

Shedding Light on the Origin of Egyptian Sheep Breeds by Evolutionary Comparison of Mitochondrial D-Loop

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Sequencing
2.3. Construction of the Database
2.4. Phylogenetic and Data Analyses
3. Results
3.1. Distribution of Haplogroups in the Different Breeds
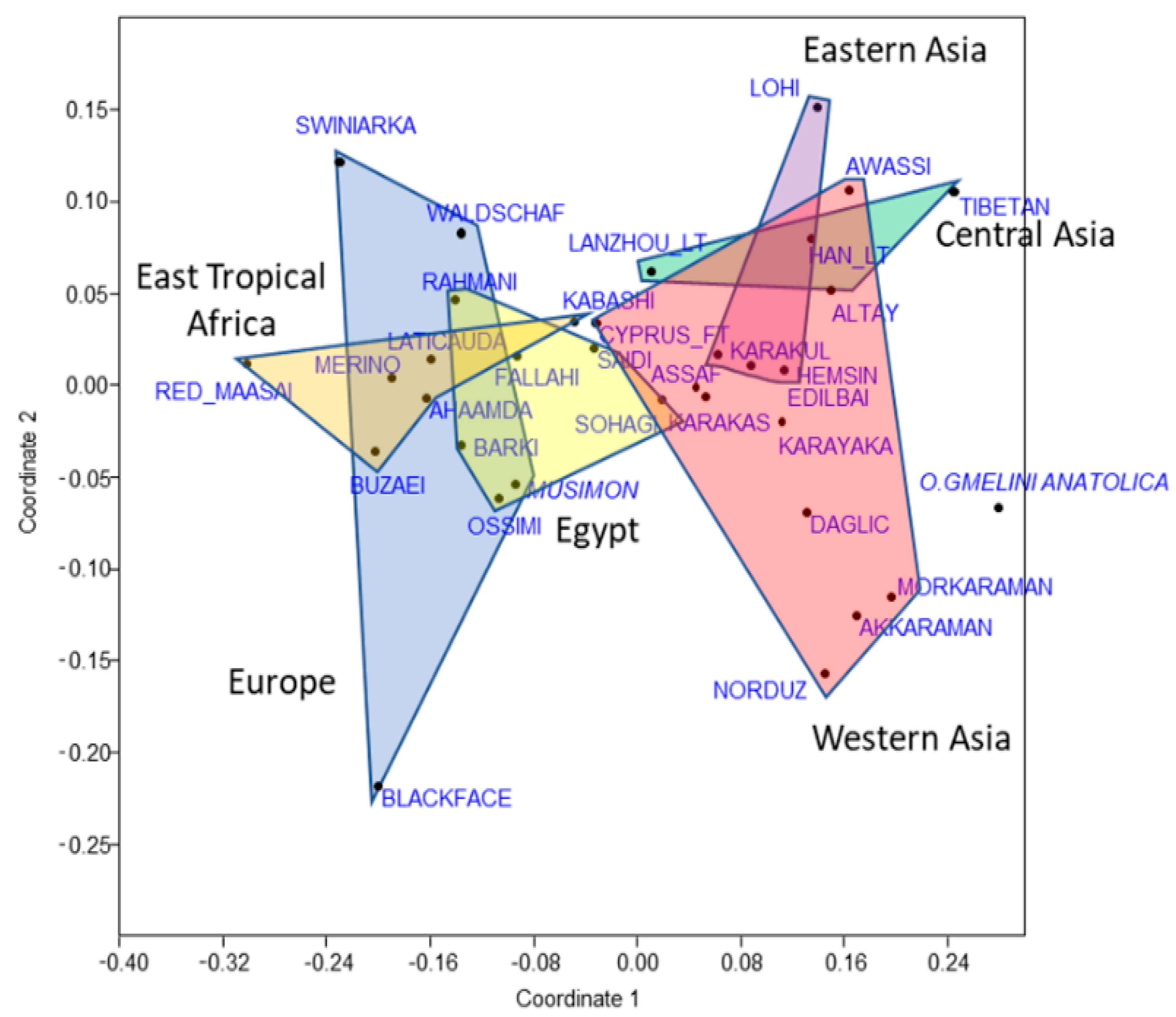
3.2. Geographical Distribution of Populations and Fst Values
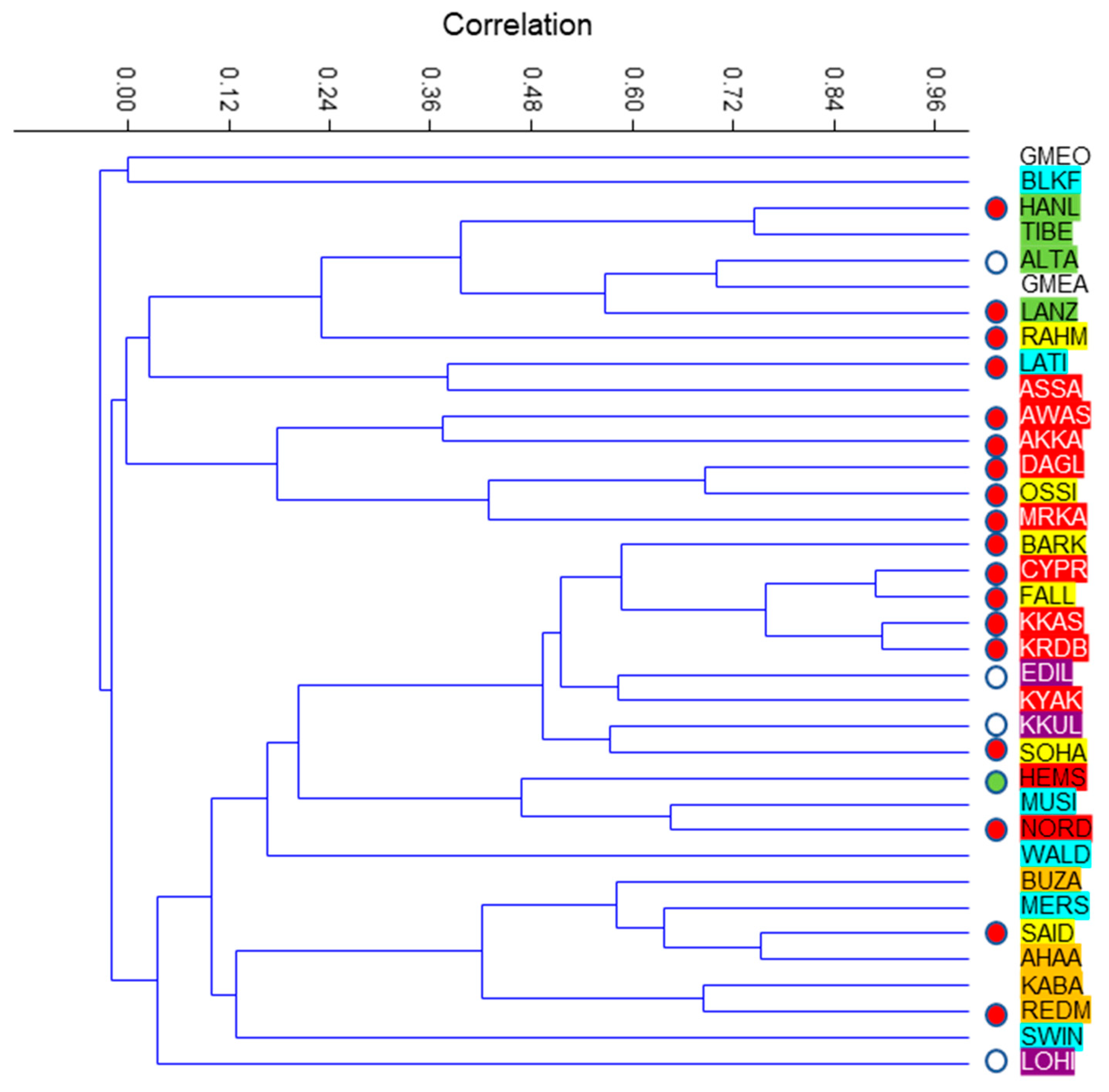
3.3. Phylogenetic Approach of the Relationships between Breeds
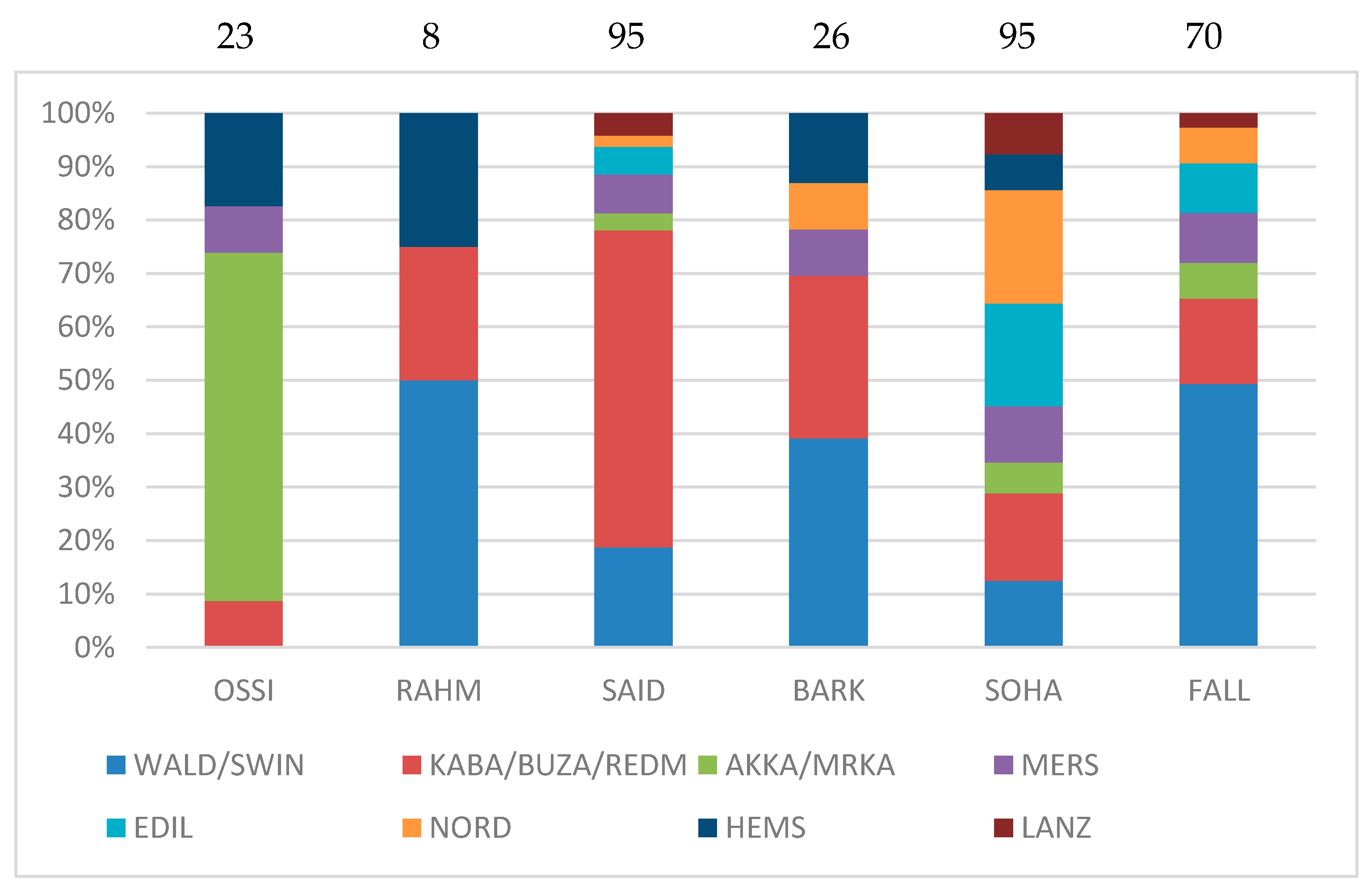
3.4. Quantification of Affinities between Egyptian and Other Breeds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peters, J.; von den Driesch, A.; Helmer, D. The upper Euphrates-Tigris basin: Cradle of agro-pastoralism? In The First Steps of Animal Domestication: New Archaeological Approaches, Proceedings of the 9th ICAZ Conference, Durham, UK, 23–28 August 2002; Vigne, J.-D., Peters, J., Helmer, D., Eds.; Oxbow Books: Oxford, UK, 2005; pp. 96–124. [Google Scholar]
- Rezaei, H.R.; Naderi, S.; Chintauan-Marquier, I.C.; Taberlet, P.; Virk, A.T.; Naghash, H.R.; Rioux, D.; Kaboli, M.; Pompanon, F. Evolution and taxonomy of the wild species of the genus Ovis (Mammalia, Artiodactyla, Bovidae). Mol. Phylogenet. Evol. 2010, 54, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Demirci, S.; Baştanlar, E.K.; Dağtaş, N.D.; Pişkin, E.; Engin, A.; Özer, F.; Yüncü, E.; Doğan, Ş.A.; Togan, İ. Mitochondrial DNA diversity of modern, ancient and wild sheep (Ovis gmelinii anatolica) from Turkey: New insights on the evolutionary history of sheep. PLoS ONE 2013, 8, e81952. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.; Hiendleder, S.; Kijas, J.W. Haplogroup relationships between domestic and wild sheep resolved using a mitogenome panel. Heredity 2011, 106, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [Green Version]
- Deniskova, T.E.; Dotsev, A.V.; Selionova, M.I.; Kunz, E.; Medugorac, I.; Reyer, H.; Wimmers, K.; Barbato, M.; Traspov, A.A.; Brem, G.; et al. Population structure and genetic diversity of 25 Russian sheep breeds based on whole-genome genotyping. Genet. Sel. Evol. 2018, 50, 29. [Google Scholar] [CrossRef] [Green Version]
- Kalds, P.; Luo, Q.; Sun, K.; Zhou, S.; Chen, Y.; Wang, X. Trends towards revealing the genetic architecture of sheep tail patterning: Promising genes and investigatory pathways. Anim. Genet. 2021, 52, 799–812. [Google Scholar] [CrossRef] [PubMed]
- Moradi, M.H.; Nejati-Javaremi, A.; Moradi-Shahrbabak, M.; Dodds, K.G.; McEwan, J.C. Genomic scan of selective sweeps in thin and fat tail sheep breeds for identifying of candidate regions associated with fat deposition. BMC Genet. 2012, 13, 10. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.H.; Peng, W.F.; Yang, J.; Zhao, Y.X.; Li, W.R.; Liu, M.J.; Ma, Y.H.; Zhao, Q.J.; Yang, G.L.; Wang, F.; et al. Mitogenomic meta-analysis identifies two phases of migration in the history of eastern Eurasian sheep. Mol. Biol. Evol. 2015, 32, 2515–2533. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.S.; Ren, X.; Yang, G.L.; Xie, X.L.; Zhao, Y.X.; Zhang, M.; Shen, Z.Q.; Ren, Y.L.; Gao, L.; Shen, M.; et al. Genome-wide association analysis identifies the genetic basis of fat deposition in the tails of sheep (Ovis aries). Anim. Genet. 2017, 48, 560–569. [Google Scholar] [CrossRef] [Green Version]
- Ahbara, A.; Bahbahani, H.; Almathen, F.; Al Abri, M.; Agoub, M.O.; Abeba, A.; Kebede, A.; Musa, H.H.; Mastrangelo, S.; Pilla, F.; et al. Genome-wide variation, candidate regions and genes associated with fat deposition and tail morphology in Ethiopian indigenous sheep. Front. Genet. 2019, 9, 699. [Google Scholar] [CrossRef]
- Marshall, F. The Origins and Spread of Domestic Animals in East Africa. In The Origins and Development of African Livestock: Archaeology, Genetics, Linguistics and Ethnography; University College London Press: London, UK, 2000; pp. 191–221. [Google Scholar]
- Gifford-Gonzalez, D.; Hanotte, O. Domesticating animals in Africa: Implications of genetic and archaeological findings. J. World Prehist. 2011, 24, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Ryder, M.L. Sheep and Man; Gerald Duckworth & Co., Ltd.: London, UK, 1983; p. 846. [Google Scholar]
- Elshazly, A.G.; Youngs, C.R. Feasibility of utilizing advanced reproductive technologies for sheep breeding in Egypt. Part 1. Genetic and nutritional resources. Egypt. J. Sheep Goats Sci. 2019, 14, 39–52. [Google Scholar]
- De Pauw, E.; Rischkowsky, B.; Abou-Naga, A.; Ansari-Renani, H.R.; Boujenane, I.; Gursoy, O. Use of GIS tools for the integration of production environment descriptors of animal genetic resources. In Final Report of Project Practical Application of Production Environment Descriptors for Animal Genetic Resources–Country Case Studies for Sheep and Goat Breeds (Letter of Agreement of FAO with ICARDA PR 43410); FAO: Rome, Italy, 2011; pp. 16–25. [Google Scholar]
- Galal, S.; Abdel-Rasoul, F.; Anous, M.R.; Shaat, I.M. On-station characterization of small ruminant breeds in Egypt. In Characterization of Small Ruminant Breeds in West Asia and North Africa; Iñiguez, L., Ed.; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 2005; Volume 2, pp. 141–193. [Google Scholar]
- Mohapatra, A.; Shinde, A.K. Fat-tailed sheep-an important sheep genetic resource for meat production in tropical countries: An overview. Indian J. Small Rumin. 2018, 24, 1. [Google Scholar] [CrossRef]
- Elshennawy, M. Sheep development program in Egypt. Strategies for sheep and goat breeding. Strateg. Sheep Goat Breed. 1995, 11, 27–32. [Google Scholar]
- Porter, V.; Alderson, L.; Hall, S.J.; Sponenberg, D.P. Mason’s World Encyclopedia of Livestock Breeds and Breeding; CABI: Singapore, 2016; Volume 2, p. 200. [Google Scholar]
- Guirgis, R.A. Egyptian Sheep Resources. Anim. Genet. Resour. Inf. 1994, 13, 47–58. [Google Scholar] [CrossRef]
- Othman, O.E.; Balabel, E.A.; Abdel-Samad, M.F. Mitochondrial DNA diversity in five Egyptian sheep breeds. Glob. Vet. 2014, 12, 369–375. [Google Scholar]
- Othman, O.E.; Pariset, L.; Balabel, E.A.; Marioti, M. Genetic characterization of Egyptian and Italian sheep breeds using mitochondrial DNA. J. Gen. Eng. Biotechnol. 2015, 13, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Othman, O.E.; Germot, A.; Khodary, M.G.; Petit, D.; Maftah, A. Cytochrome b Diversity and Phylogeny of Six Egyptian Sheep Breeds. Ann. Res. Rev. Biol. 2018, 22, 1–11. [Google Scholar] [CrossRef]
- Othman, O.E.; Payet-Duprat, N.; Harkat, S.; Laoun, A.; Maftah, A.; Lafri, M.; Da Silva, A. Sheep diversity of five Egyptian breeds: Genetic proximity revealed between desert breeds: Local sheep breeds diversity in Egypt. Small Rumin. Res. 2016, 144, 346–352. [Google Scholar] [CrossRef]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep mitochondrial DNA variation in European, Caucasian, and Central Asian areas. Mol. Biol. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3. 0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Yano, T. Dating the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Yang, Z. Maximum likelihood phylogenetic estimation from DNA sequences with variable rates over sites: Approximate methods. J. Mol. Evol. 1994, 39, 306–314. [Google Scholar] [CrossRef] [Green Version]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Kandoussi, A.; Boujenane, I.; Auger, S.; Serranito, B.; Germot, A.; Piro, M.; Maftah, A.; Petit, D. The origin of sheep settlement in Western Mediterranean. Sci. Rep. 2020, 10, 10225. [Google Scholar] [CrossRef]
- Serranito, B. Newick Extra v. 2. 2022. Available online: https://github.com/bserranito/NEWICK_extra (accessed on 4 August 2022).
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: PAleontological STatistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Uzun, M.; Gutiérrez-Gil, B.; Arranz, J.J.; Primitivo, F.S.; Saatci, M.; Kaya, M.; Bayón, Y. Genetic relationships among Turkish sheep. Gen. Sel. Evol. 2006, 38, 1–12. [Google Scholar] [CrossRef]
- Kandoussi, A.; Boujenane, I.; Piro, M.; Petit, D. Genetic diversity and population structure of Moroccan Beni Ahsen: Is this endangered ovine breed one of the ancestors of Merino? Ruminants 2022, 2, 201–211. [Google Scholar] [CrossRef]
- Mahfouz, E.R.; Othman, O.E.; El Nahas, S.M.; El Barody, M.A. Genetic variation between some Egyptian sheep breeds using RAPD-PCR. Res. J. Cell Mol. Biol. 2008, 2, 46–52. [Google Scholar]
- Gáspárdy, A.; Berger, B.; Zabavnik-Piano, J.; Kovács, E.; Annus, K.; Zenke, P.; Sáfár, L.; Maróti-Agóts, Á. Comparison of mtDNA control region among descendant breeds of the extinct Zaupel sheep revealed haplogroup C and D in Central Europe. Vet. Med. Sci. 2021, 7, 2330–2338. [Google Scholar] [CrossRef]
- Mason, I.L. The Sheep Breeds of the Mediterranean; FAO and Commonwealth Agricultural Bureaux: Farnham Royal, UK, 1967. [Google Scholar]
- Wezyk, S.; Wiener, G. Programmes for preservation of livestock breeds in Eastern Europe. FAO Anim. Prod. Health Pap. 1990, 80, 132–146. [Google Scholar]
- Ryder, M.L. A survey of European primitive breeds of sheep. Ann. Génét. Sél. Anim. 1981, 13, 381–418. [Google Scholar] [CrossRef]
- Ciani, E.; Lasagna, E.; D’Andrea, M.; Alloggio, I.; Marroni, F.; Ceccobelli, S.; Delgado Bermejo, J.V.; Sarti, F.M.; Kijas, J.; Lenstra, J.A.; et al. International Sheep Genomics Consortium. Merino and Merino-derived sheep breeds: A genome-wide intercontinental study. Genet. Sel. Evol. 2015, 47, 64. [Google Scholar] [CrossRef] [Green Version]
- Rushdi, H.E. Microsatellite-based genetic diversity among Egyptian sheep breeds. J. Anim. Poult. Prod. 2018, 9, 219–226. [Google Scholar] [CrossRef] [Green Version]
- Al-Atiyat, R.M.; Aljumaah, R.S. Genetic distances and phylogenetic trees of different Awassi sheep populations based on DNA sequencing. Gen. Mol. Res. 2014, 13, 6557–6568. [Google Scholar] [CrossRef]
- Saleh, A.A.; Sobhy, M.A.; Sharaby, M.A. A note on the potentials of Random Amplified Polymorphic DNA (RAPD) Technique to depict genetic variability among Egyptian and exotic Sheep Breeds raised locally. Alex. J. Agric. Res. 2017, 62, 93–97. [Google Scholar]
- Abied, A.; Ahbara, A.M.; Berihulay, H.; Xu, L.; Islam, R.; El-Hag, F.M.; Rekik, M.; Haile, A.; Han, J.L.; Ma, Y.; et al. Genome divergence and dynamics in the thin-tailed desert sheep from Sudan. Front. Genet. 2021, 12, 1296. [Google Scholar] [CrossRef]
- Mwacharo, J.M.; Kim, E.S.; Elbeltagy, A.R.; Aboul-Naga, A.M.; Rischkowsky, B.A.; Rothschild, M.F. Genomic footprints of dryland stress adaptation in Egyptian fat-tail sheep and their divergence from East African and western Asia cohorts. Sci. Rep. 2017, 7, 17647. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
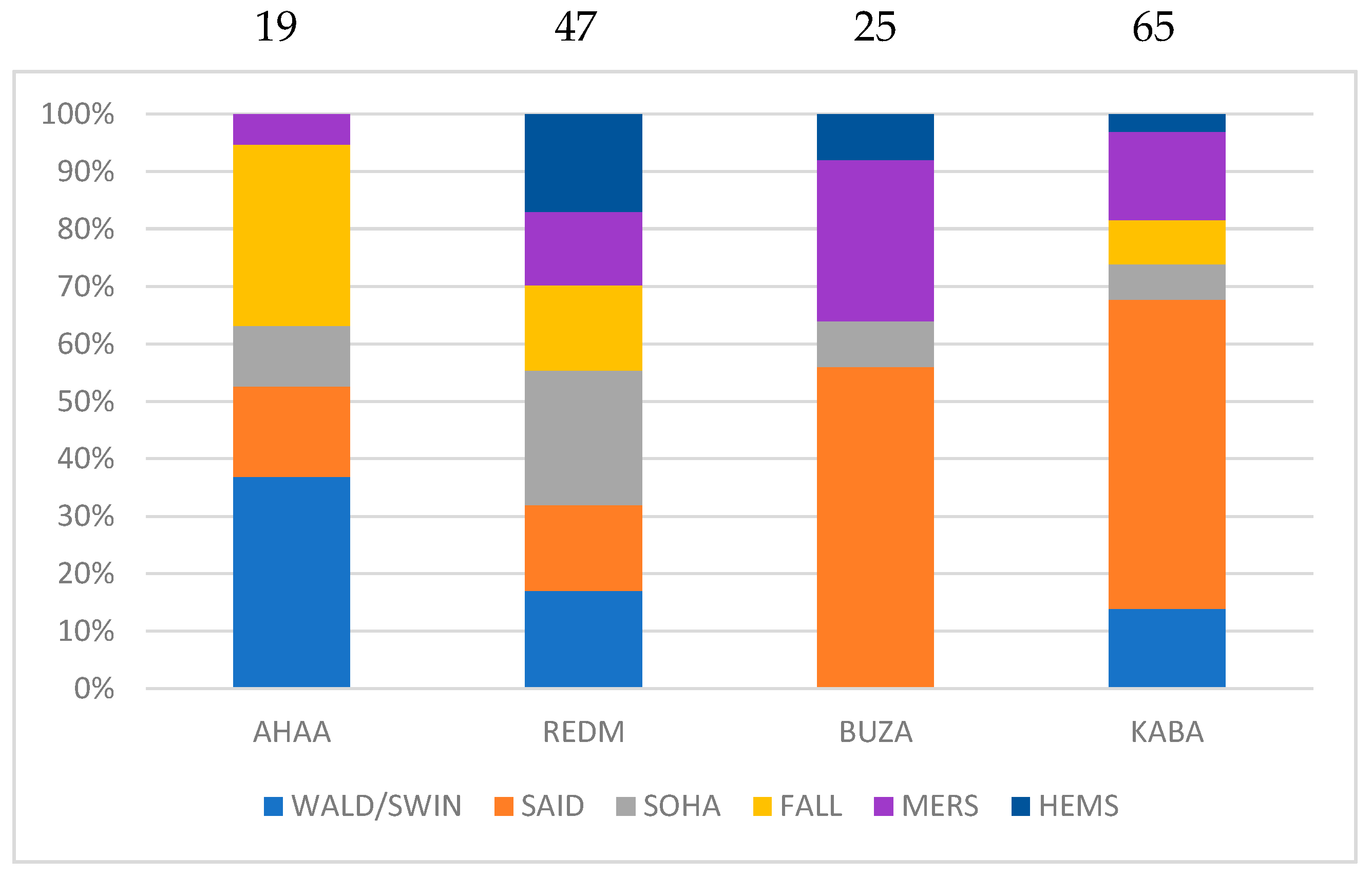{kind=link}
| Egypt | East Tropical Africa | Europe | Western Asia | Central Asia | Eastern Asia | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fat-tailed breeds | Barki BARK (17) | Fallahi (Fellahi) FALL (12) | Red Maasai REDM (20) | Laticauda LATI (5) | Akkaraman AKKA (6) | Awassi AWAS (11) | Cyprus fat tailed CYPR (8) | Edilbai (Edilbay) EDIL (11) | Altay ALTA (18) | |
| Ossimi OSSI (12) | Rahmani RAHM (11) | Daglic DAGL (19) | Hemsin HEMS (16) | Karadi KRDB (2) | Karakul KKUL (11) | Han large tailed HANL (18) | ||||
| Saidi SAID (11) | Sohagi SOHA (17) | Karakas KKAS (4) | Morkaraman MRKA (21) | Norduz NORD (16) | Lanzhou large tailed LANZ (18) | |||||
| Tuj TUJ0 (1) | ||||||||||
| Thin-tailed breeds | Ahaamda (Al Ahamda) AHAA (16) | Latxa Black face BLKF (19) | Merino MERS (20) | Assaf ASSA (4) | Karayaka KYAK (14) | Ovis gmelini anatolica GMEA (4) | Lohi LOHI (5) | Tibetan TIBE (17) | ||
| Buzaei (Buzee) BUZA (20) | Ovis aries musimom MUSI (3) | Swiniarka SWIN (19) | Ovis gmelini ophion GMEO (1) | |||||||
| Kabashi KABA (20) | Waldschaf (Waldshaf) WALD (20) | |||||||||
| Hapl A | Hapl B | Hapl C | Hapl D | Hapl E | |
|---|---|---|---|---|---|
| East Tropical Africa | 7 | 69 | 0 | 0 | 0 |
| Egypt | 11 | 63 | 5 | 0 | 1 |
| Europe | 4 | 82 | 0 | 0 | 0 |
| Western Asia | 37 | 39 | 38 | 4 | 8 |
| Central Asia | 10 | 13 | 4 | 0 | 0 |
| Eastern Asia | 38 | 29 | 5 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germot, A.; Khodary, M.G.; Othman, O.E.-M.; Petit, D. Shedding Light on the Origin of Egyptian Sheep Breeds by Evolutionary Comparison of Mitochondrial D-Loop. Animals 2022, 12, 2738. https://doi.org/10.3390/ani12202738
Germot A, Khodary MG, Othman OE-M, Petit D. Shedding Light on the Origin of Egyptian Sheep Breeds by Evolutionary Comparison of Mitochondrial D-Loop. Animals. 2022; 12(20):2738. https://doi.org/10.3390/ani12202738
Chicago/Turabian StyleGermot, Agnès, Muhammad Gamal Khodary, Othman El-Mahdy Othman, and Daniel Petit. 2022. "Shedding Light on the Origin of Egyptian Sheep Breeds by Evolutionary Comparison of Mitochondrial D-Loop" Animals 12, no. 20: 2738. https://doi.org/10.3390/ani12202738