
Copy Number Variation of the SOX6 Gene and Its Associations with Growth Traits in Ashidan Yak

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Welfare
2.2. Body Size Traits and the Blood Samples Collection
2.3. Isolation and Identification of Genomic the DNA and RNA
2.4. Information of Candidate Gene
2.5. Primer Design
2.6. Copy Number Variation Identification and Gene Expression
2.7. CNV Correlation Analysis and Expression Profiling
3. Results
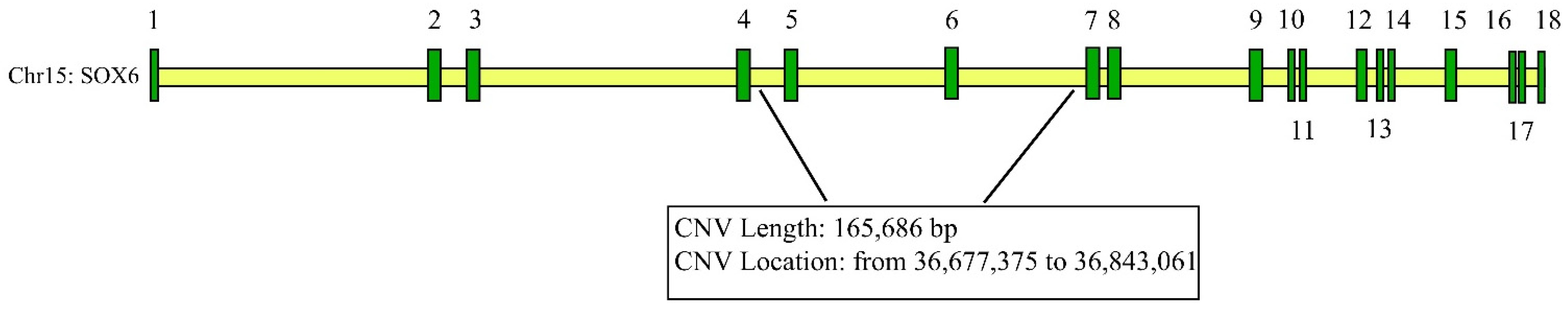
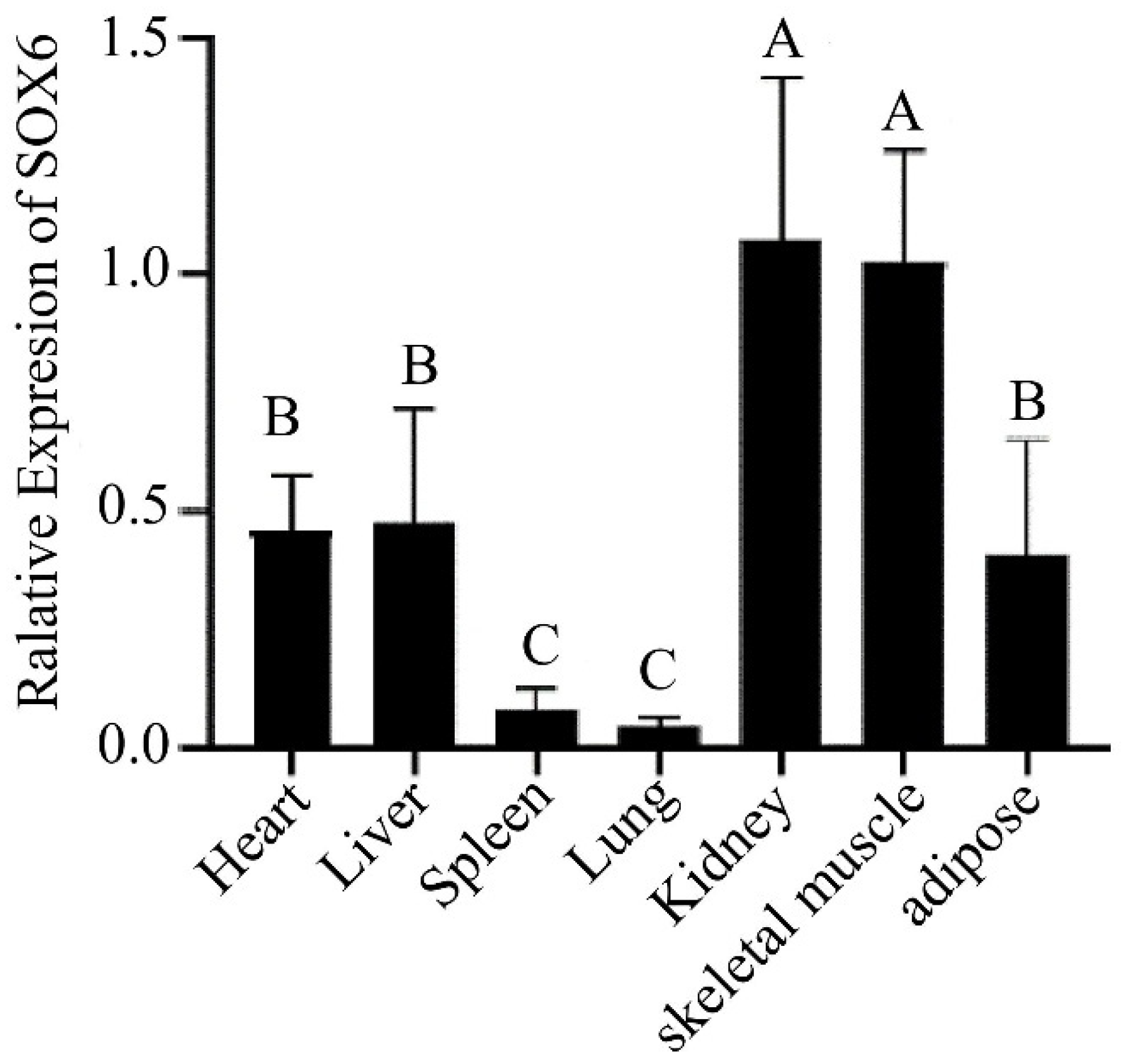
3.1. Expression Profiling of SOX6 Gene and Distribution of Different CNV Types in Yaks
3.2. Correlation between the SOX6-CNV and Four Growth Traits at Different Ages of 311 Yaks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feuk, L.; Marshall, C.R.; Scherer, W.S.W. Structural variants: Changing the landscape of chromosomes and design of disease studies. Hum. Mol. Genet. 2006, 15, R57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, S.Y.; Li, L.J.; Zhang, Z.J.; Wang, E.Y.; Huang, Y.Z. Copy number variation of MYLK4 gene and its growth traits of Capra hircus (goat). Anim. Biotechnol. 2019, 31, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Korbel, J.O.; Urban, A.E.; Affourtit, J.P.; Godwin, B.; Grubert, F.; Simons, J.F.; Kim, P.M.; Palejev, D.; Carriero, N.J.; Du, L. Paired-End Mapping Reveals Extensive Structural Variation in the Human Genome. Science 2007, 318, 420–426. [Google Scholar] [CrossRef] [Green Version]
- Rogers, R.L.; Cridland, J.M.; Shao, L.; Hu, T.T.; Peter, A.; Thornton, K.R. Landscape of standing variation for tandem duplications in Drosophila yakuba and Drosophila simulans. Mol. Biol. Evol. 2014, 37, 1750–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stothard, P.; Choi, J.; Basu, U.; Sumnerthomson, J.M.; Yan, M.; Liao, X.; Moore, S.S. Whole genome resequencing of Black Angus and Holstein cattle for SNP and CNV discovery. BMC Genom. 2011, 12, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iourov, I.Y.; Vorsanova, S.G.; Yurov, Y.B. The variome concept: Focus on CNVariome. Mol. Cytogenet. 2019, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Dorshorst, B.; Molin, A.M.; Rubin, C.J.; Johansson, A.M.; StrMstedt, L.; Pham, M.H.; Chen, C.F.; Hallb, F.K.; Shwell, C.A.; Andersson, L. A Complex Genomic Rearrangement Involving the Endothelin 3 Locus Causes Dermal Hyperpigmentation in the Chicken. PLoS Genet. 2011, 7, e1002412. [Google Scholar] [CrossRef] [Green Version]
- Seo, B.Y.; Park, E.W.; Ahn, S.J.; Lee, S.H.; Jeon, J.T. An accurate method for quantifying and analyzing copy number variation in porcine KIT by an oligonucleotide ligation assay. BMC Genet. 2007, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.C.; Cao, X.K.; He, H.; Song, L.F.; Huang, C.; Dang, L.P.; Zhang, J.Y.; Lei, C.Z.; Cheng, H.; Qi, X.L.; et al. The Detection of GBP4 Gene Copy Number Variation and Its Effect on Five Bovine Growth Traits. China Cattle Sci. 2016, 42, 9–12. [Google Scholar]
- Wright, D.; Boije, H.; Meadows, J.R.S.; Bed’hom, B.; Gourichon, D.; Vieaud, A.; Tixier-Boichard, M.; Rubin, C.J.; Imsland, F. Copy Number Variation in Intron 1 of SOX5 Causes the Pea-comb Phenotype in Chickens. PLoS Genet. 2009, 5, e1000512. [Google Scholar] [CrossRef]
- Ding, L.; Wang, Y.; Kreuzer, M.; Guo, X.; Mi, J.; Gou, Y.; Shang, Z.; Zhang, Y.; Zhou, J.; Wang, H.; et al. Seasonal variations in the fatty acid profile of milk from yaks grazing on the Qinghai-Tibetan plateau. J. Dairy Res. 2013, 80, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, L.Z.; Shi, T.; Zhou, Y.; Cai, H.F. Copy number variations of MICAL-L2 shaping gene expression contribute to different phenotypes of cattle. Mamm. Genome 2013, 24, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.M.; Li, Z.; He, H.; Song, C.C.; Zhang, Z.J. Associations of GBP2 gene copy number variations with growth traits and transcriptional expression in Chinese cattle. Gene 2018, 647, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Henrique, D.; Junior, G.; Cesar, A.; Freua, M.C.; Gomes, R.C.; Da, L.; Leme, P.R.; Fukumasu, H.; Carvalho, M.E.; Ventura, R.V. Copy number variations and genome-wide associations reveal putative genes and metabolic pathways involved with the feed conversion ratio in beef cattle. J. Appl. Genet. 2016, 57, 495–504. [Google Scholar]
- Kondoh, H.; Kamachi, Y. SOX-partner code for cell specification: Regulatory target selection and underlying molecular mechanisms. Int. J. Biochem. Cell Biol. 2009, 42, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [Green Version]
- Hamada-Kanazawa, M.; Ishikawa, K.; Ogawa, D.; Kanai, M.; Kawai, Y.; Narahara, M.; Miyake, M. Suppression of SOX6 in P19 cells leads to failure of neuronal differentiation by retinoic acid and induces retinoic acid-dependent apoptosis. FEBS Lett. 2004, 577, 60–66. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, V.; Crombrugghe, B.D.; Behringer, R.R. The transcription factors L-SOX5 and SOX6 are essential for cartilage formation. Dev. Cell 2001, 1, 277–290. [Google Scholar]
- Lefebvre, V. The SOXD transcription factors—SOX5, SOX6, and SOX13—Are key cell fate modulators. Int. J. Biochem. Cell Biol. 2010, 42, 429–432. [Google Scholar] [CrossRef] [Green Version]
- Jackson, H.E.; Ono, Y.; Wang, X.; Elworthy, S.; Cunliffe, V.T.; Ingham, P.W. The role of SOX6 in zebrafish muscle fiber type specification. Skelet. Muscle 2015, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.M. Studies on the Transcriptional Regulation Mechanisms of Transcription Factors Required for Skeletal Development; Northeast Normal University: Changchun, China, 2009. [Google Scholar]
- Aza-Carmona, M.; Shears, D.J.; Yuste-Checa, P.; Barca-Tierno, V.; Hisado-Oliva, A.; Belinchón, A.; Benito-Sanz, S.; Rodríguez, J.I.; Argente, J.; Campos-Barros, A.; et al. SHOX interacts with the chondrogenic transcription factors SOX5 and SOX6 to activate the aggrecan enhancer. Hum. Mol. Genet. 2011, 20, 1547–1559. [Google Scholar] [CrossRef] [PubMed]
- Klewer, H.S.E.; Samson, R.A. SOX6 is a candidate gene for p100H myopathy, heart block, and sudden neonatal death. Proc. Natl. Acad. Sci. USA 2000, 97, 4180–4185. [Google Scholar]
- Jia, C.; Wang, H.; Li, C.; Wu, X.; Yan, P. Genome-wide detection of copy number variations in polled yak using the Illumina BovineHD BeadChip. BMC Genom. 2019, 20, 376. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.P.; Bailey, D.R.; Shannon, N.H. Linear body measurements of cattle before and after 20 years of selection for postweaning gain when fed two different diets. J. Anim. Sci. 1993, 71, 1712. [Google Scholar] [CrossRef]
- Edwards, E.; Saunders, N.; Logan, J.; Sails, A.D.; Ad, S. Real-Time PCR: Current Technology and Applications; Caister Academic Press: Poole, UK, 2009; p. 284. [Google Scholar]
- Ali, S.; Srivastava, A.K.; Chopra, R.; Aggarwal, S.; Garg, V.K.; Bhattacharya, S.N.; Bamezai, R. IL12B SNPs and copy number variation in IL23R gene associated with susceptibility to leprosy. J. Med. Genet. 2013, 50, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.S.; Cheong, H.S.; Kim, L.H.; Namgung, S.; Park, T.J.; Chun, J.Y.; Kim, J.Y.; Pasaje, C.; Jin, S.L.; Shin, H.D. Identification of copy number variations and common deletion polymorphisms in cattle. BMC Genom. 2010, 11, 232. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, B.; Shi, T.; Huang, Y.; Liu, G.E.; Lan, X.; Lei, C.; Chen, H. Copy number variation of bovine SHH gene is associated with body conformation traits in Chinese beef cattle. J. Appl. Genet. 2019, 60, 199–207. [Google Scholar] [CrossRef]
- Knudsen, O. Studies on spermatocytogenesis in the bull. Int. J. Fertil. 1958, 3, 389–403. [Google Scholar]
- Yang, X.; Song, Z.; Wu, C.; Wang, W.; Li, G.; Zhang, W.; Wu, L.; Lu, K. Constructing a database for the relations between CNV and human genetic diseases via systematic text mining. BMC Bioinform. [Electron. Resour.] 2018, 19 (Suppl. 19), 528. [Google Scholar] [CrossRef]
- Ma, Y.L.; Wen, Y.F.; Cao, X.K.; Cheng, J.; Chen, H. Copy number variation (CNV) in the IGF1R gene across four cattle breeds and its association with economic traits. Arch. Anim. Breed. 2019, 62, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Jing, Z.; Wang, X.; Cheng, Y.; Wei, C.; Hou, D.; Li, T.; Li, W.; Han, R.; Li, H.; Sun, G. Detection of CNV in the SH3RF2 gene and its effects on growth and carcass traits in chickens. BMC Genet. 2020, 21, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.G.; Zhang, Y.B.; Yan, H.; Zhang, L.C.; Hou, X.H.; Liu, X.; Wang, L.X. Identification of Candidate Genes for Porcine Bone Rate Traitsby Genome-wide Association of Copy Number Variation. Acta Vet. Zootech. Sin. 2019, 50, 7. [Google Scholar]
- Huang, C.; Ge, F.; Ren, W.; Zhang, Y.; Liang, C. Copy Number Variation of the HPGDS Gene in the Ashidan yak and Its Associations with Growth Traits. Gene 2020, 772, 145382. [Google Scholar] [CrossRef]
- Ikeda, T.; Kawaguchi, H.; Kamekura, S.; Kou, I.; Hoshi, K.; Nakamura, K.; Ikegawa, S.; Chung, U. Combination of SOX5, SOX6 and SOX9 (the SOX trio) provides signals suffucient for introduction of permanent cartilage. Arthritis Rheum. 2004, 50, 3561–3573. [Google Scholar] [CrossRef] [PubMed]
- Frances, C.; Edwina, W.; Paul, D.; Peter, K.; Alan, A. The Sry-related HMG box-containing gene SOX6 is expressed in the adult testis and developing nervous system of the mouse. Nucleic Acids Res. 1995, 23, 3365. [Google Scholar]
- Cantone, M.; Küspert, M.; Reiprich, S.; Lai, X.; Eberhardt, M.; Gttle, P.; Beyer, F.; Azim, K.; Küry, P.; Wegner, M. A gene regulatory architecture that controls region-independent dynamics of oligodendrocyte differentiation. Glia 2019, 67, 825–843. [Google Scholar] [CrossRef]
- Zhang, L.; Xue, Z.; Yan, J.; Wang, J.; Liu, Q.; Jiang, H. LncRNA Riken-201 and Riken-203 modulates neural development by regulating the SOX6 through sequestering miRNAs. Cell Prolif. 2019, 52, e12573. [Google Scholar] [CrossRef]
- Sluijter, J.; Mil, A.V.; Vliet, P.V.; Metz, C.; Liu, J.; Doeven Da Ns, P.A.; Goumans, M.J. MicroRNA-1 and -499 regulate differentiation and proliferation in human-derived cardiomyocyte progenitor cells. Arter. Thromb. Vasc. Biol 2010, 30, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Lübbert, M.; Jones, P.A. Epigenetic Regulation of Globin Genes and Disturbances in Hemoglobinopathies; Springer: Berlin/Heidelberg, Germany, 2014; pp. 89–106. [Google Scholar] [CrossRef] [Green Version]
- An, C.I.; Dong, Y.; Nobuko, H. Genome-wide mapping of SOX6 binding sites in skeletal muscle reveals both direct and indirect regulation of muscle terminal differentiation by SOX6. BMC Dev. Biol. 2011, 11, 59. [Google Scholar] [CrossRef] [Green Version]
- Rooij, E.V.; Quiat, D.; Johnson, B.A.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Kelm, R.J.; Olson, E.N. A family of microRNAs encoded by myosin genes governs myosin expression and muscle performance. Dev. Cell 2009, 17, 662–673. [Google Scholar] [CrossRef] [Green Version]
- Smits, P. SOX5 and SOX6 are needed to develop and maintain source, columnar, and hypertrophic chondrocytes in the cartilage growth plate. J. Cell Biol. 2004, 164, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Quiat, D.; Voelker, K.A.; Pei, J.; Grishin, N.V.; Grange, R.W.; Bassel-Duby, R.; Olson, E.N. Concerted regulation of myofiber-specific gene expression and muscle performance by the transcriptional repressor SOX6. Proc. Natl. Acad. Sci. USA 2011, 108, 10196–10201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Y. MicroRNA-499-5p Regulates the Formation of Porcine Slow Myofibers by Targeting SOX6. Master’s Thesis, Sichuan Agricultural University, Ya’an, China, 2017. [Google Scholar]
- Wiener, G.; Han, J.; Long, R. The yak. Rap Publ. 2011, 44, 57–58. [Google Scholar]
- Qiu, Q.; Zhang, G.; Ma, T.; Qian, W.; Wang, J. The yak genome and adaptation to life at high altitude. Nat. Genet. 2012, 44, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.F. Study on Main Factors Influencing the Growth of Yak under the Condition of Grazing Grass at Different Growing Times during Warm Season. China Dairy Cattle 2021, 5–8. [Google Scholar] [CrossRef]
- Lin, X.R. Copy Number Variation of the SOX6 Affects Chicken Growth Traits. Master’s Thesis, South China Agricultural University, Guangzhou, China, 2016. [Google Scholar]



{kind=link}
{kind=link}
| Reagents and Instruments | Manufacturer |
|---|---|
| EasyPure Blood Genomic DNA Kit | TransGen Biotech, Beijing, China |
| Trizol reagent | TransGen Biotech, Beijing, China |
| PrimeScript™ Reagent Kit and gDNA Eraser | TaKaRa Bio Inc., Dalian, China |
| RNase-free DNase | TaKaRa Bio Inc., Dalian, China |
| Thermo Scientific NanoDrop 2000C | ThermoFisher Scientific Inc., Waltham, MA, USA |
| LightCycler® 96 Instrument | Roche, Basel, Switzerland |
| Level | Gene | Primer Sequence (5′–3′) | Product Length (bp) | Tm (°C) |
|---|---|---|---|---|
| DNA | SOX6 | F: GCAACTACCACACCGTCACCTC | 114 | 59 |
| R: TCCGCCGTCTGTCTTCATACCA | ||||
| DNA | BTF | F: AACCAGGAGAAACTCGCCAA | 166 | 59 |
| R: TTCGGTGAAATGCCCTCTCG | ||||
| mRNA | SOX6 | F: CGTTTGGGCAGGAGTTTGGA | 148 | 60 |
| R: CGTTTGGTGGCTGTGGAGTT | ||||
| mRNA | β-Actin | F: GCAGGTCATCACCATCGG | 158 | 60 |
| R: CCGTGTTGGCGTAGAGGT |
| Age | Growth Trait | CNV (Type Mean ± SE) | p Value | ||
|---|---|---|---|---|---|
| Loss (n = 17) | Normal (n = 182) | Gain (n = 112) | |||
| 6 months | Body weight (kg) | 83.82 ± 2.826 | 84.45 ± 0.762 | 83.89 ± 0.996 | 0.896 |
| Withers height (cm) | 94.94 ± 1.133 a | 94.80 ± 0.410 ab | 93.19 ± 0.475 b | 0.036 * | |
| Body length (cm) | 91.76 ± 1.868 | 91.61 ± 0.492 | 92.82 ± 0.422 | 0.829 | |
| Chest girth (cm) | 121.76 ± 2.006 b | 124.81 ± 0.569 a | 122.78 ± 0.731 b | 0.047 * | |
| 12 months | Body weight (kg) | 83.35 ± 3.240 | 83.49 ± 0.778 | 81.13 ± 1.028 | 0.185 |
| Withers height (cm) | 90.47 ± 1.275 | 90.77 ± 0.294 | 90.04 ± 0.408 | 0.348 | |
| Body length (cm) | 95.35 ± 1.228 | 96.27 ± 0.351 | 95.18 ± 0.459 | 0.156 | |
| Chest girth (cm) | 115.76 ± 1.675 | 117.64 ± 0.358 | 116.45 ± 0.511 | 0.090 | |
| 18 months | Body weight (kg) | 121.55 ± 4.857 | 123.67 ± 1.126 | 121.63 ± 1.308 | 0.487 |
| Withers height (cm) | 99.50 ± 1.863 b | 101.03 ± 0.513 b | 102.77 ± 0.537 a | 0.038 * | |
| Body length (cm) | 99.86 ± 1.522 | 102.32 ± 0.479 | 101.28 ± 0.519 | 0.160 | |
| Chest girth (cm) | 136.79 ± 2.358 | 137.56 ± 0.851 | 139.08 ± 0.850 | 0.434 | |
| 30 months | Body weight (kg) | 155.64 ± 3.827 | 155.50 ± 1.380 | 155.48 ± 1.625 | 0.999 |
| Withers height (cm) | 100.20 ± 1.277 | 100.08 ± 0.451 | 98.86 ± 0.544 | 0.208 | |
| Body length (cm) | 113.87 ± 1.591 | 113.81 ± 0.477 | 112.53 ± 0.638 | 0.257 | |
| Chest girth (cm) | 147.20 ± 2.066 ab | 148.10 ± 0.722 a | 145.14 ± 0.953 b | 0.045 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Huang, C.; Liu, M.; Dai, R.; Wu, X.; Ma, X.; Chu, M.; Bao, P.; Pei, J.; Guo, X.; et al. Copy Number Variation of the SOX6 Gene and Its Associations with Growth Traits in Ashidan Yak. Animals 2022, 12, 3074. https://doi.org/10.3390/ani12223074
Li X, Huang C, Liu M, Dai R, Wu X, Ma X, Chu M, Bao P, Pei J, Guo X, et al. Copy Number Variation of the SOX6 Gene and Its Associations with Growth Traits in Ashidan Yak. Animals. 2022; 12(22):3074. https://doi.org/10.3390/ani12223074
Chicago/Turabian StyleLi, Xinyi, Chun Huang, Modian Liu, Rongfeng Dai, Xiaoyun Wu, Xiaoming Ma, Min Chu, Pengjia Bao, Jie Pei, Xian Guo, and et al. 2022. "Copy Number Variation of the SOX6 Gene and Its Associations with Growth Traits in Ashidan Yak" Animals 12, no. 22: 3074. https://doi.org/10.3390/ani12223074