
Development and Evaluation of a Commercial Direct-Fed Microbial (Zymospore®) on the Fecal Microbiome and Growth Performance of Broiler Chickens under Experimental Challenge Conditions

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Characterization of Bacillus subtilis Strains
2.2. Direct-Fed Microbial (DFM) Product
2.3. Experiment 1
2.3.1. Housing Conditions
2.3.2. Animal Source and Diets
2.3.3. Experimental Design
2.3.4. Challenge Model to Induce Dysbiosis
2.3.5. Sampling, DNA Extraction, Sequencing, and Bioinformatic Analysis
2.4. Experiment 2
2.4.1. Housing Conditions
2.4.2. Animal Source and Diets
2.4.3. Experimental Design
2.4.4. Litter Filtrate to Recapitulate Commercial Farm Conditions
2.5. Statistical Analysis
3. Results
3.1. Bacillus Species Characterization and Stain Selection
3.2. Experiment 1
3.2.1. Necrotic Enteritis Challenge and Performance Data
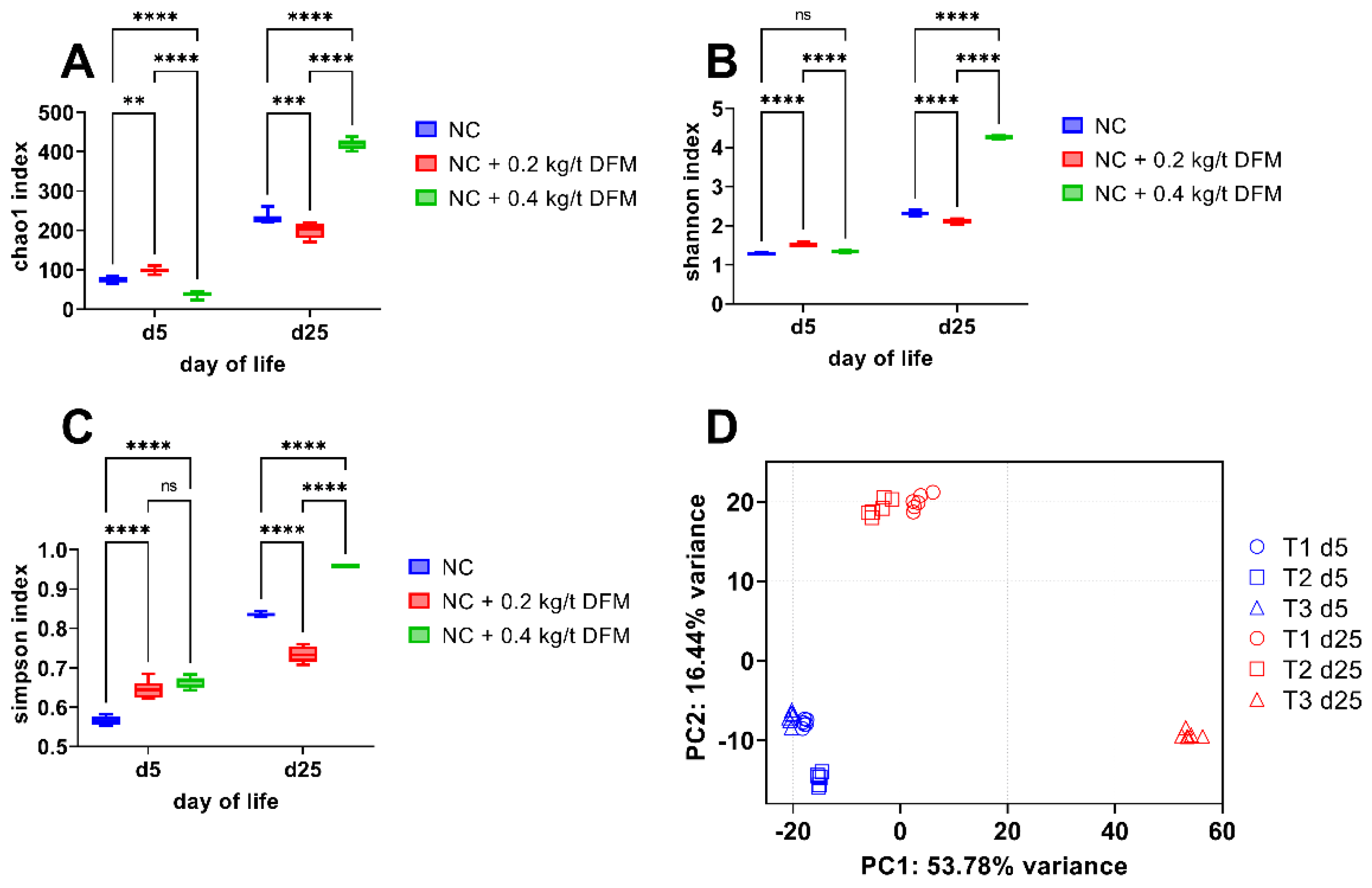
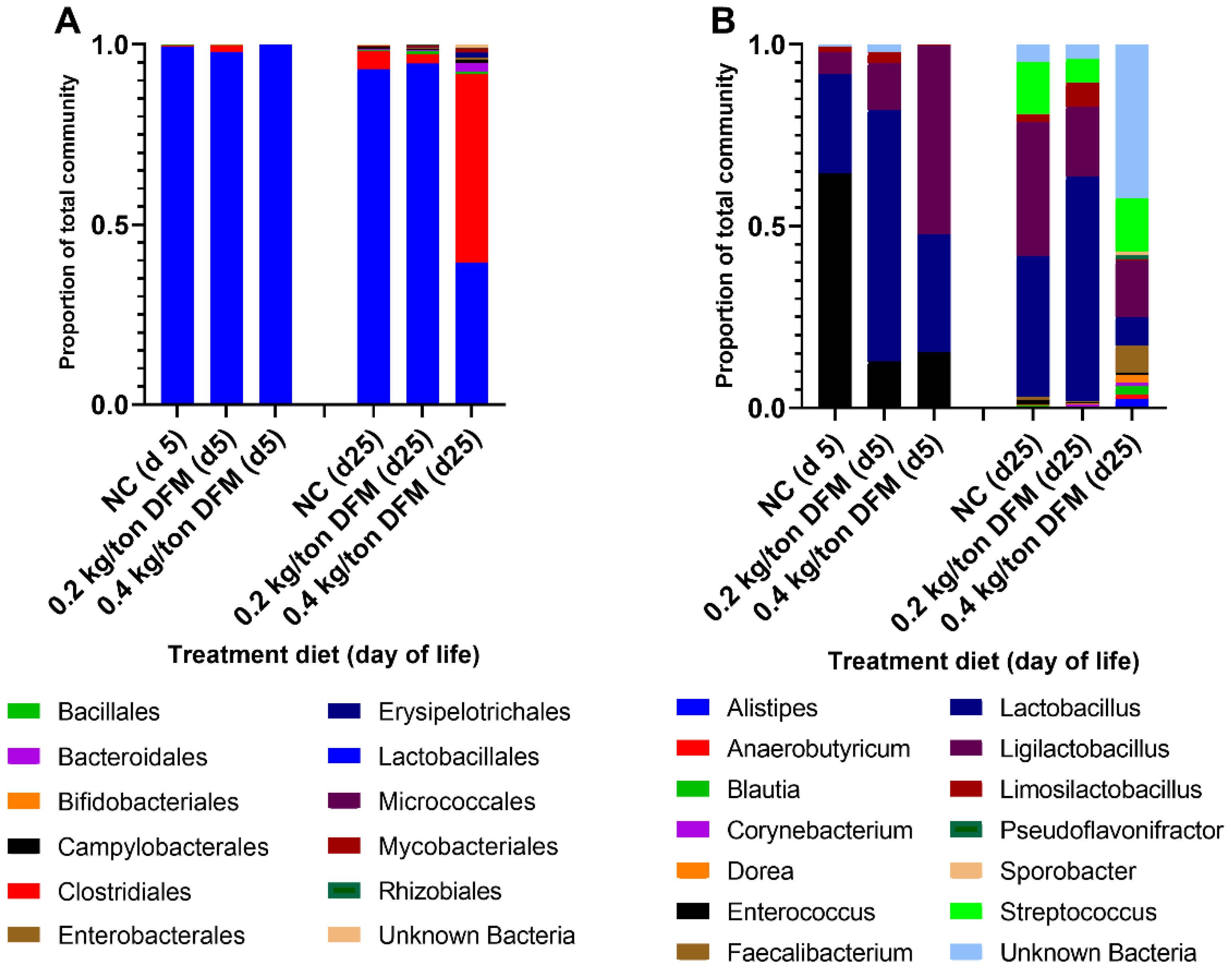
3.2.2. Fecal Microbiome Analysis
3.3. Experiment 2
Intestinal Dysbiosis Challenge and Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Teirlynck, E.; Gussem, M.; Dewulf, J.; Haesebrouck, F.; Ducatelle, R.; Van Immerseel, F. Morphometric evaluation of “dysbacteriosis” in broilers. Avian Pathol. 2011, 40, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Roberts, T.; Wilson, J.; Guthrie, A.; Cookson, K.; Vancraeynest, D.; Schaeffer, J.; Moody, R.; Clark, S. New issues and science in broiler chicken intestinal health: Intestinal microbial composition, shifts, and impacts. Worlds Poult. Sci. J. 2015, 71, 259–270. [Google Scholar] [CrossRef]
- Parker, D.S. Manipulation of the functional activity of the gut by dietary and other means (antibiotics/probiotics) in ruminants. J. Nutr. 1990, 120, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Plata, G.; Baxter, N.T.; Susanti, D.; Volland-Munson, A.; Gangaiah, D.; Nagireddy, A.; Mane, S.P.; Balakuntla, J.; Hawkins, T.B.; Mahajan, M.A. Growth promotion and antibiotic induced metabolic shifts in the chicken gut microbiome. Commun. Biol. 2022, 5, 293. [Google Scholar] [CrossRef] [PubMed]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. probiotic strains as a potential tool for limiting the use of antibiotics, and improving the growth and health of pigs and chickens. Front. Microbiol. 2022, 13, 801827. [Google Scholar] [CrossRef]
- The White House. National Action Plan for Combating Antibiotic-Resistant Bacteria. 2015. Available online: https://www.cdc.gov/drugresistance/pdf/national_action_plan_for_combating_antibotic-resistant_bacteria.pdf (accessed on 13 March 2022).
- Tellez, G.; Latorre, J.D. Alternatives to Antimicrobial Growth Promoters and Their Impact in Gut Microbiota, Health and Disease; Frontiers Media: Lausanne, Switzerland, 2017; pp. 6–92. [Google Scholar] [CrossRef]
- Bai, W.K.; Zhang, F.J.; He, T.J.; Su, P.W.; Ying, X.Z.; Zhang, L.L.; Wang, T. Dietary probiotic Bacillus subtilis strain fmbj increases antioxidant capacity and oxidative stability of chicken breast meat during storage. PLoS ONE 2016, 11, e0167339. [Google Scholar] [CrossRef]
- Pandey, K.R.; Naik, S.R.; Vakil, B.V. Probiotics, prebiotics and synbiotics—A review. J. Food Sci. Technol. 2015, 52, 7577–7587. [Google Scholar] [CrossRef]
- Wang, B.; Zhou, Y.; Tang, L.; Zeng, Z.; Gong, L.; Wu, Y.; Li, W.-F. Effects of Bacillus amyloliquefaciens instead of antibiotics on growth performance, intestinal health, and intestinal microbiota of broilers. Front. Vet. Sci. 2021, 8, 679368. [Google Scholar] [CrossRef]
- Li, C.-L.; Wang, J.; Zhang, H.-J.; Wu, S.-G.; Hui, Q.-R.; Yang, C.-B.; Fang, R.-J.; Qi, G.-H. Intestinal morphologic and microbiota responses to dietary Bacillus spp. in a broiler chicken model. Front. Physiol. 2019, 9, 1968. [Google Scholar] [CrossRef]
- Jadamus, A.; Vahjen, W.; Simon, O. Growth behaviour of a spore-forming probiotic strain in the gastrointestinal tract of broiler chicken and piglets. Arch. Tierernahr. 2001, 54, 1–17. [Google Scholar] [CrossRef]
- Tellez, G.; Pixley, C.; Wolfenden, R.; Layton, S.; Hargis, B.M. Probiotics/direct-fed microbials for Salmonella control in poultry. Food Res. Int. 2012, 45, 628–633. [Google Scholar] [CrossRef]
- Menconi, A.; Morgan, M.J.; Pumford, N.R.; Hargis, B.M.; Tellez, G. Physiological properties and Salmonella growth inhibition of probiotic Bacillus strains isolated from environmental and poultry sources. Int. J. Bacteriol. 2013, 2013, 958408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vreeland, R.H.; Rosenzweig, W.D.; Powers, D.W. Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal. Nature 2000, 407, 897–900. [Google Scholar] [CrossRef] [PubMed]
- Shivaramaiah, S.; Pumford, N.; Morgan, M.; Wolfenden, R.; Wolfenden, A.; Torres-Rodriguez, A.; Hargis, B.M.; Téllez, G. Evaluation of Bacillus species as potential candidates for direct-fed microbials in commercial poultry. Poult. Sci. 2011, 90, 1574–1580. [Google Scholar] [CrossRef] [PubMed]
- Cartman, S.T.; La Ragione, R.M.; Woodward, M.J. Bacillus subtilis spores germinate in the chicken gastrointestinal tract. Appl. Environ. Microbiol. 2008, 74, 5254–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casula, G.; Cutting, S.M. Bacillus probiotics: Spore germination in the gastrointestinal tract. Appl. Environ. Microbiol. 2002, 68, 2344–2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Gueye, S.A.; Lupetti, A.; Senesi, S. Survival and persistence of Bacillus clausii in the human gastrointestinal tract following oral administration as spore-based probiotic formulation. J. Appl. Microbiol. 2015, 119, 552–559. [Google Scholar] [CrossRef]
- Duc, L.H.; Hong, H.A.; Barbosa, T.M.; Henriques, A.O.; Cutting, S.M. Characterization of Bacillus probiotics available for human use. Appl. Environ. Microbiol. 2004, 70, 2161–2171. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, C.W.; Doyle, J.D.; Hugley, B. A new solid medium for enumerating cellulose-utilizing bacteria in soil. Appl. Environ. Microbiol. 1995, 61, 2016–2019. [Google Scholar] [CrossRef] [Green Version]
- Mazotto, A.M.; Rodrigues, R.R.C.; Cedrola, S.M.L.; de Lima, M.F.; Couri, S.; Paraguai de Souza, E.; Vermelho, A.B. Keratinase production by three Bacillus spp. using feather meal and whole feather as substrate in a submerged fermentation. Enzym. Res. 2011, 2011, 523780. [Google Scholar] [CrossRef] [Green Version]
- Latorre, J.D.; Hernandez-Velasco, X.; Kuttappan, V.A.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Bielke, L.R.; Prado-Rebolledo, O.F.; Morales, E.; Hargis, B.M.; et al. Selection of Bacillus spp. for cellulase and xylanase production as direct-fed microbials to reduce digesta viscosity and Clostridium perfringens proliferation using an in vitro digestive model in different poultry diets. Front. Vet. Sci. 2015, 2, 25. [Google Scholar] [CrossRef] [PubMed]
- Latorre, J.D.; Hernandez-Velasco, X.; Wolfenden, R.E.; Vicente, J.L.; Wolfenden, A.D.; Menconi, A.; Bielke, L.R.; Hargis, B.M.; Tellez, G. Evaluation and selection of Bacillus species based on enzyme production, antimicrobial activity, and biofilm synthesis as direct-fed microbial candidates for poultry. Front. Vet. Sci. 2016, 3, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latorre, J.; Hernandez-Velasco, X.; Vicente, J.; Wolfenden, R.; Hargis, B.; Tellez, G. Effects of the inclusion of a Bacillus direct-fed microbial on performance parameters, bone quality, recovered gut microflora, and intestinal morphology in broilers consuming a grower diet containing corn distillers dried grains with solubles. Poult. Sci. 2017, 96, 2728–2735. [Google Scholar] [CrossRef] [PubMed]
- Hosoi, T.; Ametani, A.; Kiuchi, K.; Kaminogawa, S. Improved growth and viability of lactobacilli in the presence of Bacillus subtilis (natto), catalase, or subtilisin. Can. J. Microbiol. 2000, 46, 892–897. [Google Scholar] [CrossRef]
- Jani, S.A.; Chudasama, C.J.; Patel, D.B.; Bhatt, P.S.; Patel, H.N. Optimization of extracellular protease production from alkali thermo tolerant actinomycetes: Saccharomonospora viridis SJ-21. Bull. Environ. Pharmacol. Life Sci. 2012, 1, 84–92. [Google Scholar]
- Rostagno, H.S.; Albino, L.F.T.; Hannas, M.I.; Donzele, J.L.; Sakomura, N.K.; Perazzo, F.G.; Saraiva, A.; Texeira de Abreu, M.L.; Rodrigues, P.B.; Oliveira, R.F.; et al. Tabelas Brasileiras para Aves e Suínos. Composição de Alimentos e Exigências Nutricionais, 4th ed.; Universidade Federal deViçosa: Viçosa, Brazil, 2017. [Google Scholar]
- Belote, B.; Tujimoto-Silva, A.; Hümmelgen, P.; Sanches, A.; Wammes, J.; Hayashi, R.; Santin, E. Histological parameters to evaluate intestinal health on broilers challenged with Eimeria and Clostridium perfringens with or without enramycin as growth promoter. Poult. Sci. 2018, 97, 2287–2294. [Google Scholar] [CrossRef]
- Stefanello, C.; Rosa, D.P.; Dalmoro, Y.K.; Segatto, A.L.; Vieira, M.S.; Moraes, M.L.; Santin, E. Protected blend of organic acids and essential oils improves growth performance, nutrient digestibility, and intestinal health of broiler chickens undergoing an intestinal challenge. Front. Vet. Sci. 2020, 6, 491. [Google Scholar] [CrossRef]
- Degnan, P.H.; Ochman, H. Illumina-based analysis of microbial community diversity. ISME J. 2012, 6, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Sakomura, N.K.; Rostagno, H.S. Métodos de Pesquisa em Nutrição de Monogástricos; Fundação de Apoio a Pesquisa, Ensino e Extensão: Jaboticabal, Brazil, 2007; 262p. [Google Scholar]
- SAS Institute Inc. SAS User Guide. Version 9.1; SAS Institute Inc.: Carry, NC, USA, 2002. [Google Scholar]
- Hernandez-Patlan, D.; Solis-Cruz, B.; Pontin, K.P.; Hernandez-Velasco, X.; Merino-Guzman, R.; Adhikari, B.; López-Arellano, R.; Kwon, Y.M.; Hargis, B.M.; Arreguin-Nava, M.A.; et al. Impact of a Bacillus direct-fed microbial on growth performance, intestinal barrier integrity, necrotic enteritis lesions, and ileal microbiota in broiler chickens using a laboratory challenge model. Front. Vet. Sci. 2019, 6, 108. [Google Scholar] [CrossRef]
- Palliyeguru, M.W.C.D.; Rose, S.P. Sub-clinical necrotic enteritis: Its aetiology and predisposing factors in commercial broiler production. Worlds Poult. Sci. J. 2014, 70, 803–816. [Google Scholar] [CrossRef]
- Abriouel, H.; Franz, C.M.A.P.; Omar, N.B.; Gálvez, A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiol. Rev. 2011, 35, 201–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priest, F.G. Extracellular enzyme synthesis in the genus Bacillus. Bacteriol. Rev. 1977, 41, 711–753. [Google Scholar] [CrossRef]
- Lee, K.W.; Kim, D.K.; Lillehoj, H.S.; Jang, S.I.; Lee, S.H. Immune modulation by Bacillus subtilis-based direct-fed microbials in commercial broiler chickens. Anim. Feed Sci. Technol. 2015, 200, 76–85. [Google Scholar] [CrossRef]
- Sandvang, D.; Skjoet-Rasmussen, L.; Cantor, M.D.; Mathis, G.F.; Lumpkins, B.S.; Blanch, A. Effects of feed supplementation with 3 different probiotic Bacillus strains and their combination on the performance of broiler chickens challenged with Clostridium perfringens. Poult. Sci. 2021, 100, 100982. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, B.; Hernandez-Patlan, D.; Solis-Cruz, B.; Kwon, Y.M.; Arreguin, M.A.; Latorre, J.D.; Hernandez-Velasco, X.; Hargis, B.M.; Tellez-Isaias, G. Evaluation of the antimicrobial and anti-inflammatory properties of Bacillus-DFM (Norum™) in broiler chickens infected with Salmonella Enteritidis. Front. Vet. Sci. 2019, 6, 282. [Google Scholar] [CrossRef] [Green Version]
- Shehata, A.A.; Yalçın, S.; Latorre, J.D.; Basiouni, S.; Attia, Y.A.; Abd El-Wahab, A.; Visscher, C.; El-Seedi, H.R.; Huber, C.; Hafez, H.M.; et al. Probiotics, prebiotics, and phytogenic substances for optimizing gut health in poultry. Microorganisms 2022, 10, 395. [Google Scholar] [CrossRef]
- Hong, H.A.; Duc, L.H.; Cutting, S.M. The use of bacterial spore formers as probiotics. FEMS Microbiol. Rev. 2005, 29, 813–835. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Huang, J.M.; Khaneja, R.; Hiep, L.; Urdaci, M.; Cutting, S. The safety of Bacillus subtilis and Bacillus indicus as food probiotics. J. Appl. Microbiol. 2008, 105, 510–520. [Google Scholar] [CrossRef]
- Amuguni, H.; Tzipori, S. Bacillus subtilis: A temperature resistant and needle free delivery system of immunogens. Hum. Vaccin. Immunother. 2012, 8, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Penaloza-Vazquez, A.; Ma, L.M.; Rayas-Duarte, P. Isolation and characterization of Bacillus spp. strains as potential probiotics for poultry. Can. J. Microbiol. 2019, 65, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Nithya, V.; Halami, P.M. Evaluation of the probiotic characteristics of Bacillus species isolated from different food sources. Ann. Microbiol. 2013, 63, 129–137. [Google Scholar] [CrossRef]
- Stein, T. Bacillus subtilis antibiotics: Structures, syntheses and specific functions. Mol. Microbiol. 2005, 56, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Monisha, R.; Uma, M.; Murthy, V.K. Partial purification and characterization of Bacillus pumilus xylanase from soil source. KUSET 2009, 5, 137–148. [Google Scholar]
- Latorre, J.D.; Hernandez-Velasco, X.; Kogut, M.H.; Vicente, J.L.; Wolfenden, R.; Wolfenden, A.; Hargis, B.M.; Kuttappan, V.A.; Tellez, G. Role of a Bacillus subtilis direct-fed microbial on digesta viscosity, bacterial translocation, and bone mineralization in turkey poults fed with a rye-based diet. Front. Vet. Sci. 2014, 1, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.M.; Shahm, T.M.; Reddy, B.; Deshpande, S.; Rank, D.N.; Joshi, C.G. Taxonomic and gene-centric metagenomics of the fecal microbiome of low and high feed conversion ratio (FCR) broilers. J. Appl. Genet. 2014, 55, 145–154. [Google Scholar] [CrossRef]
- Singh, K.M.; Shah, T.; Deshpande, S.; Jakhesara, S.J.; Koringa, P.G.; Rank, D.N.; Joshi, C.G. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Mol. Biol. Rep. 2012, 39, 10595–10602. [Google Scholar] [CrossRef]
- Díaz-Sánchez, S.; Perrotta, A.R.; Rockafellow, I.; Alm, E.J.; Okimoto, R.; Hawken, R.; Hanning, I. Using fecal microbiota as biomarkers for predictions of performance in the selective breeding process of pedigree broiler breeders. PLoS ONE 2019, 14, e0216080. [Google Scholar] [CrossRef] [Green Version]
- La Ragione, R.M.; Woodward, M.J. Competitive exclusion by Bacillus subtilis spores of Salmonella enterica serotype Enteritidis and Clostridium perfringens in young chickens. Vet. Microbiol. 2003, 94, 245–256. [Google Scholar] [CrossRef]
- Revolledo, L.; Ferreira, A.; Mead, G. Prospects in Salmonella control: Competitive exclusion, probiotics, and enhancement of avian intestinal immunity. J. Appl. Poult. Res. 2006, 15, 341–351. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. Gut microbiota of laying hens and its manipulation with prebiotics and probiotics to enhance gut health and food safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strain | Form | Texture | Surface | Color | Elevation | Size | Margins |
|---|---|---|---|---|---|---|---|
| BS-009 | Irregular | Rough | Dry | Yellowish white | Flat | 0.5–1 cm | Curled |
| BS-020 | Irregular | Rough | Dry | Yellowish white | Flat | 0.5–1 cm | Curled |
| BS-024 | Irregular | Rough | Mucoid | Yellowish white | Flat | 0.5–1 cm | Curled/Lobate |
| Item | Pre-Starter (1 to 7 d) | Starter (7 to 21 d) | Grower (21 to 35 d) | Finisher (35 to 42 d) |
|---|---|---|---|---|
| Ingredients, % | ||||
| Corn | 48.41 | 50.63 | 57.05 | 63.43 |
| Soybean meal | 44.03 | 41.71 | 35.31 | 29.96 |
| Soybean oil | 3.74 | 4.35 | 4.70 | 4.22 |
| Dicalcium phosphate | 1.13 | 0.83 | 0.64 | 0.29 |
| Limestone | 1.38 | 1.23 | 1.14 | 0.97 |
| Salt | 0.55 | 0.52 | 0.50 | 0.47 |
| DL-Met, 99% | 0.35 | 0.33 | 0.28 | 0.24 |
| L-Lys HCl, 78% | 0.13 | 0.13 | 0.15 | 0.17 |
| L-Thr, 98.5% | 0.05 | 0.05 | 0.04 | 0.03 |
| Choline chloride, 60% | 0.03 | 0.04 | 0.05 | 0.07 |
| Vitamin and mineral premix 1 | 0.21 | 0.19 | 0.16 | 0.16 |
| Nutrient and energy composition, % or as shown | ||||
| ME, Mcal/kg | 2.97 | 3.05 | 3.15 | 3.20 |
| Crude protein | 24.16 | 23.27 | 20.86 | 18.92 |
| Ca | 1.01 | 0.88 | 0.79 | 0.63 |
| Av. P | 0.48 | 0.42 | 0.37 | 0.30 |
| Na | 0.23 | 0.22 | 0.21 | 0.20 |
| Cl | 0.42 | 0.41 | 0.40 | 0.40 |
| Choline, mg/kg | 1600 | 1600 | 1500 | 1500 |
| Lys dig. 2 | 1.31 | 1.26 | 1.12 | 1.01 |
| Met + Cys dig. | 0.66 | 0.64 | 0.56 | 0.50 |
| Thr dig. | 0.98 | 0.94 | 0.84 | 0.76 |
| Trp dig. | 0.86 | 0.83 | 0.74 | 0.67 |
| Arg dig. | 0.28 | 0.27 | 0.24 | 0.21 |
| Val dig. | 1.53 | 1.47 | 1.29 | 1.15 |
| Ile dig. | 1.01 | 0.97 | 0.87 | 0.78 |
| Leu dig. | 0.95 | 0.91 | 0.80 | 0.72 |
| Item | Pre-Starter (1 to 14 d) | Finisher (15 to 42 d) |
|---|---|---|
| Ingredients (%) | ||
| Corn | 54.75 | 57.99 |
| Soybean flour 46% | 20.15 | 0.00 |
| Deactivated soybean | 0.00 | 16.00 |
| Soybean expeller | 20.00 | 18.14 |
| Wheat | 0.68 | 4.79 |
| Grit | 1.39 | 1.06 |
| Salt | 0.42 | 0.40 |
| Mycotoxin binder | 0.30 | 0.30 |
| Dicalcium phosphate | 1.16 | 0.54 |
| Lysine | 0.25 | 0.18 |
| Methionine powder | 0.37 | 0.25 |
| Threonine | 0.08 | 0.03 |
| Choline chloride | 0.10 | 0.08 |
| Trace mineral premix 1 | 0.10 | 0.10 |
| Vitamin premix 2 | 0.15 | 0.10 |
| Nutrient and energy composition, % or as shown | ||
| ME, Mcal/kg | 2.95 | 3.10 |
| Crude protein | 21 | 18 |
| Ca | 1.02 | 0.82 |
| Av. P | 0.45 | 0.42 |
| Lys dig. | 1.2 | 1.0 |
| Met dig. | 0.48 | 0.40 |
| Strain | Cellulase | Amylase | Lipases | Proteases |
|---|---|---|---|---|
| BS-009 | 3.55 ± 0.34 | 1.95 ± 0.08 | 3.16 ± 0.10 | 2.33 ± 0.09 |
| BS-020 | 2.02 ± 0.08 | 1.71 ± 0.13 | 3.12 ± 0.40 | 1.96 ± 0.11 |
| BS-024 | 2.30 ± 0.19 | 1.38 ± 0.05 | 2.32 ± 0.19 | 2.51 ± 0.16 |
| Strain | 15 °C | 37 °C | 45 °C | |||
|---|---|---|---|---|---|---|
| 2 h | 4 h | 2 h | 4 h | 2 h | 4 h | |
| BS-009 | 7.03 ± 0.26 | 7.13 ± 0.32 | 7.42 ± 0.10 | 7.40 ± 0.30 | 7.20 ± 0.17 | 6.77 ± 0.68 |
| BS-020 | 6.97 ± 0.06 | 7.20 ± 0.35 | 6.40 ± 0.17 | 6.30 ± 0.30 | 7.30 ± 0.0 | 7.55 ± 0.81 |
| BS-024 | 7.26 ± 0.24 | 7.16 ± 0.15 | 6.95 ± 0.09 | 6.95 ± 0.09 | 7.10 ± 0.17 | 6.93 ± 0.13 |
| Strain | NaCl 3.5% | NaCl 6.5% | ||
|---|---|---|---|---|
| 2 h | 4 h | 2 h | 4 h | |
| BS-009 | 7.14 ± 0.15 | 7.33 ± 0.35 | 6.92 ± 0.08 | 6.77 ± 0.07 |
| BS-020 | 7.28 ± 0.04 | 7.15 ± 0.32 | 7.36 ± 0.10 | 7.03 ± 0.05 |
| BS-024 | 7.12 ± 0.21 | 7.15 ± 0.15 | 6.96 ± 0.34 | 6.73 ± 0.15 |
| Strain | pH2 | pH3 | ||
|---|---|---|---|---|
| 2 h | 4 h | 2 h | 4 h | |
| BS-009 | 6.93 ± 0.08 | 7.33 ± 0.61 | 7.32 ± 0.28 | 6.87 ± 0.11 |
| BS-020 | 7.16 ± 0.28 | 7.14 ± 0.29 | 7.01 ± 0.02 | 6.62 ± 0.54 |
| BS-024 | 6.95 ± 0.09 | 6.67 ± 0.58 | 6.98 ± 0.03 | 6.95 ± 0.05 |
| Item | 1 d | 7 d | 14 d | 21 d | 28 d | 35 d | 42 d | ADG 1 |
|---|---|---|---|---|---|---|---|---|
| Body weight (g) | ||||||||
| Negative control | 45 | 200 | 526 | 990 | 1747 | 2449 | 3294 | 77.35 |
| DFM, 0.2 kg/t | 45 | 199 | 531 | 1008 | 1767 | 2482 | 3340 | 78.45 |
| DFM, 0.3 kg/t | 45 | 205 | 543 | 1026 | 1805 | 2537 | 3402 | 79.93 |
| DFM, 0.4 kg/t | 45 | 200 | 536 | 1014 | 1772 | 2497 | 3353 | 78.77 |
| Positive control | 45 | 196 | 537 | 1006 | 1753 | 2462 | 3328 | 78.15 |
| SEM 2 | 0.49 | 5.72 | 18.70 | 26.33 | 36.02 | 73.29 | 77.10 | 1.834 |
| p-value | 0.8416 | 0.1843 | 0.6004 | 0.2720 | 0.0766 | 0.3002 | 0.2116 | 0.2103 |
| Body weight gain (g/d/b) | 1–7 d | 7–14 d | 14–21 d | 21–28 d | 29–35 d | 35–42 d | ||
| Negative control | 155 | 326 | 554 | 667 | 702 | 845 | ||
| DFM, 0.2 kg/t | 154 | 331 | 568 | 668 | 715 | 858 | ||
| DFM, 0.3 kg/t | 160 | 338 | 573 | 689 | 731 | 865 | ||
| DFM, 0.4 kg/t | 155 | 335 | 568 | 668 | 725 | 856 | ||
| Positive control | 151 | 341 | 559 | 656 | 710 | 865 | ||
| SEM | 5.66 | 15.01 | 16.33 | 33.50 | 76.68 | 57.46 | ||
| p-value | 0.1596 | 0.4425 | 0.2720 | 0.5449 | 0.9644 | 0.9702 | ||
| Feed intake (g/b) | 1–7 d | 7–14 d | 14–21 d | 21–28 d | 29–35 d | 35–42 d | ||
| Negative control | 182 | 426 | 738 | 989 | 1186 | 1298 | ||
| DFM, 0.2 kg/t | 176 | 424 | 753 | 965 | 1165 | 1307 | ||
| DFM, 0.3 kg/t | 182 | 432 | 757 | 965 | 1193 | 1300 | ||
| DFM, 0.4 kg/t | 180 | 430 | 758 | 960 | 1186 | 1301 | ||
| Positive control | 173 | 433 | 739 | 963 | 1184 | 1312 | ||
| SEM | 7.14 | 15.95 | 25.12 | 74.31 | 142.10 | 87.72 | ||
| p-value | 0.1240 | 0.8236 | 0.4647 | 0.9574 | 0.9979 | 0.9987 | ||
| FCR 3 | 1–7 d | 7–14 d | 14–21 d | 21–28 d | 29–35 d | 35–42 d | ||
| Negative control | 1.171 | 1.310 | 1.333 | 1.483 | 1.696 | 1.542 | ||
| DFM, 0.2 kg/t | 1.142 | 1.282 | 1.325 | 1.443 | 1.637 | 1.530 | ||
| DFM, 0.3 kg/t | 1.143 | 1.279 | 1.321 | 1.400 | 1.631 | 1.502 | ||
| DFM, 0.4 kg/t | 1.156 | 1.283 | 1.335 | 1.437 | 1.640 | 1.519 | ||
| Positive control | 1.142 | 1.271 | 1.320 | 1.464 | 1.664 | 1.516 | ||
| SEM | 0.0235 | 0.0349 | 0.0237 | 0.0694 | 0.1368 | 0.0929 | ||
| p-value | 0.1742 | 0.4011 | 0.7216 | 0.3314 | 0.9188 | 0.9589 | ||
| Mortality (%) | 1–7 d | 7–14 d | 14–21 d | 21–28 d | 29–35 d | 35–42 d | Total mortality | |
| Negative control | 0.67 | 0.67 | 0.00 | 0.76 | 0.72 | 0.67 | 3.51 | |
| DFM, 0.2 kg/t | 0.00 | 2.17 | 0.67 | 0.00 | 0.76 | 0.67 | 4.29 | |
| DFM, 0.3 kg/t | 0.67 | 2.17 | 0.67 | 2.25 | 0.00 | 0.67 | 6.51 | |
| DFM, 0.4 kg/t | 0.00 | 0.67 | 0.00 | 0.72 | 0.76 | 0.00 | 2.18 | |
| Positive control | 0.67 | 2.17 | 0.00 | 0.00 | 0.79 | 0.00 | 3.70 | |
| SEM | 1.26 | 3.05 | 1.03 | 1.59 | 1.66 | 1.26 | 4.73 | |
| p-value | 0.7359 | 0.7826 | 0.5674 | 0.1272 | 0.9063 | 0.7359 | 0.6174 | |
| BW g/broiler (d 42) | FI g/broiler (d 1–42) | FCR 1 (d 1–42) | Total Mortality (d 1–42) | |
|---|---|---|---|---|
| Basal diet without filtrate (NC−) | 2810 b | 5302 | 1.97 a | 5/100 (5.00%) |
| BMD without filtrate (BMD−) | 3161 a | 5319 | 1.76 b | 3/100 (3.00%) |
| BMD with filtrate (BMD+) | 3055 a | 5402 | 1.82 b | 3/100 (3.00%) |
| DFM with filtrate (DFM+) | 3108 a | 5456 | 1.83 b | 3/100 (3.00%) |
| SEM 2 | 110 | 3891 | 0.04 | |
| p-value | 0.0011 | 0.6784 | 0.0007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza, O.F.; Vecchi, B.; Gumina, E.; Matté, F.; Gazoni, F.L.; Hernandez-Velasco, X.; Hall, J.W.; Stefanello, C.; Layton, S. Development and Evaluation of a Commercial Direct-Fed Microbial (Zymospore®) on the Fecal Microbiome and Growth Performance of Broiler Chickens under Experimental Challenge Conditions. Animals 2022, 12, 1436. https://doi.org/10.3390/ani12111436
de Souza OF, Vecchi B, Gumina E, Matté F, Gazoni FL, Hernandez-Velasco X, Hall JW, Stefanello C, Layton S. Development and Evaluation of a Commercial Direct-Fed Microbial (Zymospore®) on the Fecal Microbiome and Growth Performance of Broiler Chickens under Experimental Challenge Conditions. Animals. 2022; 12(11):1436. https://doi.org/10.3390/ani12111436
Chicago/Turabian Stylede Souza, Otoniel F., Bruno Vecchi, Emanuel Gumina, Fabrizio Matté, Fabio L. Gazoni, Xochitl Hernandez-Velasco, Jeffrey W. Hall, Catarina Stefanello, and Sherry Layton. 2022. "Development and Evaluation of a Commercial Direct-Fed Microbial (Zymospore®) on the Fecal Microbiome and Growth Performance of Broiler Chickens under Experimental Challenge Conditions" Animals 12, no. 11: 1436. https://doi.org/10.3390/ani12111436