
Evaluation of Predisposing Factors of Necrotic Enteritis in Experimentally Challenged Broiler Chickens

 ,
, 
 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Samples
2.2. In Vivo Test
2.3. Inoculum Preparation and Challenge
2.4. Intestine Gross Lesion Scoring
2.5. Quantification of Aerobic and Anaerobic Bacteria
2.6. Histological Analysis
2.7. Statistical Analysis
3. Results
3.1. Experimental Infection
3.2. Evaluation of Gross Lesions of the Intestine
3.3. Quantification of Aerobic and Anaerobic Bacteria in the Liver
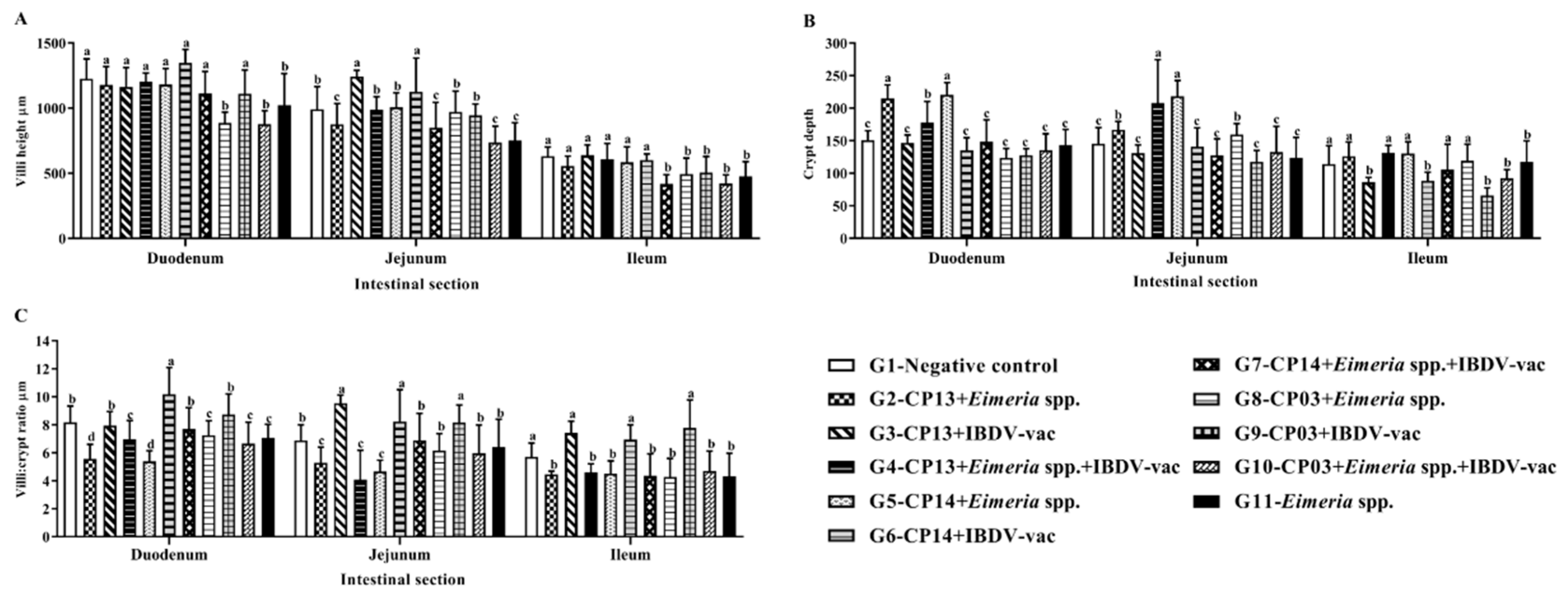
3.4. Histological Evaluation of the Intestine
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Immerseel, F.; Lyhs, U.; Pedersen, K.; Prescott, J.F. Recent Breakthroughs Have Unveiled the Many Knowledge Gaps in Clostridium perfringens-Associated Necrotic Enteritis in Chickens: The First International Conference on Necrotic Enteritis in Poultry. Avian Pathol. 2016, 45, 269–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The True Cost of Necrotic Enteritis. Poultry World. Available online: https://www.poultryworld.net/poultry/the-true-cost-of-necrotic-enteritis/ (accessed on 20 May 2022).
- Zahoor, I.; Ghayas, A.; Basheer, A. Genetics and genomics of susceptibility and immune response to necrotic enteritis in chicken: A review. Mol. Biol. Rep. 2018, 45, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.J. Necrotic enteritis predisposing factors in broiler chickens. Avian Pathol. 2016, 45, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsiouris, V. Poultry management: A useful tool for the control of necrotic enteritis in poultry. Avian Pathol. 2016, 45, 323–325. [Google Scholar] [CrossRef]
- Revitt-Mills, S.; Vidor, C.J.; Watts, T.D.; Lyras, D.; Rood, J.I.; Adams, V. Virulence Plasmids of the Pathogenic Clostridia. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Rood, J.I.; Adams, V.; Lacey, J.; Lyras, D.; McClane, B.A.; Melville, S.B.; Moore, R.J.; Popoff, M.R.; Sarker, M.R.; Songer, J.G.; et al. Expansion of the Clostridium perfringens toxin-based typing scheme. Anaerobe 2018, 53, 5–10. [Google Scholar] [CrossRef]
- Kiu, R.; Brown, J.; Bedwell, H.; Leclaire, C.; Caim, S.; Pickard, D.; Dougan, G.; Dixon, R.A.; Hall, L.J. Genomic analysis on broiler-associated Clostridium perfringens strains and exploratory caecal microbiome investigation reveals key factors linked to poultry necrotic enteritis. Anim. Microbiome 2019, 1, 12. [Google Scholar] [CrossRef] [Green Version]
- Mehdizadeh Gohari, I.; Navarro, M.A.; Li, J.; Shrestha, A.; Uzal, F.; McClane, B.A. Pathogenicity and Virulence of Clostridium perfringens. Virulence 2021, 12, 723–753. [Google Scholar] [CrossRef]
- Kaldhusdal, M.; Benestad, S.L.; Løvland, A. Epidemiologic Aspects of Necrotic Enteritis in Broiler Chickens—Disease Occurrence and Production Performance. Avian Pathol. 2016, 45, 271–274. [Google Scholar] [CrossRef]
- Crispo, M.; Stoute, S.T.; Uzal, F.A.; Bickford, A.A.; Shivaprasad, H.L. Nonenteric Lesions of Necrotic Enteritis in Commercial Chickens in California: 25 Cases (2009–2018). Avian Dis. 2020, 64, 356–364. [Google Scholar] [CrossRef]
- Broom, L.J. Necrotic Enteritis; Current Knowledge and Diet-Related Mitigation. World’s Poult. Sci. J. 2017, 73, 281–292. [Google Scholar] [CrossRef]
- Lu, M.; Li, R.W.; Zhao, H.; Yan, X.; Lillehoj, H.S.; Sun, Z.; Oh, S.; Wang, Y.; Li, C. Effects of Eimeria Maxima and Clostridium perfringens Infections on Cecal Microbial Composition and the Possible Correlation with Body Weight Gain in Broiler Chickens. Res. Vet. Sci. 2020, 132, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Nicholds, J.F.; McQuain, C.; Hofacre, C.L.; Mathis, G.F.; Fuller, A.L.; Telg, B.E.; Montoya, A.F.; Williams, S.M.; Berghaus, R.D.; Jones, M.K. The Effect of Different Species of Eimeria with Clostridium perfringens on Performance Parameters and Induction of Clinical Necrotic Enteritis in Broiler Chickens. Avian Dis. 2021, 65, 132–137. [Google Scholar] [CrossRef]
- Paiva, D.; McElroy, A. Necrotic Enteritis: Applications for the Poultry Industry. J. Appl. Poult. Res. 2014, 23, 557–566. [Google Scholar] [CrossRef]
- Shojadoost, B.; Vince, A.R.; Prescott, J.F. The Successful Experimental Induction of Necrotic Enteritis in Chickens by Clostridium perfringens: A Critical Review. Vet. Res. 2012, 43, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McReynolds, J.L.; Byrd, J.A.; Anderson, R.C.; Moore, R.W.; Edrington, T.S.; Genovese, K.J.; Poole, T.L.; Kubena, L.F.; Nisbet, D.J. Evaluation of Immunosuppressants and Dietary Mechanisms in an Experimental Disease Model for Necrotic Enteritis. Poult. Sci. 2004, 83, 1948–1952. [Google Scholar] [CrossRef]
- Sersun Calefi, A.; Quinteiro-Filho, W.M.; de Siqueira, A.; Nascimento Lima, A.P.; Gimenes Cruz, D.S.; Queiroz Hazarbassanov, N.; Auciello Salvagni, F.; Borsoi, A.; de Oliveira Massoco Salles Gomes, C.; Maiorka, P.C.; et al. Heat Stress, Eimeria Spp. and C. perfringens Infections Alone or in Combination Modify Gut Th1/Th2 Cytokine Balance and Avian Necrotic Enteritis Pathogenesis. Vet. Immunol. Immunopathol. 2019, 210, 28–37. [Google Scholar] [CrossRef]
- Cooper, K.K.; Songer, J.G. Necrotic Enteritis in Chickens: A Paradigm of Enteric Infection by Clostridium perfringens Type A. Anaerobe 2009, 15, 55–60. [Google Scholar] [CrossRef]
- Annett, C.B.; Viste, J.R.; Chirino-Trejo, M.; Classen, H.L.; Middleton, D.M.; Simko, E. Necrotic Enteritis: Effect of Barley, Wheat and Corn Diets on Proliferation of Clostridium perfringens Type A. Avian Pathol. 2002, 31, 598–601. [Google Scholar] [CrossRef]
- Latorre, J.D.; Hernandez-Velasco, X.; Kuttappan, V.A.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Bielke, L.R.; Prado-Rebolledo, O.F.; Morales, E.; Hargis, B.M.; et al. Selection of Bacillus Spp. for Cellulase and Xylanase Production as Direct-Fed Microbials to Reduce Digesta Viscosity and Clostridium perfringens Proliferation Using an In Vitro Digestive Model in Different Poultry Diets. Front. Vet. Sci. 2015, 2, 25. [Google Scholar] [CrossRef]
- Yang, W.-Y.; Chou, C.-H.; Wang, C. Characterization of Toxin Genes and Quantitative Analysis of NetB in Necrotic Enteritis (NE)-Producing and Non-NE-Producing Clostridium perfringens Isolated from Chickens. Anaerobe 2018, 54, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, M.; Yuan, W.; Song, Z.; Liao, S.; Qi, N.; Li, J.; Lv, M.; Wu, C.; Lin, X.; Hu, J.; et al. Experimental Induction of Necrotic Enteritis with or without Predisposing Factors Using NetB Positive Clostridium perfringens Strains. Gut Pathog. 2021, 13, 68. [Google Scholar] [CrossRef] [PubMed]
- Keyburn, A.L.; Yan, X.-X.; Bannam, T.L.; Van Immerseel, F.; Rood, J.I.; Moore, R.J. Association between Avian Necrotic Enteritis and Clostridium Perfringens Strains Expressing NetB Toxin. Vet. Res. 2010, 41, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, E.; Wang, W.; Gan, L.; Li, Z.; Guo, S.; Guo, Y. Effects of Thymol and Carvacrol Supplementation on Intestinal Integrity and Immune Responses of Broiler Chickens Challenged with Clostridium perfringens. J. Anim. Sci. Biotechnol. 2016, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Van Damme, L.; Cox, N.; Callens, C.; Haesebrouck, F.; Dargatz, M.; Ducatelle, R.; Van Immerseel, F.; Goossens, E.C. Perfringens Challenge Reduces Matrix Metalloproteinase Activity in the Jejunal Mucosa of Eimeria-Infected Broiler Chickens. Vet. Res. 2020, 51, 100. [Google Scholar] [CrossRef]
- Teirlynck, E.; Gussem, M.D.E.; Dewulf, J.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Morphometric Evaluation of “Dysbacteriosis” in Broilers. Avian Pathol. 2011, 40, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Latorre, J.D.; Adhikari, B.; Park, S.H.; Teague, K.D.; Graham, L.E.; Mahaffey, B.D.; Baxter, M.F.A.; Hernandez-Velasco, X.; Kwon, Y.M.; Ricke, S.C.; et al. Evaluation of the Epithelial Barrier Function and Ileal Microbiome in an Established Necrotic Enteritis Challenge Model in Broiler Chickens. Front. Vet. Sci. 2018, 5, 199. [Google Scholar] [CrossRef]
- de Souza, M.; Cicero, C.E.; Menck-Costa, M.F.; Justino, L.; Gerez, J.R.; Baptista, A.A.S.; Bracarense, A.P.F.R.L. Histological Evaluation of the Intestine of Broiler Chickens: Comparison of Three Sampling Methods. Semin. Ciências Agrárias 2021, 42, 3247–3258. [Google Scholar] [CrossRef]
- de Souza, M.; Baptista, A.A.S.; Valdiviezo, M.J.J.; Justino, L.; Menck-Costa, M.F.; Ferraz, C.R.; da Gloria, E.M.; Verri, W.A.; Bracarense, A.P.F.R.L. Lactobacillus Spp. Reduces Morphological Changes and Oxidative Stress Induced by Deoxynivalenol on the Intestine and Liver of Broilers. Toxicon 2020, 185, 203–212. [Google Scholar] [CrossRef]
- Terciolo, C.; Bracarense, A.P.; Souto, P.C.M.C.; Cossalter, A.-M.; Dopavogui, L.; Loiseau, N.; Oliveira, C.A.F.; Pinton, P.; Oswald, I.P. Fumonisins at Doses below EU Regulatory Limits Induce Histological Alterations in Piglets. Toxins 2019, 11, 548. [Google Scholar] [CrossRef] [Green Version]
- Fasina, Y.O.; Lillehoj, H.S. Characterization of Intestinal Immune Response to Clostridium perfringens Infection in Broiler Chickens. Poult. Sci. 2019, 98, 188–198. [Google Scholar] [CrossRef] [PubMed]
- Lacey, J.A.; Stanley, D.; Keyburn, A.L.; Ford, M.; Chen, H.; Johanesen, P.; Lyras, D.; Moore, R.J. Clostridium perfringens-Mediated Necrotic Enteritis Is Not Influenced by the Pre-Existing Microbiota but Is Promoted by Large Changes in the Post-Challenge Microbiota. Vet. Microbiol. 2018, 227, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Toghyani, M.; Kheravii, S.K.; Pineda, L.; Han, Y.; Swick, R.A.; Wu, S.-B. Potential of Blended Organic Acids to Improve Performance and Health of Broilers Infected with Necrotic Enteritis. Anim. Nutr. 2021, 7, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yuan, B.; Yan, X.; Sun, Z.; Lillehoj, H.S.; Lee, Y.; Baldwin-Bott, C.; Li, C. Clostridium perfringens-Induced Host-Pathogen Transcriptional Changes in the Small Intestine of Broiler Chickens. Pathogens 2021, 10, 1607. [Google Scholar] [CrossRef]
- Prescott, J.F.; Smyth, J.A.; Shojadoost, B.; Vince, A. Experimental Reproduction of Necrotic Enteritis in Chickens: A Review. Avian Pathol. 2016, 45, 317–322. [Google Scholar] [CrossRef]
- Wilson, K.M.; Chasser, K.M.; Duff, A.F.; Briggs, W.N.; Latorre, J.D.; Barta, J.R.; Bielke, L.R. Comparison of Multiple Methods for Induction of Necrotic Enteritis in Broilers. I. J. Appl. Poult. Res. 2018, 27, 577–589. [Google Scholar] [CrossRef]
- Stanley, D.; Wu, S.-B.; Rodgers, N.; Swick, R.A.; Moore, R.J. Differential Responses of Cecal Microbiota to Fishmeal, Eimeria and Clostridium perfringens in a Necrotic Enteritis Challenge Model in Chickens. PLoS ONE 2014, 9, e104739. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.F.; Parreira, V.R.; Mehdizadeh Gohari, I.; Lepp, D.; Gong, J. The Pathogenesis of Necrotic Enteritis in Chickens: What We Know and What We Need to Know: A Review. Avian Pathol. 2016, 45, 288–294. [Google Scholar] [CrossRef] [Green Version]
- Van Waeyenberghe, L.; De Gussem, M.; Verbeke, J.; Dewaele, I.; De Gussem, J. Timing of Predisposing Factors Is Important in Necrotic Enteritis Models. Avian Pathol. 2016, 45, 370–375. [Google Scholar] [CrossRef]
- Shini, S.; Aland, R.C.; Bryden, W.L. Avian Intestinal Ultrastructure Changes Provide Insight into the Pathogenesis of Enteric Diseases and Probiotic Mode of Action. Sci. Rep. 2021, 11, 167. [Google Scholar] [CrossRef]
- Zaytsoff, S.J.M.; Lyons, S.M.; Garner, A.M.; Uwiera, R.R.E.; Zandberg, W.F.; Abbott, D.W.; Inglis, G.D. Host Responses to Clostridium perfringens Challenge in a Chicken Model of Chronic Stress. Gut Pathog. 2020, 12, 24. [Google Scholar] [CrossRef]
- Kumar, A.; Toghyani, M.; Kheravii, S.K.; Pineda, L.; Han, Y.; Swick, R.A.; Wu, S.-B. Organic Acid Blends Improve Intestinal Integrity, Modulate Short-Chain Fatty Acids Profiles and Alter Microbiota of Broilers under Necrotic Enteritis Challenge. Anim. Nutr. 2022, 8, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Olkowski, A.A.; Wojnarowicz, C.; Chirino-Trejo, M.; Drew, M.D. Responses of Broiler Chickens Orally Challenged with Clostridium perfringens Isolated from Field Cases of Necrotic Enteritis. Res. Vet. Sci. 2006, 81, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Bortoluzzi, C.; Vieira, B.S.; Hofacre, C.; Applegate, T.J. Effect of Different Challenge Models to Induce Necrotic Enteritis on the Growth Performance and Intestinal Microbiota of Broiler Chickens. Poult. Sci. 2019, 98, 2800–2812. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Guo, Y.; Wang, Z.; Yuan, J. Exogenous Lysozyme Influences Clostridium Perfringens Colonization and Intestinal Barrier Function in Broiler Chickens. Avian Pathol. 2010, 39, 17–24. [Google Scholar] [CrossRef]
- Huang, T.; Gao, B.; Chen, W.-L.; Xiang, R.; Yuan, M.-G.; Xu, Z.-H.; Peng, X.-Y. Temporal Effects of High Fishmeal Diet on Gut Microbiota and Immune Response in Clostridium perfringens-Challenged Chickens. Front. Microbiol. 2018, 9, 2754. [Google Scholar] [CrossRef] [Green Version]
- Chalmers, G.; Bruce, H.L.; Toole, D.L.; Barnum, D.A.; Boerlin, P. Necrotic Enteritis Potential in a Model System Using Clostridium perfringens Isolated from Field Outbreaks. Avian Dis. 2007, 51, 834–839. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M.; Alizadeh, M.; Aldawoodi, A. Effects of CLFchimera Peptide on Intestinal Morphology, Integrity, Microbiota, and Immune Cells in Broiler Chickens Challenged with Necrotic Enteritis. Sci. Rep. 2020, 10, 17704. [Google Scholar] [CrossRef]
- Saitoh, Y.; Suzuki, H.; Tani, K.; Nishikawa, K.; Irie, K.; Ogura, Y.; Tamura, A.; Tsukita, S.; Fujiyoshi, Y. Structural Insight into Tight Junction Disassembly by Clostridium Perfringens Enterotoxin. Science 2015, 347, 775–778. [Google Scholar] [CrossRef]
- Kiu, R.; Hall, L.J. An Update on the Human and Animal Enteric Pathogen Clostridium perfringens. Emerg. Microbes Infect. 2018, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Caly, D.L.; D’Inca, R.; Auclair, E.; Drider, D. Alternatives to Antibiotics to Prevent Necrotic Enteritis in Broiler Chickens: A Microbiologist’s Perspective. Front. Microbiol. 2015, 6, 1336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, P.; Kiess, A.; Adhikari, R.; Jha, R. An Approach to Alternative Strategies to Control Avian Coccidiosis and Necrotic Enteritis. J. Appl. Poult. Res. 2020, 29, 515–534. [Google Scholar] [CrossRef]
- Yegani, M.; Korver, D.R. Factors Affecting Intestinal Health in Poultry. Poult. Sci. 2008, 87, 2052–2063. [Google Scholar] [CrossRef]
- Redondo, L.M.O.; Redondo, E.A.; Delgado, F.O.; La sala, L.F.; Fernandez Myakawa, M.E. An Experimental Reproduction of Necrotic Enteritis in Broiler Chickens. J. Vet. Sci. Med. 2016, 4. [Google Scholar] [CrossRef]
- Sanches, A.W.D.; Belote, B.L.; Hümmelgen, P.; Heemann, A.C.W.; Soares, I.; Tujimoto-Silva, A.; Tirado, A.G.C.; Cunha, A.F.; Santin, E. Basal and Infectious Enteritis in Broilers under the I See Inside Methodology: A Chronological Evaluation. Front. Vet. Sci. 2020, 6, 512. [Google Scholar] [CrossRef] [Green Version]
- Crawley, S.W.; Mooseker, M.S.; Tyska, M.J. Shaping the Intestinal Brush Border. J. Cell Biol. 2014, 207, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonissen, G.; Eeckhaut, V.; Van Driessche, K.; Onrust, L.; Haesebrouck, F.; Ducatelle, R.; Moore, R.J.; Van Immerseel, F. Microbial Shifts Associated with Necrotic Enteritis. Avian Pathol. 2016, 45, 308–312. [Google Scholar] [CrossRef] [Green Version]
- Criado-Mesas, L.; Abdelli, N.; Noce, A.; Farré, M.; Pérez, J.F.; Solà-Oriol, D.; Martin-Venegas, R.; Forouzandeh, A.; González-Solé, F.; Folch, J.M. Transversal Gene Expression Panel to Evaluate Intestinal Health in Broiler Chickens in Different Challenging Conditions. Sci. Rep. 2021, 11, 6315. [Google Scholar] [CrossRef]
- M’Sadeq, S.A.; Wu, S.-B.; Choct, M.; Forder, R.; Swick, R.A. Use of Yeast Cell Wall Extract as a Tool to Reduce the Impact of Necrotic Enteritis in Broilers. Poult. Sci. 2015, 94, 898–905. [Google Scholar] [CrossRef]
- Golder, H.M.; Geier, M.S.; Forder, R.E.A.; Hynd, P.I.; Hughes, R.J. Effects of Necrotic Enteritis Challenge on Intestinal Micro-Architecture and Mucin Profile. Br. Poult. Sci. 2011, 52, 500–506. [Google Scholar] [CrossRef]
- Ierardi, E.; Losurdoa, G.; Iannonea, A.; Piscitellib, D.; Amorusoa, A.; Baronea, M.; Principia, M.; Pisania, A.; Di Leo, A. Lymphocytic Duodenitis or Microscopic Enteritis and Gluten-Related Conditions: What Needs to Be Explored? Ann. Gastroenterol. 2017, 30, 380–392. [Google Scholar] [CrossRef] [PubMed]
- Ruhnke, I.; Andronicos, N.M.; Swick, R.A.; Hine, B.; Sharma, N.; Kheravii, S.K.; Wu, S.-B.; Hunt, P. Immune Responses Following Experimental Infection with Ascaridia galli and Necrotic Enteritis in Broiler Chickens. Avian Pathol. 2017, 46, 602–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, J.; Wang, Z.; Xu, Z.; Pang, Y.; Leng, M.; Tang, S.; Zhang, X.; Qin, J.; Chen, F.; Lin, W. Pathogenicity and Molecular Characterization of Infectious Bursal Disease Virus in China. Poult. Sci. 2022, 101, 101502. [Google Scholar] [CrossRef] [PubMed]
- Gholamiandehkordi, A.R.; Timbermont, L.; Lanckriet, A.; Broeck, W.V.D.; Pedersen, K.; Dewulf, J.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R.; Immerseel, F.V. Quantification of Gut Lesions in a Subclinical Necrotic Enteritis Model. Avian Pathol. 2007, 36, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Collier, C.T.; Hofacre, C.L.; Payne, A.M.; Anderson, D.B.; Kaiser, P.; Mackie, R.I.; Gaskins, H.R. Coccidia-Induced Mucogenesis Promotes the Onset of Necrotic Enteritis by Supporting Clostridium perfringens Growth. Vet. Immunol. Immunopathol. 2008, 122, 104–115. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Groups | Description | n |
|---|---|---|
| G1 | Negative control | 9 |
| G2 | CP * 13 + Eimeria spp. | 9 |
| G3 | CP13 + IBDV-vac ** | 9 |
| G4 | CP13 + Eimeria spp. + IBDV-vac | 9 |
| G5 | CP14 + Eimeria spp. | 9 |
| G6 | CP14 + IBDV-vac | 9 |
| G7 | CP14 + Eimeria spp. + IBDV-vac | 9 |
| G8 | CP03 + Eimeria spp. | 9 |
| G9 | CP03 + IBDV-vac | 9 |
| G10 | CP03 + Eimeria spp. + IBDV-vac | 9 |
| G11 | Eimeria spp. | 9 |
| Total | 99 |
| Groups | Means |
|---|---|
| G1 | 2.75 ± 1.48 a |
| G2 | 2.89 ± 1.57 a |
| G3 | 3.45 ± 0.51 a |
| G4 | 1.67 ± 0.68 b |
| G5 | 2.78 ± 0.94 a |
| G6 | 2.56 ± 1.54 a |
| G7 | 1.78 ± 0.94 b |
| G8 | 3.34 ± 1.08 a |
| G9 | 2.23 ± 0.43 b |
| G10 | 2.43 ± 2.87 a |
| G11 | 2.56 ± 1.09 a |
| Groups | Aerobic | Anaerobic |
|---|---|---|
| G1 | 0.78 ± 0.91 | 0.75 ± 1.20 b |
| G2 | 1.15 ± 0.93 | 2.25 ± 0.52 a |
| G3 | 0.16 ± 0.49 | 1.23 ± 1.18 b |
| G4 | 1.06 ± 0.99 | 2.61 ± 0.32 a |
| G5 | 0.461 ± 0.94 | 2.26 ± 0.38 a |
| G6 | 0.86 ± 1.07 | 2.38 ± 0.49 a |
| G7 | 0.58 ± 0.95 | 2.65 ± 0.71 a |
| G8 | 0.57 ± 0.92 | 1.44 ± 1.58 b |
| G9 | 0.22 ± 0.44 | 0.88 ± 1.53 b |
| G10 | 0.71 ± 1.22 | 2.46 ± 0.81 a |
| G11 | 0.51 ± 0.61 | 0.99 ± 1.23 b |
| Groups | Duodenum | Jejunum | Ileum | |
|---|---|---|---|---|
| iIEL | iIEL | iIEL | Goblet Cells | |
| G1 | 22.6 ± 6.04 b | 18.32 ± 3.68 | 9.38 ± 12.74 | 96.65 ± 17.98 |
| G2 | 26.42 ± 6.36 b | 25.11 ± 13.09 | 6.11 ± 5.13 | 97.53 ± 16.87 |
| G3 | 21.54 ± 5.02 b | 31.83 ± 5.64 | 7.26 ± 1.37 | 90.02 ± 20.13 |
| G4 | 24.76 ± 8.04 b | 24.08 ± 9.45 | 6.96 ± 2.99 | 123.08 ± 8.09 |
| G5 | 33.56 ± 9.35 a | 25.07 ± 11.23 | 5.51 ± 2.67 | 100.89 ± 13.71 |
| G6 | 28.90 ± 5.73 b | 26.19 ± 5.99 | 10.22 ± 4.80 | 100.22 ± 21.71 |
| G7 | 27.12 ± 9.70 b | 32.89 ± 17.20 | 12.05 ± 9.72 | 88.57 ± 12.38 |
| G8 | 38.46 ± 12.76 a | 35 ± 10.76 | 9.85 ± 4.86 | 104.06 ± 10.14 |
| G9 | 28.34 ± 11.65 b | 33.17 ± 9.38 | 8.65 ± 3.81 | 107.95 ± 24.52 |
| G10 | 23.79 ± 12.86 b | 36.24 ± 16.50 | 9.54 ± 6.01 | 102.55 ± 18.33 |
| G11 | 39.54 ± 8.12 a | 31.44 ± 8.32 | 13.32 ± 8.52 | 114.8 ± 15.26 |
| Groups | Duodenum | Jejunum |
|---|---|---|
| G1 | 1.20 ± 0.45 | 1.2 ± 0.83 c |
| G2 | 3.17 ± 1.33 | 5.83 ± 2.79 a |
| G5 | 2.50 ± 2.07 | 4.5 ± 1.52 b |
| G8 | 2.83 ± 1.47 | 8.17 ± 2.79 a |
| G11 | 2.67 ± 1.03 | 8.67 ± 3.20 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Justino, L.; Baptista, A.A.S.; Souza, M.d.; Menck-Costa, M.F.; Pires, B.G.; Cicero, C.E.; Bracarense, A.P.F.R.L.; Kaneko, V.M.; Oba, A.; Okamoto, A.S.; et al. Evaluation of Predisposing Factors of Necrotic Enteritis in Experimentally Challenged Broiler Chickens. Animals 2022, 12, 1880. https://doi.org/10.3390/ani12151880
Justino L, Baptista AAS, Souza Md, Menck-Costa MF, Pires BG, Cicero CE, Bracarense APFRL, Kaneko VM, Oba A, Okamoto AS, et al. Evaluation of Predisposing Factors of Necrotic Enteritis in Experimentally Challenged Broiler Chickens. Animals. 2022; 12(15):1880. https://doi.org/10.3390/ani12151880
Chicago/Turabian StyleJustino, Larissa, Ana A. S. Baptista, Marielen de Souza, Maísa F. Menck-Costa, Bárbara G. Pires, Claudineia E. Cicero, Ana P. F. R. L. Bracarense, Vanessa M. Kaneko, Alexandre Oba, Adriano S. Okamoto, and et al. 2022. "Evaluation of Predisposing Factors of Necrotic Enteritis in Experimentally Challenged Broiler Chickens" Animals 12, no. 15: 1880. https://doi.org/10.3390/ani12151880