
Isolation and Characterization of Mannanase-Producing Bacteria for Potential Synbiotic Application in Shrimp Farming
 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Diets
2.1.1. Diets Supplemented with Mannan-Oligosaccharides (MOS)
2.1.2. Shrimp Feeding Trial
2.2. Shrimp Survival Assessment under Vibrio harveyi Exposure
2.3. Tissue Sample Collection and Bacterial Screening
2.4. Enzyme Production and Culture Conditions
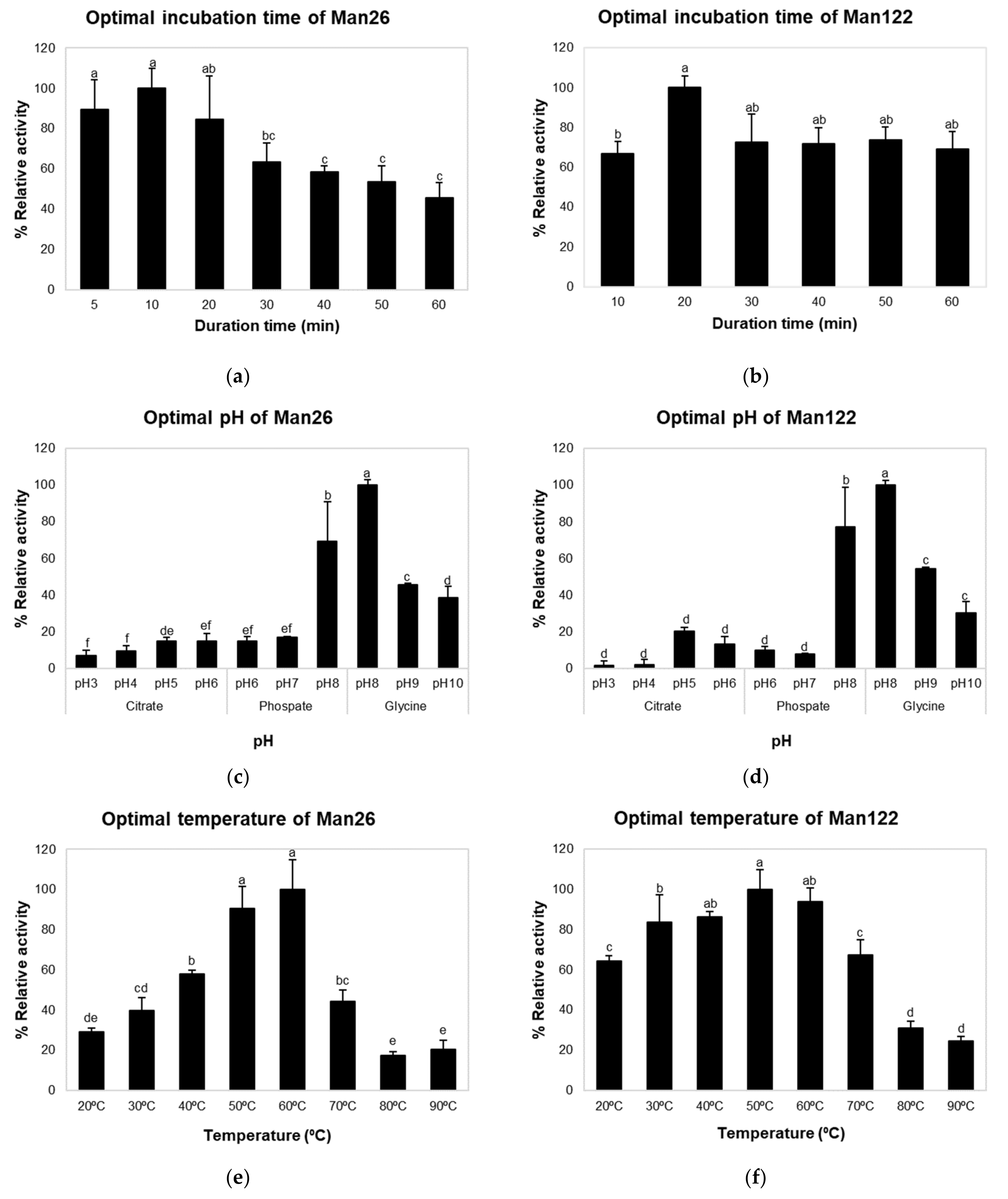
2.5. Enzyme Activity Assay and Determination of Optimal Reaction Time, pH, and Temperature
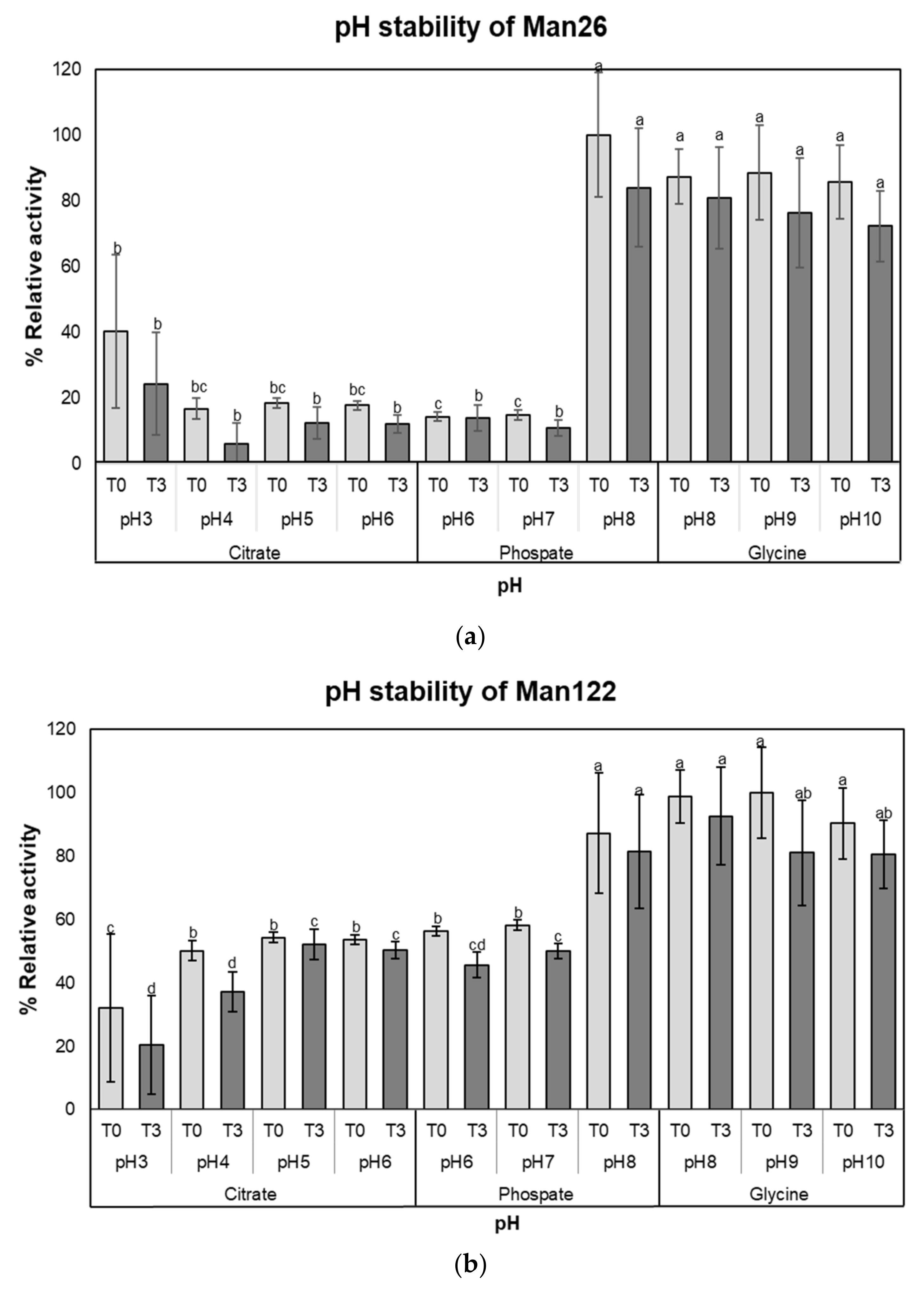
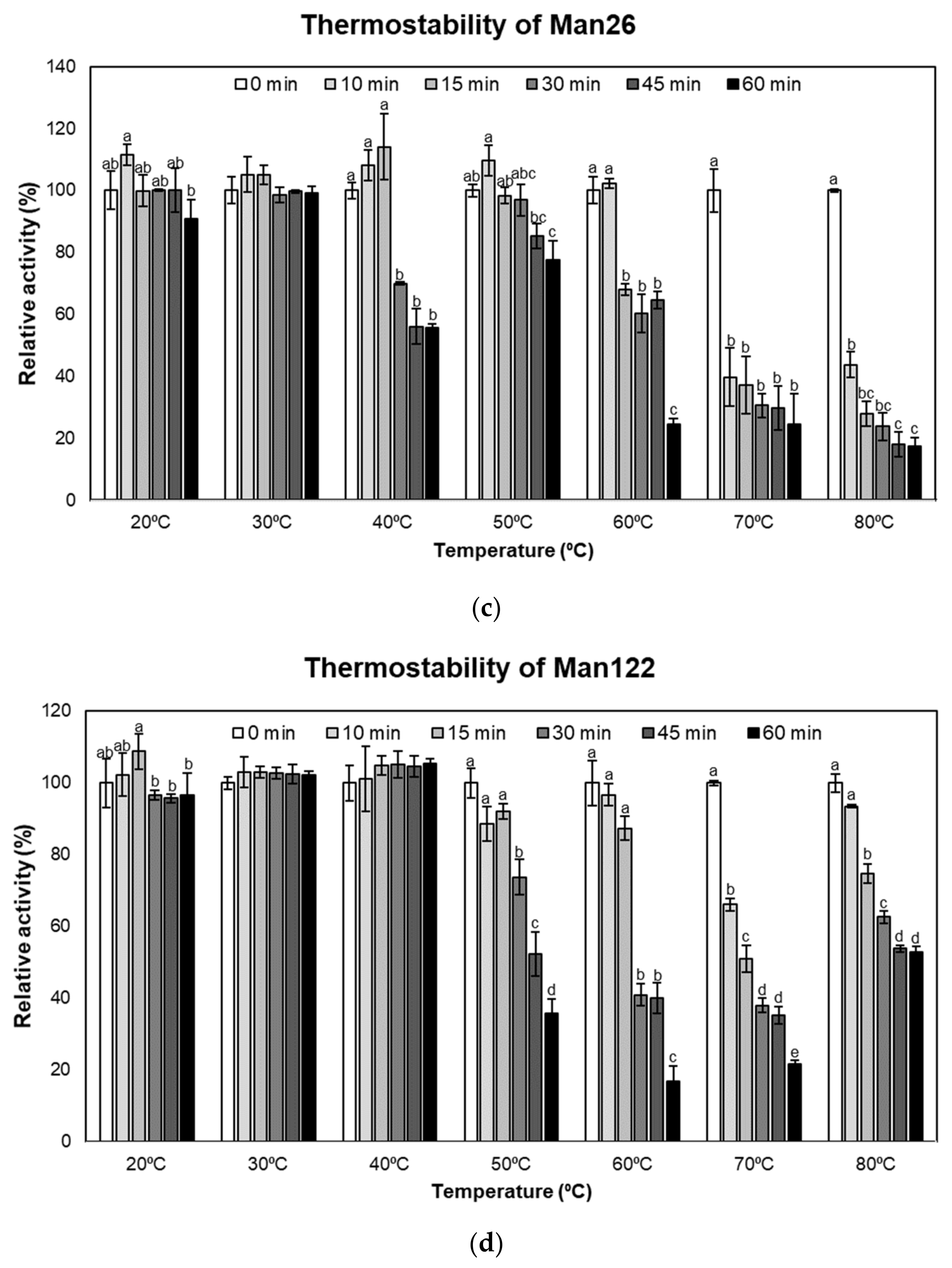
2.6. Determination of Stability and Kinetics
2.7. Determination of Substrate Specificity and Effects of Different Chemicals
2.8. Degradation of Feedstuffs
2.9. Statistical Analysis
3. Results
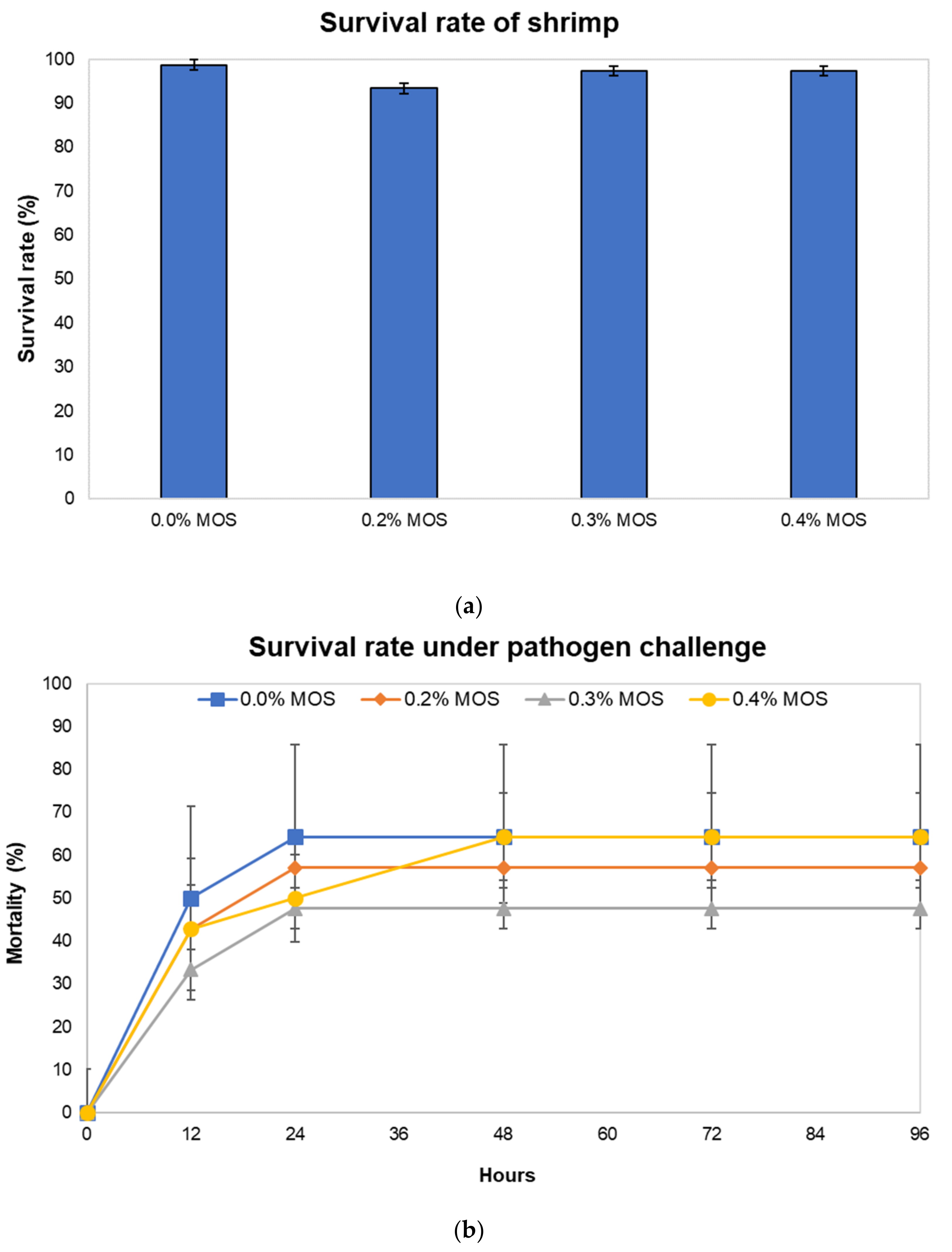
3.1. Growth Performance, Survival Rate of Shrimp under Normal Condition and under Exposure to Pathogenic Vibrio harveyi
3.2. Screening of Bacterial Isolates with Potential Function to Hydrolyze MOS
3.3. Enzymatic Characterization of Mannanase
3.4. Enzyme Activity on Selected Carbohydrate Feedstuffs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Fishery and Aquaculture Statistic 2019; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- OECD/FAO. OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021. [Google Scholar]
- FAO. FAO 2021 the State of World Fisheries and Aquaculture 2020; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Kongrueng, J.; Yingkajorn, M.; Bunpa, S.; Sermwittayawong, N.; Singkhamanan, K.; Vuddhakul, V. Characterization of Vibrio parahaemolyticus causing acute hepatopancreatic necrosis disease in southern Thailand. J. Fish Dis. 2015, 38, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Asche, F.; Anderson, J.L.; Botta, R.; Kumar, G.; Abrahamsen, E.B.; Nguyen, L.T.; Valderrama, D. The economics of shrimp disease. J. Invertebr. Pathol. 2021, 186, 107397. [Google Scholar] [CrossRef] [PubMed]
- Dashtiannasab, A. Chapter 32—White Spot Syndrome Virus. In Emerging and Reemerging Viral Pathogens; Ennaji, M.M., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 717–728. [Google Scholar]
- Amarakoon, G.; Wijegoonawardane, P. A comparative analysis of Yellow Head Virus (YHD) diagnostic methods adopted in Sri Lanka to investigate the accuracy and specificity of the virus. J. World Aquac. Soc. 2017, 66, 181–192. [Google Scholar]
- Dawood, M.; Koshio, S.; Esteban, M. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2017, 10, 950–974. [Google Scholar] [CrossRef]
- Chen, M.; Chen, X.-Q.; Tian, L.-X.; Liu, Y.-J.; Niu, J. Beneficial impacts on growth, intestinal health, immune responses and ammonia resistance of pacific white shrimp (Litopenaeus vannamei) fed dietary synbiotic (mannan oligosaccharide and Bacillus licheniformis). Aquac. Rep. 2020, 17, 100408. [Google Scholar] [CrossRef]
- Encarnação, P. Functional feed additives in aquaculture feeds. In Aquafeed Formulation; Nates, S.F., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 217–237. [Google Scholar]
- Flachowsky, G. Chapter 5—Influence of Feed from Genetically Modified Plants on the Composition and Quality of Foods of Animal Origin. In Genetically Engineered Foods; Holban, A.M., Grumezescu, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 109–141. [Google Scholar]
- Li, E.; Xu, C.; Wang, X.; Wang, S.; Zhao, Q.; Zhang, M.; Qin, J.G.; Chen, L. Gut Microbiota and its Modulation for Healthy Farming of Pacific White Shrimp Litopenaeus vannamei. Rev. Fish. Sci. Aquac. 2018, 26, 381–399. [Google Scholar] [CrossRef]
- Holt, C.C.; Bass, D.; Stentiford, G.D.; van der Giezen, M. Understanding the role of the shrimp gut microbiome in health and disease. J. Invertebr. Pathol. 2021, 186, 107387. [Google Scholar] [CrossRef]
- Dai, W.; Sheng, Z.; Chen, J.; Xiong, J. Shrimp disease progression increases the gut bacterial network complexity and abundances of keystone taxa. Aquaculture 2020, 517, 734802. [Google Scholar] [CrossRef]
- Clarke, G.; Stilling, R.M.; Kennedy, P.J.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Minireview: Gut Microbiota: The Neglected Endocrine Organ. J. Mol. Endocrinol. 2014, 28, 1221–1238. [Google Scholar] [CrossRef]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Ochoa-Romo, J.P.; Cornejo-Granados, F.; Lopez-Zavala, A.A.; Viana, M.T.; Sánchez, F.; Gallardo-Becerra, L.; Luque-Villegas, M.; Valdez-López, Y.; Sotelo-Mundo, R.R.; Cota-Huízar, A.; et al. Agavin induces beneficial microbes in the shrimp microbiota under farming conditions. Sci. Rep. 2022, 12, 6392. [Google Scholar] [CrossRef]
- Ringø, E.; Olsen, R.E.; Gifstad, T.Ø.; Dalmo, R.A.; Amlund, H.; Hemre, G.-I.; Bakke, A.M. Prebiotics in aquaculture: A review. Aquac. Nutr. 2010, 16, 117–136. [Google Scholar] [CrossRef]
- Zhou, Z.; Ding, Z.; Huiyuan, L. Effects of Dietary Short-chain Fructooligosaccharides on Intestinal Microflora, Survival, and Growth Performance of Juvenile White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2007, 38, 296–301. [Google Scholar] [CrossRef]
- Mahious, A.S.; Gatesoupe, F.J.; Hervi, M.; Metailler, R.; Ollevier, F. Effect of dietary inulin and oligosaccharides as prebiotics for weaning turbot, Psetta maxima (Linnaeus, C. 1758). Aquac. Int. 2005, 14, 219. [Google Scholar] [CrossRef]
- Guzmán-Villanueva, L.T.; Tovar-Ramírez, D.; Gisbert, E.; Cordero, H.; Guardiola, F.A.; Cuesta, A.; Meseguer, J.; Ascencio-Valle, F.; Esteban, M.A. Dietary administration of β-1,3/1,6-glucan and probiotic strain Shewanella putrefaciens, single or combined, on gilthead seabream growth, immune responses and gene expression. Fish Shellfish Immunol. 2014, 39, 34–41. [Google Scholar] [CrossRef]
- Nopvichai, C.; Charoenwongpaiboon, T.; Luengluepunya, N.; Ito, K.; Muanprasat, C.; Pichyangkura, R. Production and purification of mannan oligosaccharide with epithelial tight junction enhancing activity. PeerJ 2019, 7, e7206. [Google Scholar] [CrossRef]
- Faustino, M.; Durão, J.; Pereira, C.F.; Pintado, M.E.; Carvalho, A.P. Mannans and mannan oligosaccharides (MOS) from Saccharomyces cerevisiae—A sustainable source of functional ingredients. Carbohydr. Polym. 2021, 272, 118467. [Google Scholar] [CrossRef]
- Zhang, P.; Yang, F.; Hu, J.; Han, D.; Liu, H.; Jin, J.; Yang, Y.; Yi, J.; Zhu, X.; Xie, S. Optimal form of yeast cell wall promotes growth, immunity and disease resistance in gibel carp (Carassius auratus gibelio). Aquac. Rep. 2020, 18, 100465. [Google Scholar] [CrossRef]
- Refstie, S.; Baeverfjord, G.; Seim, R.R.; Elvebø, O. Effects of dietary yeast cell wall β-glucans and MOS on performance, gut health, and salmon lice resistance in Atlantic salmon (Salmo salar) fed sunflower and soybean meal. Aquaculture 2010, 305, 109–116. [Google Scholar] [CrossRef]
- Abu-Elala, N.M.; Younis, N.A.; AbuBakr, H.O.; Ragaa, N.M.; Borges, L.L.; Bonato, M.A. Efficacy of dietary yeast cell wall supplementation on the nutrition and immune response of Nile tilapia. Egypt. J. Aquat. Res. 2018, 44, 333–341. [Google Scholar] [CrossRef]
- Gainza, O.; Romero, J. Effect of mannan oligosaccharides on the microbiota and productivity parameters of Litopenaeus vannamei shrimp under intensive cultivation in Ecuador. Sci. Rep. 2020, 10, 2719. [Google Scholar] [CrossRef] [Green Version]
- Rungrassamee, W.; Kingcha, Y.; Srimarut, Y.; Maibunkaew, S.; Karoonuthaisiri, N.; Visessanguan, W. Mannooligosaccharides from copra meal improves survival of the Pacific white shrimp (Litopenaeus vannamei) after exposure to Vibrio harveyi. Aquaculture 2014, 434, 403–410. [Google Scholar] [CrossRef]
- Genc, M.A.; Aktas, M.; Genc, E.; Yilmaz, E. Effects of dietary mannan oligosaccharide on growth, body composition and hepatopancreas histology of Penaeus semisulcatus (de Haan 1844). Aquac. Nutr. 2007, 13, 156–161. [Google Scholar] [CrossRef]
- Chai, P.C.; Song, X.L.; Chen, G.F.; Xu, H.; Huang, J. Dietary supplementation of probiotic Bacillus PC465 isolated from the gut of Fenneropenaeus chinensis improves the health status and resistance of Litopenaeus vannamei against white spot syndrome virus. Fish Shellfish Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef]
- Wu, C.-C.; Chang, Y.-P.; Wang, J.-J.; Liu, C.-H.; Wong, S.-L.; Jiang, C.-M.; Hsieh, S.-L. Dietary administration of Gynura bicolor (Roxb. Willd.) DC water extract enhances immune response and survival rate against Vibrio alginolyticus and white spot syndrome virus in white shrimp Litopeneaus vannamei. Fish Shellfish Immunol. 2015, 42, 25–33. [Google Scholar] [CrossRef]
- Vieira, F.D.; Buglione Neto, C.C.; Mourino, J.L.P.; Jatobá, A.; Ramirez, C.; Martins, M.L.; Barracco, M.A.A.M.; Vinatea, L.A. Time-related action of Lactobacillus plantarum in the bacterial microbiota of shrimp digestive tract and its action as immunostimulant. Pesqui. Agropecu. Bras. 2008, 43, 763–769. [Google Scholar] [CrossRef]
- Cruz, P.M.; Ibáñez, A.L.; Hermosillo, O.A.M.; Saad, H.C.R. Use of probiotics in aquaculture. ISRN Microbiol. 2012, 2012, 916845. [Google Scholar] [CrossRef]
- Sahu, M.K.; Swarnakumar, N.S.; Sivakumar, K.; Thangaradjou, T.; Kannan, L. Probiotics in aquaculture: Importance and future perspectives. Indian J. Microbiol. 2008, 48, 299–308. [Google Scholar] [CrossRef]
- Hai, N.V. The use of probiotics in aquaculture. J. Appl. Microbiol. 2015, 119, 917–935. [Google Scholar] [CrossRef]
- Butt, U.D.; Lin, N.; Akhter, N.; Siddiqui, T.; Li, S.; Wu, B. Overview of the latest developments in the role of probiotics, prebiotics and synbiotics in shrimp aquaculture. Fish Shellfish Immunol. 2021, 114, 263–281. [Google Scholar] [CrossRef]
- Sritunyalucksana, K.; Gangnonngiw, W.; Archakunakorn, S.; Fegan, D.; Flegel, T.W. Bacterial clearance rate and a new differential hemocyte staining method to assess immunostimulant activity in shrimp. Dis. Aquat. Org. 2005, 63, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Ghazala, I.; Haddar, A.; Romdhane, M.; Ellouz-Chaanouni, S. Screening and Molecular Identification of New Microbial Strains for Production of Enzymes of Biotechnological Interest. Braz. Arch. Biol. Technol. 2016, 59, e16150152. [Google Scholar] [CrossRef]
- Gessesse, A.; Gashe, B.A. Production of alkaline xylanase by an alkaliphilic Bacillus sp. isolated from an alkalinesoda lake. J. Appl. Microbiol. 1997, 83, 402–406. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sathitkowitchai, W.; Wongsurawat, T.; Jenjaroenpun, P.; Angthong, P.; Nuanpirom, J.; Sathapondecha, P.; Rungrassamee, W. Complete Genome Sequences of Mannanase-Producing Bacillus and Niallia strains isolated from the intestine of the black tiger shrimp (Penaeus monodon). Microbiol. Resour. Announc. 2022, 11, e00112-22. [Google Scholar] [CrossRef]
- Huynh, T.-G.; Shiu, Y.-L.; Nguyen, T.-P.; Truong, Q.-P.; Chen, J.-C.; Liu, C.-H. Current applications, selection, and possible mechanisms of actions of synbiotics in improving the growth and health status in aquaculture: A review. Fish Shellfish Immunol. 2017, 64, 367–382. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Dai, X.; Li, J.; Ding, F. Effects of dietary inulin and mannan oligosaccharide on immune related genes expression and disease resistance of Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2018, 76, 78–92. [Google Scholar] [CrossRef]
- Zorriehzahra, M.J.; Delshad, S.T.; Adel, M.; Tiwari, R.; Karthik, K.; Dhama, K.; Lazado, C.C. Probiotics as beneficial microbes in aquaculture: An update on their multiple modes of action: A review. Vet. Q. 2016, 36, 228–241. [Google Scholar] [CrossRef]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Nejat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [Green Version]
- Akhter, N.; Wu, B.; Memon, A.M.; Mohsin, M. Probiotics and prebiotics associated with aquaculture: A review. Fish Shellfish Immunol. 2015, 45, 733–741. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef]
- Zheng, C.c.; Wu, J.w.; Jin, Z.h.; Ye, Z.f.; Yang, S.; Sun, Y.q.; Fei, H. Exogenous enzymes as functional additives in finfish aquaculture. Aquac. Nutr. 2020, 26, 213–224. [Google Scholar] [CrossRef]
- Castillo, S.; Gatlin, D.M. Dietary supplementation of exogenous carbohydrase enzymes in fish nutrition: A review. Aquaculture 2015, 435, 286–292. [Google Scholar] [CrossRef]
- Adeola, O.; Cowieson, A.J. BOARD-INVITED REVIEW: Opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. J. Anim. Sci. 2011, 89, 3189–3218. [Google Scholar] [CrossRef]
- Nathan, F.; Elangovan, P.; Kannan, B.; Kalidoss, M. An evidential review on potential benefits of enzymes in aqua feed industry. Int. J. Curr. Microbiol. 2018, 7, 2053–2074. [Google Scholar] [CrossRef]
- Ge, J.P.; Du, R.P.; Zhao, D.; Song, G.; Jin, M.; Ping, W.X. Bio-chemical characterization of a β-mannanase from Bacillus licheniformis HDYM-04 isolated from flax water-retting liquid and its decolorization ability of dyes. RSC Adv. 2016, 6, 23612–23621. [Google Scholar] [CrossRef]
- Kim, S.; Lee, M.-H.; Lee, E.-S.; Nam, Y.-D.; Seo, D.-H. Characterization of mannanase from Bacillus sp., a novel Codium fragile cell wall-degrading bacterium. Food Sci. Biotechnol. 2017, 27, 115–122. [Google Scholar] [CrossRef]
- Mamo, G. Alkaline Active Hemicellulases. Adv. Biochem. Eng. Biotechnol. 2020, 172, 245–291. [Google Scholar] [CrossRef]
- Dawood, A.; Ma, K. Applications of microbial β-Mannanases. Front. Bioeng. Biotechnol. 2020, 8, 598630. [Google Scholar] [CrossRef]
- Vera, C.; Illanes, A.; Guerrero, C. Enzymatic production of prebiotic oligosaccharides. Curr. Opin. Food Sci. 2021, 37, 160–170. [Google Scholar] [CrossRef]
- Sathitkowitchai, W.; Nitisinprasert, S.; Keawsompong, S. Improving palm kernel cake nutrition using enzymatic hydrolysis optimized by Taguchi method. 3 Biotech 2018, 8, 407. [Google Scholar] [CrossRef]
- Rungruangsaphakun, J.; Nakphaichit, M.; Keawsompong, S. Nutritional improvement of copra meal for swine feed. Biocatal. Agric. Biotechnol. 2022, 39, 102273. [Google Scholar] [CrossRef]
- Balasubramaniam, K. Polysaccharides of the kernel of maturing and matured coconuts. J. Food Sci. 1976, 41, 1370–1373. [Google Scholar] [CrossRef]
- Düsterhöft, E.-M.; Posthumus, M.A.; Voragen, A.G.J. Non-starch polysaccharides from sunflower (Helianthus annuus) meal and palm-kernel (Elaeis guineensis) meal—investigation of the structure of major polysaccharides. J. Sci. Food Agric. 1992, 59, 151–160. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Man26 | Man122 |
|---|---|---|
| Cell morphology | Rod shape, Gram stain positive | Short rod shape, Gram stain positive |
| Colony morphology | ||
| Colony color | White, shiny | White, dull |
| Surface | - | Mucoid |
| Margin | Regular | Irregular |
| Elevation | Swell | Flat |
| Catalase | ||
| 24 h | Positive | Positive |
| 48 h | Positive | Positive |
| Source | Optimal pH | Optimal Temperature (oC) | Vmax (U·mg−1) | Km (mg·mL−1) |
|---|---|---|---|---|
| Man26 | 8.0 | 60 | 5.52 ± 0.70 | 18.92 ± 4.36 |
| Man122 | 8.0 | 50 | 41.15 ± 12.33 | 34.53 ± 14.26 |
| Substrates | Relative Activity (%) | p Value (t-test) | |
|---|---|---|---|
| Man26 | Man122 | ||
| Locust bean gum (LBG) | 100 ± 5.51 a | 100 ± 3.42 a | - |
| Guar gum | 32.97 ± 0.37 b, x | 19.13 ± 2.53 cd, y | 0.003 |
| Carboxymethyl cellulose (CMC) | 2.52 ± 1.45 e, x | 23.16 ± 1.19 c, y | 0.002 |
| Avicel | 18.31 ± 3.47 cd, x | 4.93 ± 3.66 f, y | 0.005 |
| Xylan (larchwood) | 1.95 ± 0.88 e, x | 15.78 ± 1.46 de, y | 0.008 |
| Xylan (birchwood) | 14.81 ± 1.57 d, x | 12.82 ± 1.53 e, x | 0.354 |
| Xylan (oat spelts) | 1.54 ± 1.23 e, x | 8.09 ± 0.6 f, y | 0.004 |
| Potato starch | 22.03 ± 1.95 c, x | 33.24 ± 1.74 b, x | 0.074 |
| Chemicals | Relative activity (%) | p Value (t-test) | |
|---|---|---|---|
| Man26 | Man122 | ||
| Control | 100 ab | 100a | - |
| 10 mM CaCl2 | 67.36 ± 0.15 cd, x | 71.31 ± 1.01 d, y | 0.017 |
| 10 mM KCl | 97.51 ± 0.18 ab, x | 91.34 ± 0.25 b, x | 0.219 |
| 10 mM MgCl2 | 105.38 ± 0.71 a, x | 67.81 ± 0.65 d, y | 0.016 |
| 10 mM MgSO4 | 93.37 ± 0.32 e, x | 60.35 ± 0.8 e, y | 0.022 |
| 10 mM NaCl | 93.52 ± 0.49 b, x | 85.75 ± 0.34 c, x | 0.277 |
| 10 mM Na2SO3 | 63.41 ± 0.38 d, x | 26.05 ± 0.52 f, y | 0.014 |
| 10 mM NH4Cl | 74.09 ± 0.15 c, x | 61.47 ± 0.61 e, y | 0.011 |
| 50 mM EDTA | 43.54 ± 0.2 e, x | 4.53 ± 0.13 g, y | 0.014 |
| 1% SDS | 24.55 ± 0.5 f, x | 1.38 ± 0.34 g, y | 0.015 |
| 5% SDS | 23.66 ± 0.12 f, x | 0.36 ± 0.03 g, y | 0.002 |
| 10% SDS | 20.56 ± 0.17 f, x | 0.31 ± 0.05 g, y | 0.003 |
| % | SubstratesΨ | Crude Mannanase of Man26 | |||||
|---|---|---|---|---|---|---|---|
| − | + | − | + | − | + | ||
| Reducing sugar (Mg) Ψ | Total Sugar (mg) Ψ | Protein Release (mg) Ψ | |||||
| 1% | Palm kernel cake (PKC) | 2.42 ± 0.22 | 20.15 ± 0.48 * | 14.05 ± 1.24 | 28.53 ± 14.57 | 5.33 ± 0.12 | 8.55 ± 0.26 * |
| Copra meal (CPM) | 0.76 ± 0.18 | 28.71 ± 0.60 * | 8.83 ± 0.84 | 33.21 ± 21.17 * | 5.70 ± 0.61 | 8.93 ± 0.20 * | |
| 5% | Palm kernel cake (PKC) | 13.84 ± 0.55 | 52.58 ± 1.62 * | 61.74 ± 1.48 | 79.19 ± 24.94 | 7.13 ± 0.28 | 9.60 ± 0.45 * |
| Copra meal (CPM) | 4.12 ± 0.31 | 88.14 ± 2.48 * | 35.98 ± 4.37 | 84.26 ± 54.21 | 5.61 ± 0.23 | 9.03 ± 0.13 * | |
| 1% | Maize | 3.69 ± 0.30 | 15.54 ± 0.36 * | 7.99 ± 1.54 | 24.29 ± 18.25 | 6.16 ± 0.24 | 9.15 ± 0.35 * |
| Broken rice | 17.53 ± 0.94 | 58.69 ± 2.30 * | 21.56 ± 0.49 | 54.26 ± 25.39 * | 4.77 ± 0.33 | 8.34 ± 0.52 * | |
| Soybean | 0.18 ± 0.07 | 8.07 ± 1.04 * | 46.85 ± 2.71 | 59.01 ± 11.49 | 9.29 ± 1.16 | 14.62 ± 0.26 * | |
| Feed | 5.09 ± 1.19 | 5.95 ± 1.44 | 24.89 ± 6.49 | 25.74 ± 5.24 | 8.14 ± 0.07 | 10.75 ± 0.26 * | |
| 5% | Maize | 61.88 ± 2.29 | 81.09 ± 1.40 * | 69.55 ± 4.25 | 89.25 ± 15.28 * | 7.41 ± 0.26 | 10.10 ± 0.10 * |
| Broken rice | 239.65 ± 7.81 | 261.29 ± 2.85 * | 245.05 ± 26.36 | 259.64 ± 70.81 | 8.69 ± 0.19 | 10.72 ± 0.34 * | |
| Soybean | 15.28 ± 4.37 | 21.81 ± 3.90 * | 164.26 ± 4.26 | 190.63 ± 26.31 | 10.70 ± 0.15 | 15.49 ± 0.27 * | |
| Commercial shrimp feed | 9.68 ± 1.01 | 11.05 ± 0.56 | 81.35 ± 1.67 | 84.20 ± 6.00 | 10.96 ± 0.30 | 13.43 ± 0.30 * | |
| % | SubstratesΨ | Crude Mannanase of Man122 | |||||
|---|---|---|---|---|---|---|---|
| − | + | − | + | − | + | ||
| Reducing Sugar (mg) Ψ | Total Sugar (mg) Ψ | Protein Release (mg) Ψ | |||||
| 1% | Palm kernel cake (PKC) | 1.58 ± 0.25 | 12.30 ± 0.59 * | 14.38 ± 0.74 | 24.17 ± 2.06 * | 5.15 ± 0.24 | 10.31 ± 0.18 * |
| Copra meal (CPM) | 0.80 ± 0.73 | 30.47 ± 0.87 * | 11.05 ± 2.52 | 49.19 ± 2.85 * | 5.79 ± 0.31 | 8.91 ± 0.05 * | |
| 5% | Palm kernel cake (PKC) | 15.29 ± 2.17 | 50.90 ± 3.50 * | 54.14 ± 2.71 | 97.00 ± 7.25 * | 7.32 ± 0.52 | 11.15 ± 0.23 * |
| Copra meal (CPM) | 5.49 ± 1.75 | 197.15 ± 5.49 * | 31.71 ± 0.79 | 394.60 ± 23.77 * | 6.08 ± 0.05 | 12.06 ± 0.44 * | |
| 1% | Maize | 4.95 ± 0.34 | 17.07 ± 2.06 * | 7.87 ± 2.43 | 36.58 ± 4.25 * | 6.04 ± 0.22 | 9.36 ± 0.22 * |
| Broken rice | 27.08 ± 1.52 | 34.99 ± 4.46 * | 31.02 ± 1.82 | 47.03 ± 3.73 * | 4.85 ± 0.06 | 8.33 ± 0.16 * | |
| Soybean | 0.23 ± 0.13 | 11.68 ± 0.50 * | 47.45 ± 2.11 | 48.26 ± 2.40 | 8.31 ± 0.16 | 14.78 ± 0.20 * | |
| Feed | 0.09 ± 0.05 | 22.56 ± 0.58 * | 18.20 ± 1.62 | 50.00 ± 3.60 * | 7.57 ± 0.26 | 11.2 ± 0.07 * | |
| 5% | Maize | 67.68 ± 1.10 | 85.68 ± 26.14 | 77.72 ± 5.88 | 120.84 ± 32.93 * | 7.25 ± 0.54 | 9.61 ± 0.07 * |
| Broken rice | 184.09 ± 11.75 | 322.85 ± 17.69 * | 205.05 ± 18.95 | 382.94 ± 15.27 * | 7.11 ± 0.44 | 11.07 ± 0.69 * | |
| Soybean | 11.62 ± 4.05 | 33.56 ± 1.83 * | 169.82 ± 12.65 | 178.56 ± 14.96 * | 9.42 ± 1.05 | 15.36 ± 0.23 * | |
| Commercial shrimp feed | 6.40 ± 2.98 | 33.46 ± 8.44 * | 78.23 ± 2.62 | 137.90 ± 13.47 * | 9.28 ± 0.43 | 13.13 ± 0.18 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sathitkowitchai, W.; Sathapondecha, P.; Angthong, P.; Srimarut, Y.; Malila, Y.; Nakkongkam, W.; Chaiyapechara, S.; Karoonuthaisiri, N.; Keawsompong, S.; Rungrassamee, W. Isolation and Characterization of Mannanase-Producing Bacteria for Potential Synbiotic Application in Shrimp Farming. Animals 2022, 12, 2583. https://doi.org/10.3390/ani12192583
Sathitkowitchai W, Sathapondecha P, Angthong P, Srimarut Y, Malila Y, Nakkongkam W, Chaiyapechara S, Karoonuthaisiri N, Keawsompong S, Rungrassamee W. Isolation and Characterization of Mannanase-Producing Bacteria for Potential Synbiotic Application in Shrimp Farming. Animals. 2022; 12(19):2583. https://doi.org/10.3390/ani12192583
Chicago/Turabian StyleSathitkowitchai, Witida, Ponsit Sathapondecha, Pacharaporn Angthong, Yanee Srimarut, Yuwares Malila, Wuttichai Nakkongkam, Sage Chaiyapechara, Nitsara Karoonuthaisiri, Suttipun Keawsompong, and Wanilada Rungrassamee. 2022. "Isolation and Characterization of Mannanase-Producing Bacteria for Potential Synbiotic Application in Shrimp Farming" Animals 12, no. 19: 2583. https://doi.org/10.3390/ani12192583