
Experimental Study on Primary Bird Co-Infection with Two Plasmodium relictum Lineages—pSGS1 and pGRW11


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statements
2.2. Obtained Parasites
2.3. Experimental Procedures
2.4. Microscopy of Blood Smears and Molecular Analysis
2.5. Statistics
3. Results
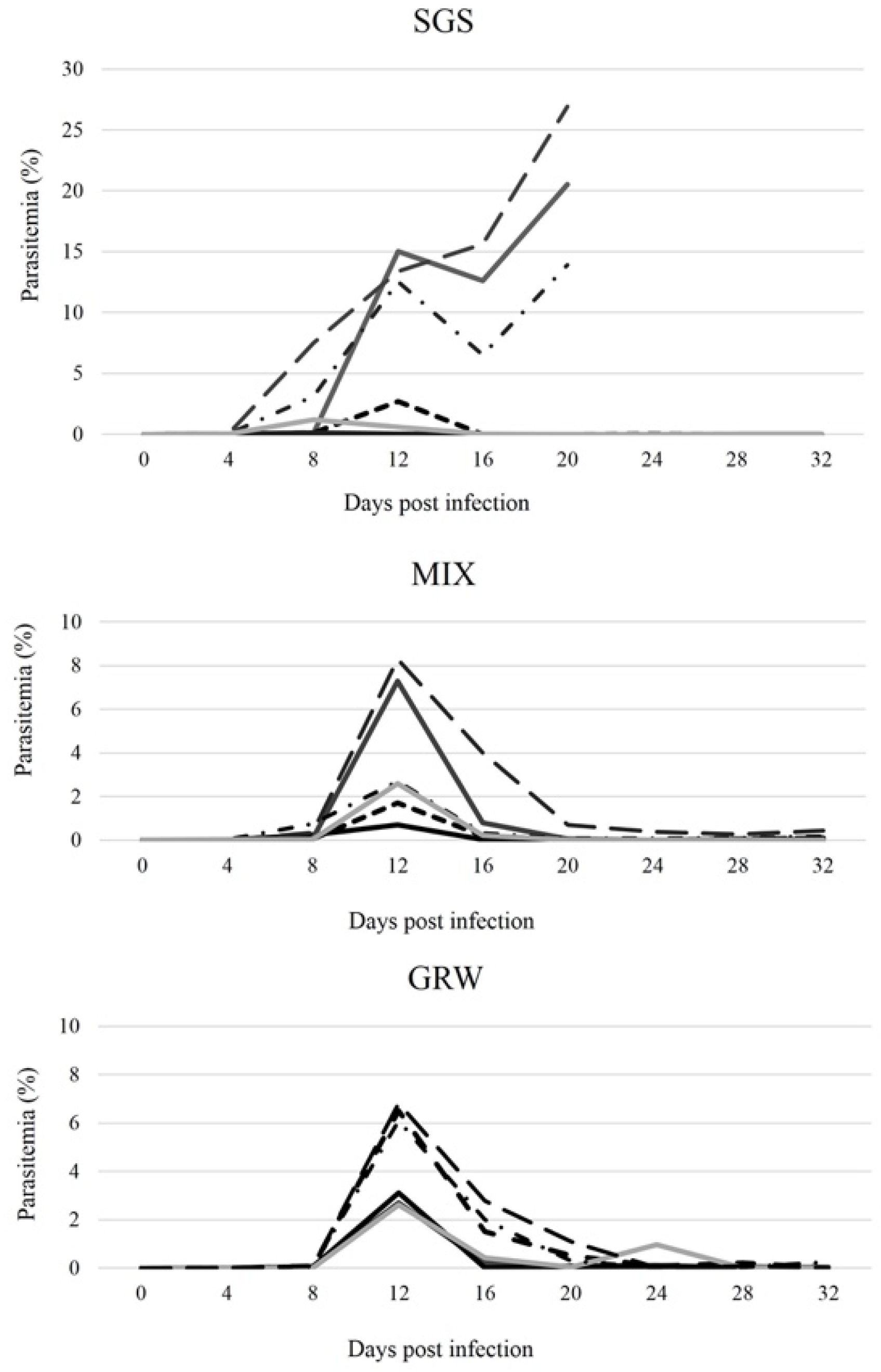
3.1. Development of Parasites within Experimentally Infected Canaries
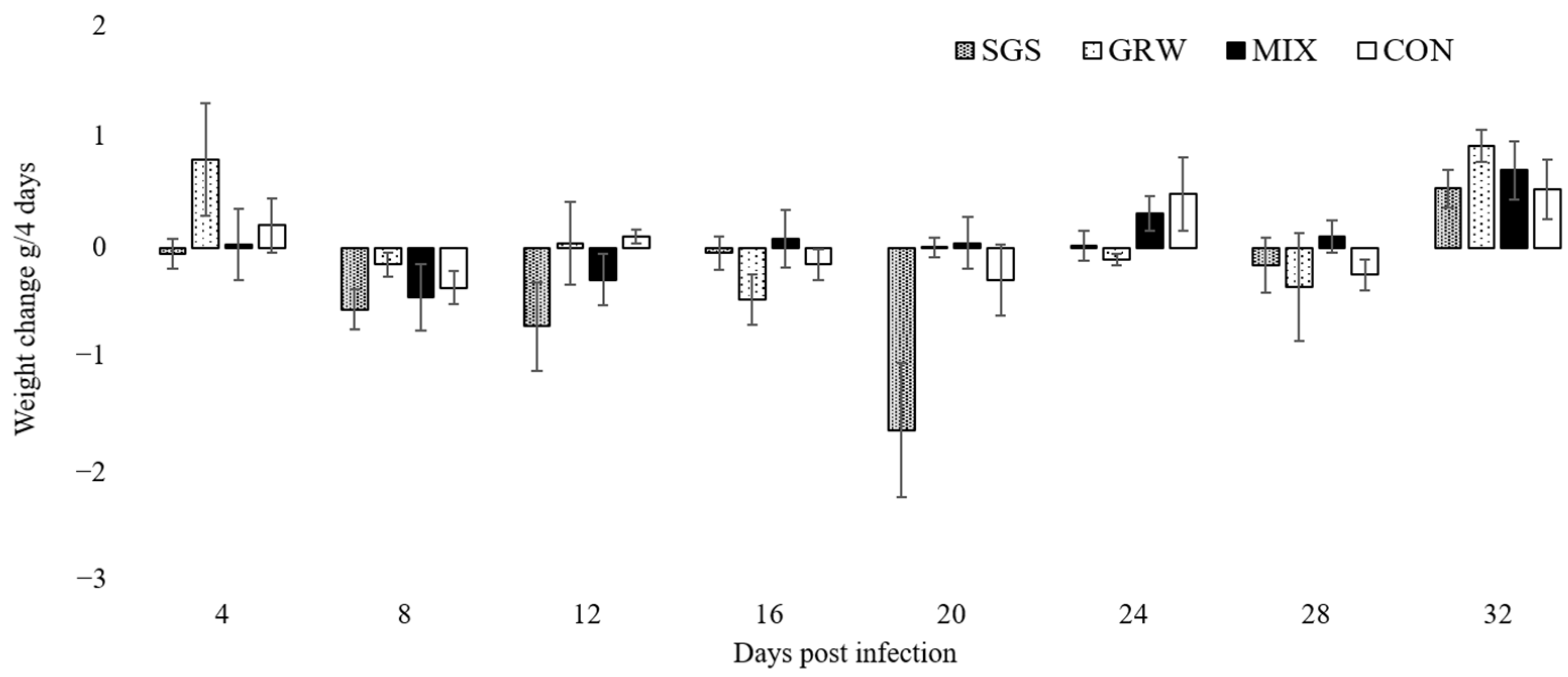
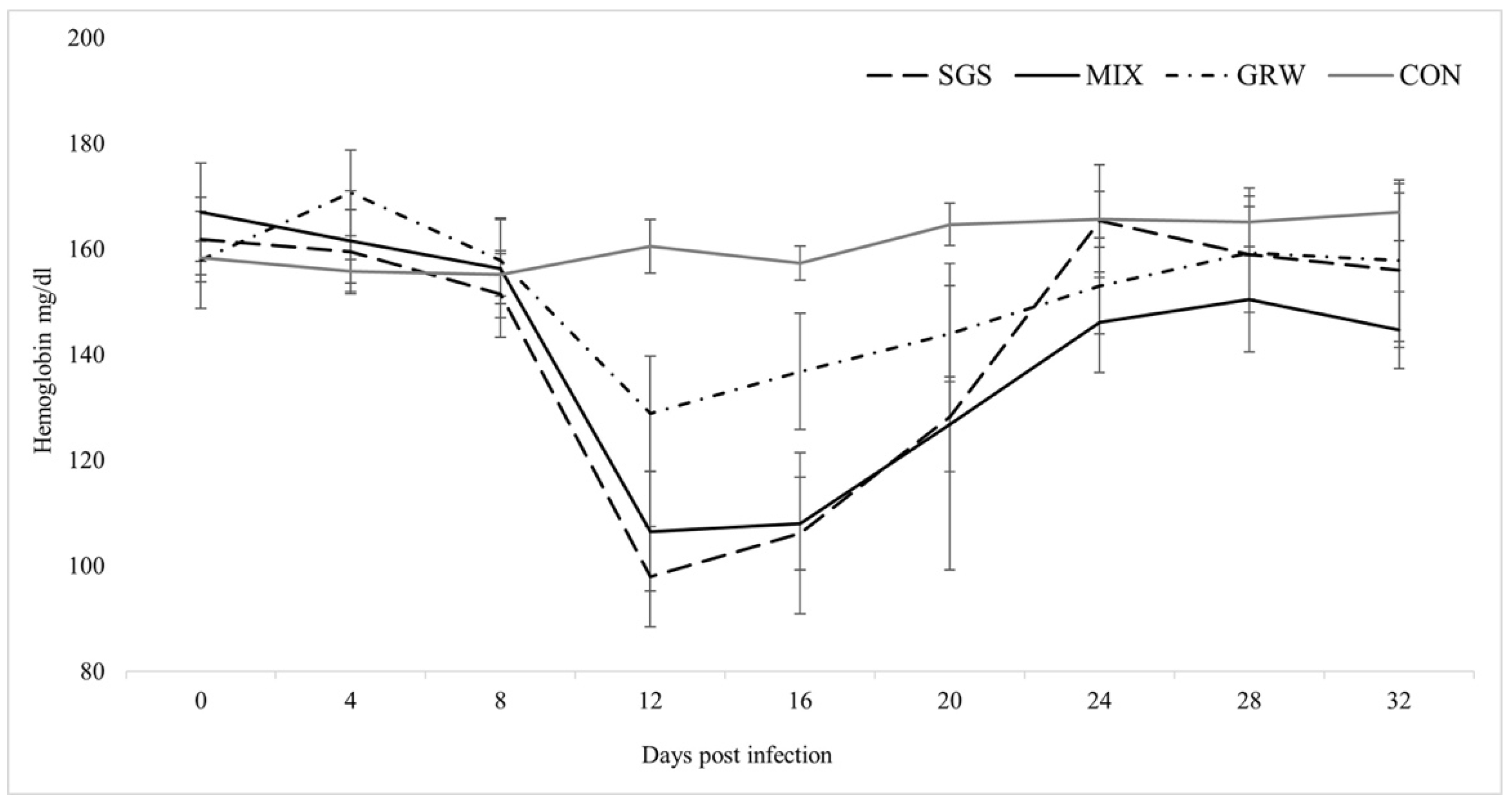
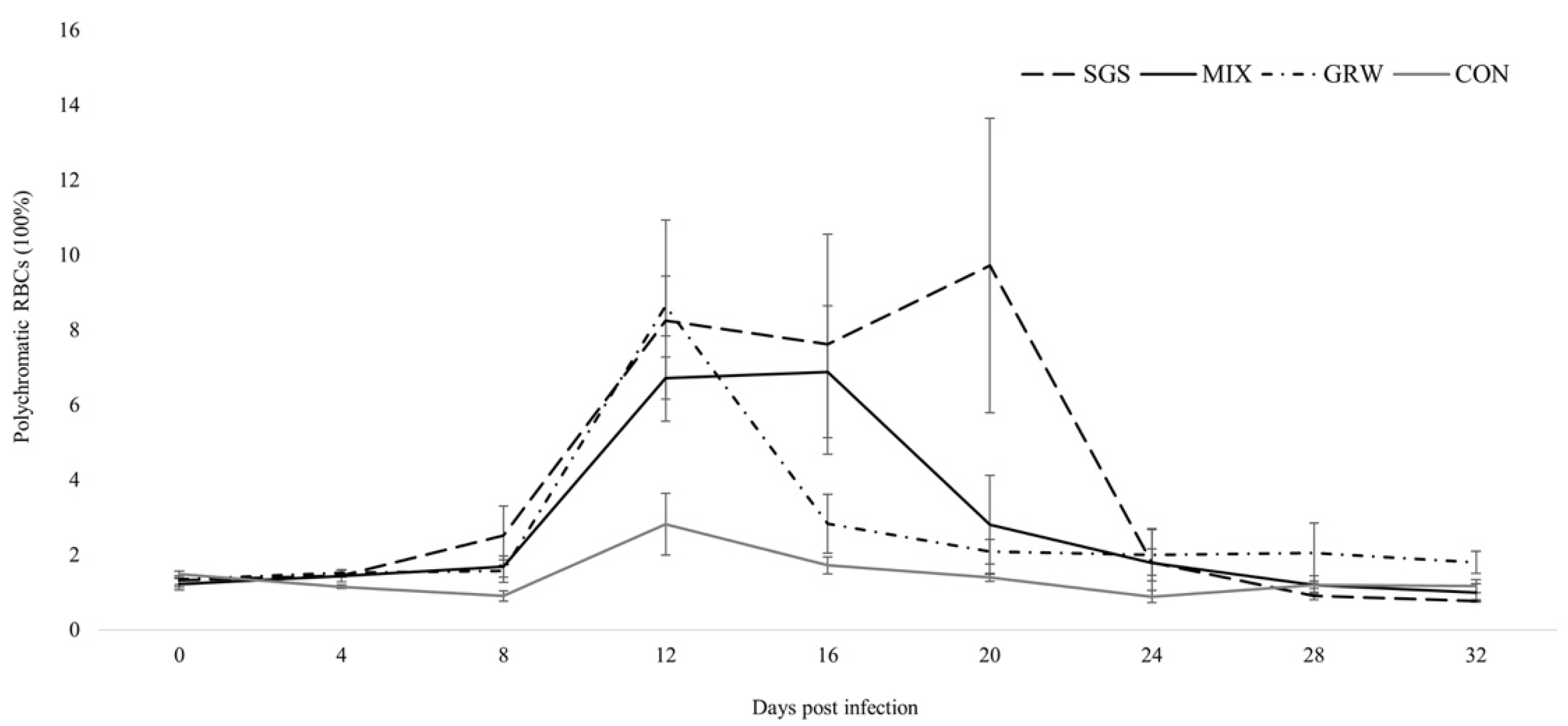
3.2. Changes of Health Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cox, F.E. Concomitant infections, parasites and immune responses. Parasitology 2001, 122, S23–S38. Available online: http://www.ncbi.nlm.nih.gov/pubmed/11442193 (accessed on 1 April 2022). [CrossRef] [PubMed] [Green Version]
- Poulin, R. Evolutionary Ecology of Parasites, 2nd ed.; Princeton University Press: Princeton, NJ, USA, 2007; pp. 1–332. [Google Scholar] [CrossRef]
- Cox, F.E.G.; Wedderburn, N.; Salaman, M.H. The effect of Rowson-Parr Virus on the severity of malaria in mice. J. Gen. Microbiol. 1974, 85, 358–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehlotra, R.K.; Miller, S.M.; Lorry, K.; Kastens, W.; Zimmerman, P.A.; Kazura, J.W.; Bockarie, M.; Alpers, M.P. Random distribution of mixed species malaria infections in Papua New Guinea. Am. J. Trop. Med. Hyg. 2000, 62, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiūnas, G.; Iezhova, T.A.; Shapoval, A.P. High prevalence of blood parasites in hawfinch Coccothraustes coccothraustes. J. Nat. Hist. 2003, 37, 2647–2652. [Google Scholar] [CrossRef]
- Palinauskas, V.; Žiegytė, R.; Ilgūnas, M.; Iezhova, T.A.; Bernotienė, R.; Bolshakov, C.; Valkiūnas, G. Description of the first cryptic avian malaria parasite, Plasmodium homocircumflexum n. sp., with experimental data on its virulence and development in avian hosts and mosquitoes. Int. J. Parasitol. 2015, 45, 51–62. [Google Scholar] [CrossRef]
- McQueen, P.G.; McKenzie, F.E. Competition for red blood cells can enhance Plasmodium vivax parasitemia in mixed-species malaria infections. Am. J. Trop. Med. Hyg. 2006, 75, 112–125. [Google Scholar] [CrossRef] [Green Version]
- Ramiro, R.S.; Pollitt, L.C.; Mideo, N.; Reece, S. Facilitation through altered resource availability in a mixed-species rodent malaria infection. Ecol. Lett. 2016, 19, 1041–1050. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Bensch, S.; Iezhova, T.A.; Križanauskiene, A.; Bolshakov, C.V. Nested cytochrome B polymerase chain reaction diagnostics underestimate mixed infections of avian blood haemosporidian parasites : Microscopy is still essential. J. Parasitol. 2006, 92, 418–422. [Google Scholar] [CrossRef]
- Valkiunas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar] [CrossRef]
- La Puente, J.M.-D.; Santiago-Alarcon, D.; Palinauskas, V.; Bensch, S. Plasmodium relictum. Trends Parasitol. 2020, 37, 355–356. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; de Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database, 3rd ed.; Invasive Species Specialist Group: Auckland, New Zealand, 2000. [Google Scholar]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Ilgūnas, M.; Palinauskas, V.; Iezhova, T.A.; Valkiūnas, G. Molecular and morphological characterization of two avian malaria parasites (Haemosporida: Plasmodiidae), with description of Plasmodium homonucleophilum n. sp. Zootaxa 2013, 3666, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Fragner, K.; Weissenböck, H.; Atkinson, C.T.; Iezhova, T.A. Characterization of Plasmodium relictum, a cosmopolitan agent of avian malaria. Malar. J. 2018, 17, 184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiūnas, G.; Zehtindjiev, P.; Hellgren, O.; Ilieva, M.; Iezhova, T.A.; Bensch, S. Linkage between mitochondrial cytochrome b lineages and morphospecies of two avian malaria parasites, with a description of Plasmodium (Novyella) ashfordi sp. nov. Parasitol. Res. 2007, 100, 1311–1322. [Google Scholar] [CrossRef] [PubMed]
- Palinauskas, V.; Kosarev, V.; Shapoval, A.P.; Bensch, S.; Valkiūnas, G. Comparison of mitochondrial cytochrome b lineages and morphospecies of two avian malaria parasites of the subgenus Haemamoeba and Giovannolaia (Haemosporida: Plasmodiidae). Zootaxa 2007, 50, 39–50. [Google Scholar] [CrossRef]
- Palinauskas, V.; Žiegytė, R.; Šengaut, J.; Bernotienė, R. Different paths—The same virulence: Experimental study on avian single and co-infections with Plasmodium relictum and Plasmodium elongatum. Int. J. Parasitol. 2018, 48, 1089–1096. [Google Scholar] [CrossRef]
- Palinauskas, V.; Valkiūnas, G.; Križanauskienė, A.; Bensch, S.; Bolshakov, C.V. Plasmodium relictum (lineage P-SGS1): Further observation of effects on experimentally infected passeriform birds, with remarks on treatment with Malarone™. Exp. Parasitol. 2009, 123, 134–139. [Google Scholar] [CrossRef]
- Ellis, V.A.; Cornet, S.; Merrill, L.; Kunkel, M.R.; Tsunekage, T.; Ricklefs, R.E. Host immune responses to experimental infection of Plasmodium relictum (lineage SGS1) in domestic canaries (Serinus canaria). Parasitol. Res. 2015, 114, 3627–3636. [Google Scholar] [CrossRef]
- Aželytė, J.; Platonova, E.; Bensch, S.; Hellgren, O.; Palinauskas, V. A comparative analysis of the dynamics of Plasmodium relictum (GRW4) development in the blood during single and co-infections. Acta Trop. 2021, 226, 106247. [Google Scholar] [CrossRef]
- Palinauskas, V.; Valkiūnas, G.; Bolshakov, C.V.; Bensch, S. Plasmodium relictum (lineage P-SGS1): Effects on experimentally infected passerine birds. Exp. Parasitol. 2008, 120, 372–380. [Google Scholar] [CrossRef]
- Palinauskas, V.; Valkiūnas, G.; Bolshakov, C.V.; Bensch, S. Plasmodium relictum (lineage SGS1) and Plasmodium ashfordi (lineage GRW2): The effects of the co-infection on experimentally infected passerine birds. Exp. Parasitol. 2011, 127, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Mukhin, A.; Palinauskas, V.; Platonova, E.; Kobylkov, D.; Vakoliuk, I.; Valkiūnas, G. The Strategy to Survive Primary Malaria Infection: An Experimental Study on Behavioural Changes in Parasitized Birds. PLoS ONE 2016, 11, e0159216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bichet, C.; Cornet, S.; Larcombe, S.; Sorci, G. Experimental inhibition of nitric oxide increases Plasmodium relictum (lineage SGS1) parasitaemia. Exp. Parasitol. 2012, 132, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cellier-Holzem, E.; Esparza-Salas, R.; Garnier, S.; Sorci, G. Effect of repeated exposure to Plasmodium relictum (lineage SGS1) on infection dynamics in domestic canaries. Int. J. Parasitol. 2010, 40, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- Larcombe, S.; Bichet, C.; Cornet, S.; Faivre, B.; Sorci, G. Food availability and competition do not modulate the costs of Plasmodium infection in dominant male canaries. Exp. Parasitol. 2013, 135, 708–714. [Google Scholar] [CrossRef] [Green Version]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Malaria infection increases bird attractiveness to uninfected mosquitoes. Ecol. Lett. 2012, 16, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Cornet, S.; Nicot, A.; Rivero, A.; Gandon, S. Evolution of Plastic Transmission Strategies in Avian Malaria. PLoS Pathog. 2014, 10, e1004308. [Google Scholar] [CrossRef] [Green Version]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55, 201–209. [Google Scholar] [CrossRef]
- Glaizot, O.; Fumagalli, L.; Iritano, K.; Lalubin, F.; Van Rooyen, J.; Christe, P. High Prevalence and Lineage Diversity of Avian Malaria in Wild Populations of Great Tits (Parus major) and Mosquitoes (Culex pipiens). PLoS ONE 2012, 7, e34964. [Google Scholar] [CrossRef]
- Marzal, A.; Ricklefs, R.E.; Valkiūnas, G.; Albayrak, T.; Arriero, E.; Bonneaud, C.; Czirják, G.; Ewen, J.; Hellgren, O.; Hořáková, D.; et al. Diversity, Loss, and Gain of Malaria Parasites in a Globally Invasive Bird. PLoS ONE 2011, 6, e21905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzal, A.; García-Longoria, L.; Callirgos, J.M.C.; Sehgal, R. Invasive avian malaria as an emerging parasitic disease in native birds of Peru. Biol. Invasions 2014, 17, 39–45. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Križanauskienė, A.; Palinauskas, V.; Sehgal, R.; Bensch, S. A Comparative Analysis of Microscopy and PCR-Based Detection Methods for Blood Parasites. J. Parasitol. 2008, 94, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, R.D.; Fedynich, A.M.; Pence, D.B. Quantification of hematozoa in blood smears. J. Wildl. Dis. 1987, 23, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Long Island, NY, USA, 1989. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Örjan, Ö.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. R. Soc. B Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, A. BioEdit: A user-friendly biological sequence alignment editor and analysis program of Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.; Fragner, K.; Platonova, E.; Weissenböck, H.; Valkiūnas, G. Patterns of Plasmodium homocircumflexum virulence in experimentally infected passerine birds. Malar. J. 2019, 18, 174. [Google Scholar] [CrossRef] [Green Version]
- Garnham, P.C.C. Malaria Parasites and Other Haemosporidia; Blackwell Scientific Publications Ltd.: Oxford, UK, 1966. [Google Scholar]
- Hellgren, O.; Kutzer, M.; Bensch, S.; Valkiūnas, G.; Palinauskas, V. Identification and characterization of the merozoite surface protein 1 (msp1) gene in a host-generalist avian malaria parasite, Plasmodium relictum (lineages SGS1 and GRW4) with the use of blood transcriptome. Malar. J. 2013, 12, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palinauskas, V.; Bernotienė, R.; Žiegytė, R.; Bensch, S.; Valkiūnas, G. Experimental evidence for hybridization of closely related lineages in Plasmodium relictum. Mol. Biochem. Parasitol. 2017, 217, 1–6. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palinauskas, V.; Žiegytė, R.; Šengaut, J.; Bernotienė, R. Experimental Study on Primary Bird Co-Infection with Two Plasmodium relictum Lineages—pSGS1 and pGRW11. Animals 2022, 12, 1879. https://doi.org/10.3390/ani12151879
Palinauskas V, Žiegytė R, Šengaut J, Bernotienė R. Experimental Study on Primary Bird Co-Infection with Two Plasmodium relictum Lineages—pSGS1 and pGRW11. Animals. 2022; 12(15):1879. https://doi.org/10.3390/ani12151879
Chicago/Turabian StylePalinauskas, Vaidas, Rita Žiegytė, Jakov Šengaut, and Rasa Bernotienė. 2022. "Experimental Study on Primary Bird Co-Infection with Two Plasmodium relictum Lineages—pSGS1 and pGRW11" Animals 12, no. 15: 1879. https://doi.org/10.3390/ani12151879