
Role of the Melanocortin System in Gonadal Steroidogenesis of Zebrafish
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Housing
2.2. Quantitative Real-Time PCR (qPCR)
2.3. Synthesis of Riboprobes
2.4. In Situ Hybridization
2.5. Primary Cell Cultures
2.5.1. Reagents
2.5.2. Isolation of Testicular and Ovarian Cells
2.5.3. In Vitro Treatments
2.5.4. Steroid Release Analysis
2.6. Data Analysis
3. Results
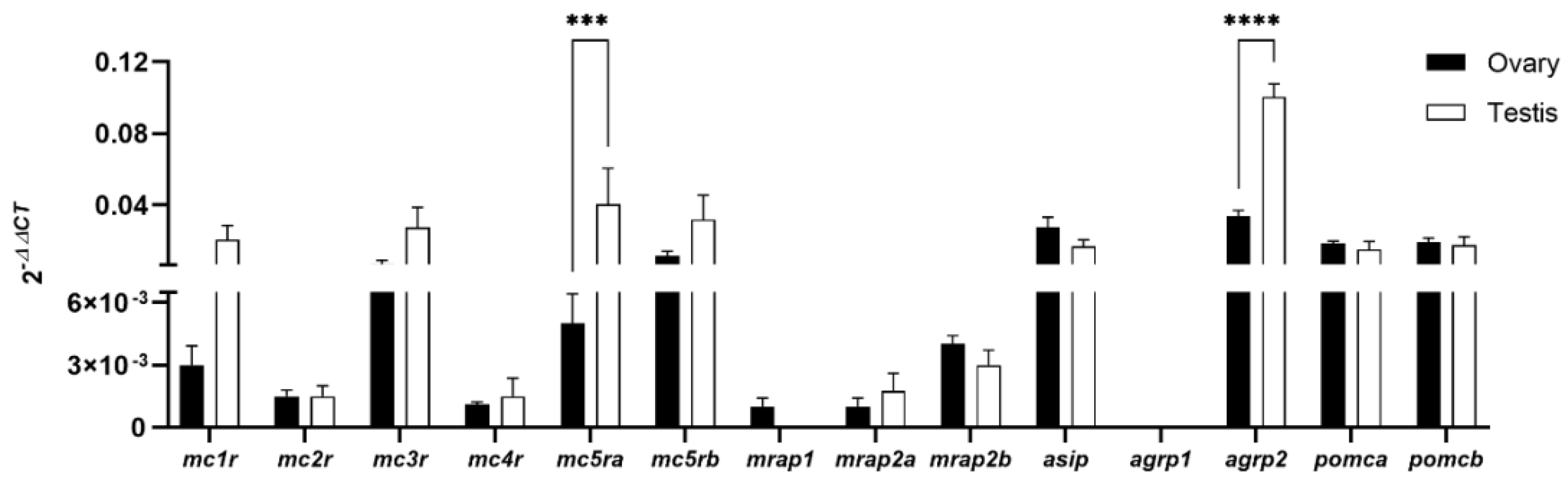
3.1. Expression of the Melanocortin System in Zebrafish Gonads
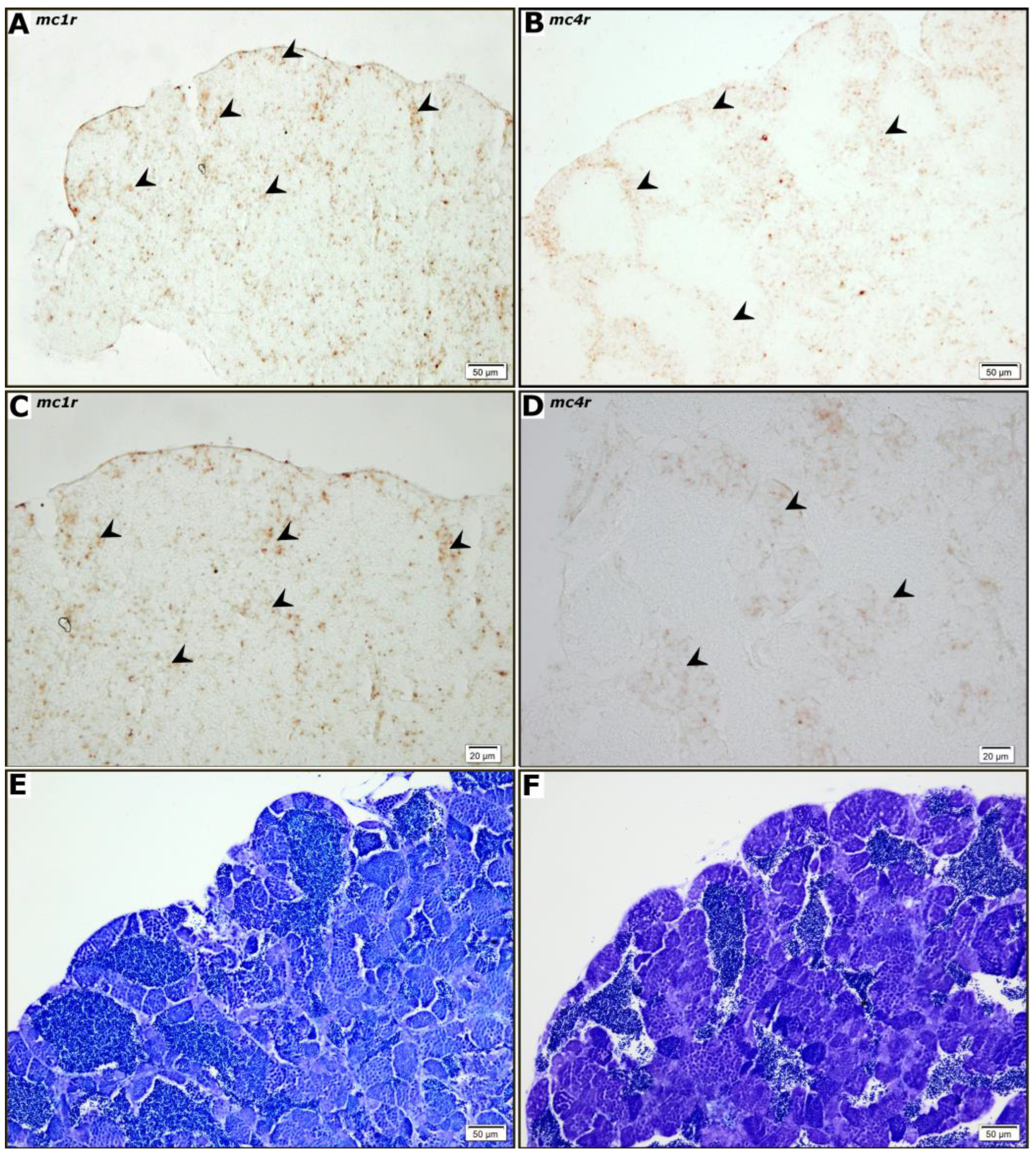
3.2. Cellular Localization of mc1r and mc4r mRNA in Ovaries and Testes
3.3. In Vitro Effects of Melanocortin Peptides on Basal and hCG-Induced Steroid Secretion by Zebrafish Gonadal Cells
3.3.1. Estradiol Secretion by Ovarian Follicles
3.3.2. Testosterone Secretion by Testicular Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerdá-Reverter, J.M.; Agulleiro, M.J.; Sánchez, E.; Guillot, R.; Ceinos, R.; Rotllant, J. Fish Melanocortin System. Eur. J. Pharmacol. 2011, 660, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortés, R.; Navarro, S.; Agulleiro, M.J.; Guillot, R.; García-Herranz, V.; Sánchez, E.; Cerdá-Reverter, J.M. Evolution of the melanocortin system. Gen. Comp. Endocrinol. 2014, 209, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Agulleiro, M.J.; Roy, S.; Sánchez, E.; Puchol, S.; Gallo-Payet, N.; Cerdá-Reverter, J.M. Role of melanocortin receptor accessory proteins in the function of zebrafish melanocortin receptor type 2. Mol. Cell. Endocrinol. 2010, 320, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Agulleiro, M.J.; Cortés, R.; Fernández-Durán, B.; Guillot, R.; Navarro, S.; Meimaridou, E.; Clark, A.J.; Cerdá-Reverter, J.M. Melanocortin 4 receptor becomes an ACTH receptor by coexpression of melanocortin receptor accessory protein 2. Mol. Endocrinol. 2013, 27, 1934–1945. [Google Scholar] [CrossRef]
- Navarro, S.; Guillot, R.; Crespo, D.; Schulz, R.W.; Ge, W.; Rotllant, J.; Cerda-Reverter, J.M.; Rocha, A. Enhanced growth without accelerated puberty in fish: A role for the melanocortin system. Aquaculture 2021, 540, 736721. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Agulleiro, M.J.; Cortés, R.; Sánchez, E.; Guillot, R.; Leal, E.; Fernández-Durán, B.; Puchol, S.; Eley, M. Involvement of MRAPs in the function of melanocortin receptors (MCRs). Gen. Comp. Endocrinol. 2013, 188, 133–136. [Google Scholar] [CrossRef]
- Quarta, C.; Claret, M.; Zeltser, L.M.; Williams, K.W.; Yeo, G.S.H.; Tschöp, M.H.; Diano, S.; Brüning, J.C.; Cota, D. POMC neuronal heterogeneity in energy balance and beyond: An integrated view. Nat. Metab. 2021, 3, 299–308. [Google Scholar] [CrossRef]
- Yeo, G.S.H.; Chao, D.H.M.; Siegert, A.M.; Koerperich, Z.M.; Ericson, M.D.; Simonds, S.E.; Larson, C.M.; Luquet, S.; Clarke, I.; Sharma, S.; et al. The melanocortin pathway and energy homeostasis: From discovery to obesity therapy. Mol. Metab. 2021, 48, 101206. [Google Scholar] [CrossRef]
- Russell, G.; Lightman, S. The human stress response. Nat. Rev. Endocrinol. 2019, 15, 525–534. [Google Scholar] [CrossRef] [Green Version]
- Gorissen, M.; Flik, G. The endocrinology of the stress response in fish an adaptation-physiological view. In Biology of Stress in Fish; Fish Physiology Series; Schreck, C.B., Tort, L., Farrell, A., Brauner, C., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 75–111. [Google Scholar]
- Cooray, S.N.; Clark, A.J. Melanocortin receptors and their accessory proteins. Mol. Cell. Endocrinol. 2011, 331, 215–221. [Google Scholar] [CrossRef]
- Schreck, C.B.; Tort, L. The concept of stress in fish. In Biology of Stress in Fish; Fish Physiology Series; Schreck, C.B., Tort, L., Farrell, A., Brauner, C., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 35, pp. 1–34. [Google Scholar]
- Soletto, L.; Hernández-Balfagó, S.; Rocha, A.; Scheerer, P.; Kleinau, G.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2-induced adrenocorticotropic hormone response of human melanocortin 4 receptor. J. Endocr. Soc. 2019, 3, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alsop, D.; Ings, J.S.; Vijayan, M.M. Adrenocorticotropic hormone suppresses gonadotropin-stimulated estradiol release from zebrafish ovarian follicles. PLoS ONE 2009, 4, e6463. [Google Scholar] [CrossRef] [PubMed]
- Cortés, R.; Agulleiro, M.J.; Navarro, S.; Guillot, R.; Sánchez, E.; Cerdá-Reverter, J.M. Melanocortin receptor accessory protein 2 (MRAP2) interplays with the zebrafish melanocortin 1 receptor (MC1R) but has no effect on its pharmacological profile. Gen. Comp. Endocrinol. 2014, 201, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Cortés, R.; Teles, M.; Oliveira, M.; Fierro-Castro, C.; Tort, L.; Cerdá-Reverter, J.M. Effects of acute handling stress on short-term central expression of orexigenic/anorexigenic genes in zebrafish. Fish Physiol. Biochem. 2018, 44, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Guillot, R.; Cortés, R.; Navarro, S.; Mischitelli, M.; García-Herranz, V.; Sánchez, E.; Cal, L.; Navarro, J.C.; Míguez, J.M.; Afanasyev, S.; et al. Behind melanocortin antagonist overexpression in the zebrafish brain: A behavioral and transcriptomic approach. Horm. Behav. 2016, 82, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Kurita, K.; Sakai, N. Functionally distinctive testicular cell lines of zebrafish to support male germ cell development. Mol. Reprod. Dev. 2004, 67, 430–438. [Google Scholar] [CrossRef]
- Pang, Y.; Ge, W. Epidermal growth factor and TGF-alpha promote zebrafish oocyte maturation in vitro: Potential role of the ovarian activin regulatory system. Endocrinology 2002, 143, 47–54. [Google Scholar] [CrossRef]
- Lampert, K.; Schmidt, C.; Fischer, P.; Volff, J.; Hoffmann, C.; Muck, J.; Lohse, M.; Ryan, M.; Schartl, M. Determination of onset of sexual maturation and mating behavior by melanocortin receptor 4 polymorphisms. Curr. Biol. 2010, 20, 1729–1734. [Google Scholar] [CrossRef]
- Liu, R.; Kinoshita, M.; Adolfi, M.C.; Schartl, M. Analysis of the role of the mc4r system in development, growth, and puberty of medaka. Front. Endocrinol. 2019, 10, 213. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wen, H.S.; Li, Y.; Lyu, L.K.; Zhang, Z.X.; Wang, X.J.; Li, J.S.; Tao, Y.X.; Qi, X. Melanocortin-4 receptor regulation of reproductive function in black rockfish (Sebastes schlegelii). Gene 2020, 741, 144541. [Google Scholar] [CrossRef] [PubMed]
- Thörnwall, M.; Dimitriou, A.; Xu, X.; Larsson, E.; Chhajlani, V. Immunohistochemical detection of the melanocortin 1 receptor in human testis, ovary and placenta using specific monoclonal antibody. Horm. Res. 1997, 48, 215–218. [Google Scholar] [CrossRef] [PubMed]
- Selz, Y.; Braasch, I.; Hoffmann, C.; Schmidt, C.; Schultheis, C.; Schartl, M.; Volff, J.N. Evolution of melanocortin receptors in teleost fish: The melanocortin type 1 receptor. Gene 2007, 401, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.Q.; Rao, Y.Z.; Zhang, Y.; Chen, R.; Tao, Y.X. Regulation of melanocortin-1 receptor pharmacology by melanocortin receptor accessory protein 2 in orange-spotted grouper (Epinephelus coioides). Gen. Comp. Endocrinol. 2020, 285, 113291. [Google Scholar] [CrossRef]
- Aluru, N.; Vijayan, M.M. Molecular characterization, tissue-specific expression, and regulation of melanocortin 2 receptor in rainbow trout. Endocrinology 2008, 149, 4577–4588. [Google Scholar] [CrossRef] [Green Version]
- Metz, J.R.; Geven, E.J.; van den Burg, E.H.; Flik, G. ACTH, alpha-MSH, and control of cortisol release: Cloning, sequencing, and functional expression of the melanocortin-2 and melanocortin-5 receptor in Cyprinus carpio. Am. J. Physiol. 2005, 289, R814–R826. [Google Scholar]
- Agulleiro, M.J.; Sánchez, E.; Leal, E.; Cortés, R.; Fernández-Durán, B.; Guillot, R.; Davis, P.; Dores, R.M.; Gallo-Payet, N.; Cerdá-Reverter, J.M. Molecular characterization and functional regulation of melanocortin 2 receptor (MC2R) in the sea bass. A putative role in the adaptation to stress. PLoS ONE 2013, 8, e65450. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Yu, H.; Mo, H.; Lan, X.; Pan, C.; Wang, L.; Zhao, H.; Zhou, J.; Li, Y. Functional Characterization of Melanocortin-3 Receptor in a Hibernating Cavefish Onychostoma macrolepis. Animals 2021, 12, 38. [Google Scholar] [CrossRef]
- Yu, H.X.; Li, Y.; Song, W.J.; Wang, H.; Mo, H.L.; Liu, Q.; Zhang, X.M.; Wang, L.X. Functional characterization of melanocortin-3 receptor in rainbow trout. Fish Physiol. Biochem. 2022, 48, 241–252. [Google Scholar] [CrossRef]
- Ji, R.L.; Huang, L.; Wang, Y.; Liu, T.; Fan, S.Y.; Tao, M.; Tao, Y.X. Topmouth culter melanocortin-3 receptor: Regulation by two isoforms of melanocortin-2 receptor accessory protein 2. Endocr. Connect. 2021, 10, 1489–1501. [Google Scholar] [CrossRef]
- Dowejko, M. Characterization of MC3 and the Other Melancortin Receptors (MC) in the Hypothalamo-Pituitary-Gonadal System of the Mouse. Ph.D. Thesis, University of Westminster, London, UK, 2014. Volume 262. [Google Scholar]
- O’Shaughnessy, P.J.; Fleming, L.M.; Jackson, G.; Hochgeschwender, U.; Reed, P.; Baker, P.J. Adrenocorticotropic hormone directly stimulates testosterone production by the fetal and neonatal mouse testis. Endocrinology 2003, 144, 3279–3284. [Google Scholar] [CrossRef] [PubMed]
- Gantz, I.; Fong, T.M. The melanocortin system. Am. J. Physiol. 2003, 284, E468–E474. [Google Scholar] [CrossRef] [PubMed]
- Haitina, T.; Klovins, J.; Andersson, J.; Fredriksson, R.; Lagerström, M.C.; Larhammar, D.; Larson, E.T.; Schiöth, H.B. Cloning, tissue distribution, pharmacology and three-dimensional modelling of melanocortin receptors 4 and 5 in rainbow trout suggest close evolutionary relationship of these subtypes. Biochem. J. 2004, 380, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Fathi, Z.; Iben, L.G.; Parker, E.M. Cloning, expression, and tissue distribution of a fifth melanocortin receptor subtype. Neurochem. Res. 1995, 20, 107–113. [Google Scholar] [CrossRef]
- Nimura, M.; Udagawa, J.; Hatta, T.; Hashimoto, R.; Otani, H. Spatial and temporal patterns of expression of melanocortin type 2 and 5 receptors the fetal mouse tissues and organs. Anat. Embryol. 2006, 211, 109–117. [Google Scholar] [CrossRef]
- Takeuchi, S.; Takahashi, S. Melanocortin receptor genes in the chicken—Tissue distributions. Gen. Comp. Endocrinol. 1998, 112, 220–231. [Google Scholar] [CrossRef]
- Ringholm, A.; Fredriksson, R.; Poliakova, N.; Yan, Y.L.; Postlethwait, J.H.; Larhammar, D.; Schiöth, H.B. One melanocortin 4 and two melanocortin 5 receptors from zebrafish show remarkable conservation in structure and pharmacology. J. Neurochem. 2002, 82, 6–18. [Google Scholar] [CrossRef]
- Chhajlani, V. Distribution of cDNA for melanocortin receptor subtypes in human tissues. Biochem. Mol. Biol. Int. 1996, 38, 73–80. [Google Scholar]
- Amweg, A.N.; Paredes, A.; Salvetti, N.R.; Lara, H.E.; Ortega, H.H. Expression of melanocortin receptors mRNA, and direct effects of ACTH on steroid secretion in the bovine ovary. Theriogenology 2011, 75, 628–637. [Google Scholar] [CrossRef]
- Chen, X.; Huang, L.; Tan, H.Y.; Li, H.; Wan, Y.; Cowley, M.; Veldhuis, J.D.; Chen, C. Deficient melanocortin-4 receptor causes abnormal reproductive neuroendocrine profile in female mice. Reproduction 2017, 153, 267–276. [Google Scholar] [CrossRef]
- Cerdá-Reverter, J.M.; Ringholm, A.; Schiöth, H.B.; Peter, R.E. Molecular cloning, pharmacological characterization, and brain mapping of the melanocortin 4 receptor in the goldfish: Involvement in the control of food intake. Endocrinology 2003, 144, 2336–2349. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, E.; Rubio, V.C.; Thompson, D.; Metz, J.; Flik, G.; Millhauser, G.L.; Cerda-Reverter, J.M. Phosphodiesterase inhibitor-dependent inverse agonism of agouti-related protein on melanocortin 4 receptor in sea bass (Dicentrarchus labrax). Am. J. Physiol. 2009, 296, R1293–R1306. [Google Scholar] [CrossRef]
- Li, J.T.; Yang, Z.; Chen, H.P.; Zhu, C.H.; Deng, S.P.; Li, G.L.; Tao, Y.X. Molecular cloning, tissue distribution, and pharmacological characterization of melanocortin-4 receptor in spotted scat, Scatophagus argus. Gen. Comp. Endocrinol. 2016, 230-231, 143–152. [Google Scholar] [CrossRef]
- Kim, J.Y.; Jung, H.J.; Yoon, M.J. VASA (DDX4) is a putative marker for spermatogonia, spermatocytes and round spermatids in stallions. Reprod. Domest. Anim. 2015, 50, 1032–1038. [Google Scholar] [CrossRef]
- Chen, S.X.J.; Schoonen, N.E.; Martijn, J.; de Waal, P.P.; Schulz, R.W. A progestin (17α,20β-dihydroxy-4-pregnen-3-one) stimulates early stages of spermatogenesis in zebrafish. Gen. Comp. Endocrinol. 2013, 185, 1–9. [Google Scholar] [CrossRef]
- Leal, M.C.; Cardoso, E.R.; Nóbrega, R.H.; Batlouni, S.R.; Bogerd, J.; França, L.R.; Schulz, R.W. Histological and stereological evaluation of zebrafish (Danio rerio) spermatogenesis with an emphasis on spermatogonial generations. Biol. Reprod. 2009, 81, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Facchinetti, F.; Polzonetti-Magni, A.; Neri, I.; Gallinelli, A.; Alessandrini, G.; Vallarino, M. Ovarian melanotropic peptides in two teleostean fishes. Ann. N. Y. Acad. Sci. 1993, 680, 502–504. [Google Scholar] [CrossRef]
- Mosconi, G.; Carnevali, O.; Facchinetti, F.; Radi, D.; Pestarino, M.; Vallarino, M.; Polzonetti-Magni, A.M. Ovarian melanotropic peptides and adaptation in two teleostean species: Sparus aurata L. and Dicentrarchus labrax L. Peptides 1994, 15, 927–931. [Google Scholar] [CrossRef]
- Masini, M.A.; Sturla, M.; Pestarino, M.; Gallinelli, A.; Facchinetti, F.; Uva, B.M. Pro-opiomelanocortin (POMC) expression and immunolocalization of POMC-related peptides in the ovary of Protopterus annectens, an African lungfish. Peptides 1997, 18, 1411–1414. [Google Scholar] [CrossRef]
- Wen, Z.Y.; Liu, T.; Qin, C.J.; Zou, Y.C.; Wang, J.; Li, R.; Tao, Y.X. MRAP2 Interaction with Melanocortin-4 Receptor in SnakeHead (Channa argus). Biomolecules 2021, 11, 481. [Google Scholar] [CrossRef]
- Rao, Y.Z.; Chen, R.; Zhang, Y.; Tao, Y.X. Orange-spotted grouper melanocortin-4 receptor: Modulation of signaling by MRAP2. Gen. Comp. Endocrinol. 2019, 284, 113234. [Google Scholar] [CrossRef] [PubMed]
- Tovo-Neto, A.; Martinez, E.; Melo, A.G.; Doretto, L.B.; Butzge, A.J.; Rodrigues, M.S.; Nakajima, R.T.; Habibi, H.R.; Nóbrega, R.H. Cortisol Directly Stimulates Spermatogonial Differentiation, Meiosis, and Spermiogenesis in Zebrafish (Danio rerio) Testicular Explants. Biomolecules 2020, 10, 429. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, Y.; Higuchi, M.; Miura, C.; Yamaguchi, S.; Tozawa, Y.; Miura, T. Roles of 11beta-hydroxysteroid dehydrogenase in fish spermatogenesis. Endocrinology 2006, 147, 5139–5146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandino, J.I.; Hattori, R.S.; Kishii, A.; Strüssmann, C.A.; Somoza, G.M. The cortisol and androgen pathways cross talk in high temperature-induced masculinization: The 11β-hydroxysteroid dehydrogenase as a key enzyme. Endocrinology 2012, 153, 6003–6011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kime, D.E. The hepatic catabolism of cortisol in teleost fish--adrenal origin of 11-oxotestosterone precursors. Gen. Comp. Endocrinol. 1978, 35, 322–328. [Google Scholar] [CrossRef]
- Schulz, R. In vitro metabolism of steroid hormones in the liver and in blood cells of male rainbow trout (Salmo gairdneri Richardson). Gen. Comp. Endocrinol. 1986, 64, 312–319. [Google Scholar] [CrossRef]
- Kumbar, J.; Ganesh, C.B. The effect of α-MSH treatment on the hypothalamic-pituitary-gonad axis in the cichlid fish Oreochromis mossambicus. Fish Physiol. Biochem. 2021, 47, 1659–1668. [Google Scholar] [CrossRef]
- Sánchez, E.; Rubio, V.C.; Cerdá-Reverter, J.M. Molecular and Pharmacological Characterization of the Melanocortin Receptor Subtype 1 in the Sea Bass. Gen. Comp. Endocrinol. 2010, 65, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, E.; Rubio, V.C.; Cerdá-Reverter, J.M. Characterization of the sea bass melanocortin 5 receptor: A putative role in hepatic lipid metabolism. J. Exp. Biol. 2009, 212, 3901–3910. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | References | Sequences (5′ → 3′) | Annealing T (°C) | Concentration |
|---|---|---|---|---|
| mc1r | [15] | F:TCCCACAAACCCTTACTGCAAG | 57 | 250 nM |
| R:TACACTGCAAAGCACCACGAAC | ||||
| mc2r | [4] | F:CCTGTTAGCACGCCATCATG | 60 | 300 nM |
| R:AGGCCGCTTTTCCTGTGTT | ||||
| P:[6FAM]AAACCGAATCGCGTCTATGCCTGGT[TAM] | ||||
| mc3r | Present work | F:TGTGATTGACCCGCTCATCTATG | 65 | 200 nM |
| R:TCTTCCCACATCCATTCTCAGTTC | ||||
| mc4r | [4] | F:GCCTCGCTCTACGTCCACAT | 60 | 300 nM |
| R:CGGCGATCCGTTTCATG | ||||
| P:[6FAM]TTCCTTCTAGCCCGGCTG[TAM] | ||||
| mc5ra | Present work | F:ATCATCTGCTGCTATAGTCTGA | 57 | 200 nM |
| R:ATCCACCGATCATATCCATCT | ||||
| mc5rb | Present work | F:CGCACTCAGGAGCCAAGAGATG | 65 | 200 nM |
| R:AGTTCCTCCAGGCACCTTCTTC | ||||
| mrap1 | [4] | F:CTTCTTCTTGATTTTGTCACTTATTTCAC | 60 | 300 nM |
| R:TCTTTACTGAGATGATGCATAACCTTTC | ||||
| P:[6FAM]CCCTCGAGTCAAAAAATCCGGTTTGC[TAM] | ||||
| mrap2a | [4] | F:AGAGCCGCCACTGATGCT | 60 | 300 nM |
| R:CCACTTGGCCTCTGGAGTTG | ||||
| P:[6FAM]CTCTCACCCATGGACGATCAGGCA[TAM] | ||||
| mrap2b | [4] | F:TTGGCTGTGAGCTGGAAGTG | 60 | 300 nM |
| R:TGAAAGAGGGAACGTGATTGG | ||||
| P:[6FAM]CATTTTCTCTGCCACCGCTGCCTG[TAM] | ||||
| pomca | [16] | F:AAATGACCCATTTCCGCTGGAG | 60 | 250 nM |
| R:CCCACCTTCGTTTCTATGCATG | ||||
| pomcb | [16] | F:AAACAACGGGAAGTATCGCATG | 60 | 250 nM |
| R:TCTGTGAACTGCTGTCCATTGC | ||||
| asip | [17] | F:CTGTGGGCGAGCTGCAAGAG | 60 | 250 nM |
| R:GCAGGGCTCCATAAACAGGAT | ||||
| agrp1 | [16] | F:GTGAATGTTGTGGTGATGG | 60 | 250 nM |
| R:TTCTTCTGCTGAGTTTATTTC | ||||
| agrp2 | [16] | F:GCTCTTCATCTGCTTGTTCTT | 55 | 250 nM |
| R:CTCCTGATTCCACACTCCT | ||||
| 18s | [15] | F:TGCATGGCCGTTCTTAGTTG | 60 | 150 nM |
| R:AGTCTCGTTCGTTATCGGAATGA |
| Mammals | Fish and Other Vertebrates | |||||
|---|---|---|---|---|---|---|
| Receptor | Organism | Organ/Tissue | Reference | Organism | Organ/Tissue | Reference |
| MC1R | Human | Leydig and corpus luteum cells | [24] | Platyfish, medaka and orange-spotted grouper | Testes and ovaries | [25,26] |
| Zebrafish | Testes | [2] | ||||
| MC2R | Mouse | Testes/Ovaries | [38] | Zebrafish, rainbow trout and carp | [3,14,27,28] | |
| Sea bass | Testes | [29] | ||||
| MC3R | Human | Testes and ovaries | [41] | |||
| Mouse | Hypothalamus-pituitary-gonad axis | [33] | ||||
| Mice | Fetal testes | [34] | Zebrafish, hibernating cavefish, rainbow trout and topmouth culter | Testes and ovaries | [30,31,32] | |
| Bovine | Granulosa and corpus luteum cells | [42] | ||||
| MC4R | Bovine | Antral follicle | [42] | Birds | Testes and ovaries | [39] |
| Mice | Ovaries | [43] | Goldfish, sea bass, spotted scat, medaka | Testes and ovaries | [22,44,45,46] | |
| Mice | Fetal testes | [34] | Zebrafish | Testes and ovaries | [4,36] | |
| MC5R | Mice | Testes | [37] | Birds | Testes and ovaries | [39] |
| Fetal mice | Spermatogonia and mesenchymal cells | [38] | Rainbow trout and zebrafish | Ovaries | [36,40] | |
| Zebrafish | Testes and ovaries | Present work | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro, S.; Crespo, D.; Schulz, R.W.; Ge, W.; Rotllant, J.; Cerdá-Reverter, J.M.; Rocha, A. Role of the Melanocortin System in Gonadal Steroidogenesis of Zebrafish. Animals 2022, 12, 2737. https://doi.org/10.3390/ani12202737
Navarro S, Crespo D, Schulz RW, Ge W, Rotllant J, Cerdá-Reverter JM, Rocha A. Role of the Melanocortin System in Gonadal Steroidogenesis of Zebrafish. Animals. 2022; 12(20):2737. https://doi.org/10.3390/ani12202737
Chicago/Turabian StyleNavarro, Sandra, Diego Crespo, Rüdiger W. Schulz, Wei Ge, Josep Rotllant, José Miguel Cerdá-Reverter, and Ana Rocha. 2022. "Role of the Melanocortin System in Gonadal Steroidogenesis of Zebrafish" Animals 12, no. 20: 2737. https://doi.org/10.3390/ani12202737