, J. Effects of creep feeding and supplemental glutamine or glutamine plus glutamate (Aminogut) on pre- and post-weaning growth performance and intestinal health of piglets. J. Anim. Sci. Biotechnol. 2013, 4, 29. [Google Scholar] [CrossRef] [PubMed]
- Toplis, P.; Blanchard, P.J.; Miller, H.M. Creep feed offered as a gruel prior to weaning enhances performance of weaned piglets. In Proceedings of the 7th Biennial Conference of the Australasian-Pig-Science-Association, Adelaide, Australia, 28 November–1 December 1999; p. 129. [Google Scholar]
- Figueroa, J.; Marchant, I.; Morales, P.; Salazar, L.C. Do Prenatally-Conditioned Flavor Preferences Affect Consumption of Creep Feed by Piglets? Animals 2019, 9, 944. [Google Scholar] [CrossRef]
- Azain, M.J.; Tomkins, T.; Sowinski, J.S.; Arentson, R.A.; Jewell, D.E. Effect of supplemental pig milk replacer on litter performance: Seasonal variation in response. J. Anim. Sci. 1996, 74, 2195–2202. [Google Scholar] [CrossRef]
- Kim, J.H.; Heo, K.N.; Odle, J.; Han, I.K.; Harrell, R.J. Liquid diets accelerate the growth of early-weaned pigs and the effects are maintained to market weight. J. Anim. Sci. 2001, 79, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Price, K.L.; Lin, X.; van Heugten, E.; Odle, R.; Willis, G.; Odle, J. Diet physical form, fatty acid chain length, and emulsification alter fat utilization and growth of newly weaned pigs1. J. Anim. Sci. 2013, 91, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Cappai, M.G.; Dall’Aglio, C.; Sander, S.J.; Ratert, C.; Dimauro, C.; Pinna, W.; Kamphues, J. Different physical forms of one diet fed to growing pigs induce morphological changes in mandubular glands and local leptin (Ob) production and receptor (ObR) expression. J. Anim. Physiol. Anim. Nutr. 2016, 100, 1067–1072. [Google Scholar] [CrossRef]
- Cappai, M.G.; Arlinghaus, M.; Sander, S.J.; Dimauro, C.; Pinna, W.; Kamphues, J. Effect of the physical form of the diet on the morphometric traits of terminal ileum and ileocecal valve for potential prevention of small bowel retrograde contamination in the growing pig. Anim. Feed Sci. Technol. 2015, 210, 184–189. [Google Scholar] [CrossRef]
- Christensen, B.; Huber, L.A. The effect of creep feed composition and form on pre- and post-weaning growth performance of pigs and the utilization of low-complexity nursery diets. Transl. Anim. Sci. 2021, 5, txab211. [Google Scholar] [CrossRef]
- Zijlstra, R.T.; Whang, K.-Y.; Easter, R.A.; Odle, J. Effect of feeding a milk replacer to early-weaned pigs on growth, body composition, and small intestinal morphology, compared with suckled littermates2. J. Anim. Sci. 1996, 74, 2948–2959. [Google Scholar] [CrossRef] [Green Version]
- Muns, R.; Magowan, E. The effect of creep feed intake and starter diet allowance on piglets’ gut structure and growth performance after weaning. J. Anim. Sci. 2018, 96, 3815–3823. [Google Scholar] [CrossRef] [PubMed]
- Pluske, J.R.; Hampson, D.J.; Williams, I.H. Factors influencing the structure and function of the small intestine in the weaned pig: A review. Livest. Prod. Sci. 1997, 51, 215–236. [Google Scholar] [CrossRef]
- Holtug, K.; Rasmussen, H.S.; Mortensen, P.B. An in vitro study of short-chain fatty acid concentrations, production and absorption in pig (Sus scrofa) colon. Comp. Biochem. Physiol. Part A Physiol. 1992, 103, 189–197. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Wang, K.; Cao, G.; Zhang, H.; Li, Q.; Yang, C. Effects of Clostridium butyricum and Enterococcus faecalis on growth performance, immune function, intestinal morphology, volatile fatty acids, and intestinal flora in a piglet model. Food Funct. 2019, 10, 7844–7854. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.E.; Kim, S.W. Intestinal microbiota and its interaction to intestinal health in nursery pigs. Anim. Nutr. 2022, 8, 169–184. [Google Scholar] [CrossRef]


{kind=link}
| Ingredient (%) | Insoluble 1 | Soluble 2 |
|---|---|---|
| Whey Permeate | 24.00 | 38.82 |
| Corn, yellow dent | 29.34 | − |
| Soybean Meal | 12.00 | − |
| Cookie Meal | 10.00 | − |
| Poultry Meal | 10.00 | − |
| Fish Meal | 4.00 | − |
| Blood Plasma 3 | 4.00 | − |
| Soy protein concentrate 4 | 4.00 | − |
| Poultry Fat | 1.00 | − |
| Soy protein isolate | − | 10.00 |
| Nonfat Dried Milk | − | 26.34 |
| Feeding Oatmeal | − | 20.00 |
| Dextrose | − | 2.23 |
| L-Lys 5 | 0.52 | 0.57 |
| DL-Met 6 | 0.25 | 0.41 |
| L-Thr 5 | 0.17 | 0.30 |
| L-Trp 5 | 0.02 | 0.12 |
| Limestone | 0.30 | 0.40 |
| Salt | 0.22 | 1.0 |
| Mineral Premix 7 | 0.15 | − |
| Vitamin Premix 8 | 0.03 | − |
| Other 9 | − | 0.80 |
| Calculated composition | ||
| DM, % | 91.4 | 95.1 |
| ME, kcal/kg | 3465 | 3132 |
| CP, % | 25.17 | 23.13 |
| Lactose, % | 19.2 | 46.5 |
| SID Lys, % | 1.51 | 1.70 |
| Ca, % | 0.85 | 0.67 |
| p, % | 0.73 | 0.68 |
| ----------------Treatment------------------ | |||||
|---|---|---|---|---|---|
| Control | Insoluble | Soluble | SEM | p > F | |
| Number of litters | 6 | 6 | 6 | ||
| Initial Body Weight (kg/pig) | |||||
| Day 7 2 | 2.27 | 2.51 | 2.68 | 0.153 | 0.213 |
| Day 14 | 3.63 a | 3.85 a | 4.62 b | 0.262 | 0.043 |
| Day 23 | 5.51 a | 6.33 a | 7.65 b | 0.369 | 0.003 |
| Day 31 | 6.58 a | 7.03 a | 8.21 b | 0.362 | 0.017 |
| Average Daily Gain (g/pig) | |||||
| Day 7–14 | 193 a | 192 a | 278 b | 20 | 0.010 |
| Day 14–23 | 208 a | 275 b | 337 c | 18 | 0.001 |
| Day 23–31 | 153 | 100 | 79 | 25 | 0.134 |
| Day 7–31 | 187 a | 197 a | 240 b | 11 | 0.006 |
| Average daily feed intake (g/litter) | |||||
| Day 7–14 | 0.00 a | 143 b | 294 c | 27 | 0.001 |
| Day 14–23 | 0.00 a | 533 b | 935 c | 81 | 0.001 |
| Day 23–31 | 1891 | 1686 | 1911 | 216 | 0.723 |
| Average daily feed intake (g/pig) | |||||
| Day 7–14 | 0.00 a | 13 b | 28 c | 2.5 | 0.001 |
| Day 14–23 | 0.00 a | 50 b | 91 c | 7.6 | 0.001 |
| Day 23–31 | 186 | 185 | 210 | 15.6 | 0.460 |
| Gain:Feed | |||||
| Day 23–31 | 0.82 a | 0.57 a,b | 0.35 b | 0.12 | 0.045 |
| ------------------------Treatment----------------------- | -----------p > F------------- | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Items | Control | Insoluble | Soluble | SEM | Trt 2 | Age | T × A 3 | |||
| D22 | D31 | D22 | D31 | D22 | D31 | |||||
| Cecal pH | 6.33 b | 6.01 a,b | 6.00 a,b | 5.65 a | 6.28 b | 5.77 a | 0.13 | 0.06 | 0.01 | 0.78 |
| Small Intestine (%) 4 | 4.04 a | 6.55 c | 4.40 a | 8.86 d | 4.79 a,b | 6.06 b,c | 0.45 | 0.01 | 0.01 | 0.01 |
| Large Intestine (%) | 2.22 a,b | 3.75 d | 2.38 a,b,c | 3.44 c,d | 1.85 a | 3.57 d | 0.61 | 0.61 | 0.01 | 0.50 |
| --------------------Treatment-------------------- | --------- p > F ---------- | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Items | Control | Insoluble | Soluble | SEM | Trt 2 | Age | T × A 3 | |||
| D22 | D31 | D22 | D31 | D22 | D31 | |||||
| Villi (μm) | 268 a,b | 262 a,b | 238 a,b | 291 a | 193 c | 230 b | 14.8 | 0.01 | 0.03 | 0.15 |
| Crypt (μm) | 90.8 b | 123.2 a | 90.6 b | 134.4 a | 84.6 b | 97.3 b | 6.36 | 0.01 | 0.01 | 0.06 |
| Villi:Crypt ratio | 3.0 a | 2.16 b | 2.65 a,b | 2.40 b | 2.34 b | 2.40 b | 0.19 | 0.54 | 0.02 | 0.08 |
| Villus area (μm2 × 10−2) | 776 a | 753 a | 592 b,c | 743 a | 482 c | 654 b | 69.6 | 0.03 | 0.09 | 0.34 |
| -----------------------Treatment-------------------- | ------------p > F---------- | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Items | Control | Insoluble | Soluble | SEM | Trt 2 | Age | T × A 3 | |||
| D22 | D31 | D22 | D31 | D22 | D31 | |||||
| Acetate | 62.7 a,b | 64.8 b | 60.4 a,b | 68.3 b | 52.8 a | 67.0 b | 3.6 | 0.41 | 0.01 | 0.27 |
| Propionate | 15.7 | 21.4 | 22.6 | 24.3 | 19.8 | 22.0 | 2.9 | 0.26 | 0.18 | 0.77 |
| Butyrate | 5.4 | 6.7 | 5.5 | 5.7 | 11.3 | 5.7 | 1.8 | 0.26 | 0.37 | 0.16 |
| Valerate | 3.0 | 1.1 | 1.6 | 1.1 | 4.1 | 2.4 | 1.1 | 0.26 | 0.17 | 0.80 |
| BCA 4 | 13.2 a | 5.9 b | 10.0 a | 0.55 c | 12.1 a | 3.0 b,c | 1.5 | 0.03 | 0.01 | 0.74 |
| Total 5 | 77.5 | 117.3 | 91.3 | 80.5 | 81.9 | 127.0 | 16.9 | 0.55 | 0.09 | 0.21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boston, T.E.; Wang, F.; Lin, X.; Leonard, S.; Kim, S.W.; McKilligan, D.; Fellner, V.; Odle, J. Gruel Creep Feeding Accelerates Growth and Alters Intestinal Health of Young Pigs. Animals 2022, 12, 2408. https://doi.org/10.3390/ani12182408
Boston TE, Wang F, Lin X, Leonard S, Kim SW, McKilligan D, Fellner V, Odle J. Gruel Creep Feeding Accelerates Growth and Alters Intestinal Health of Young Pigs. Animals. 2022; 12(18):2408. https://doi.org/10.3390/ani12182408
Chicago/Turabian StyleBoston, Timothy E., Feng Wang, Xi Lin, Suzanne Leonard, Sung Woo Kim, Denny McKilligan, Vivek Fellner, and Jack Odle. 2022. "Gruel Creep Feeding Accelerates Growth and Alters Intestinal Health of Young Pigs" Animals 12, no. 18: 2408. https://doi.org/10.3390/ani12182408