
Assessment of Mutation Drift Equilibrium and the Occurrence of a Recent Genetic Bottleneck in South Indian Zebu Cattle

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.; Huerta-Sanchez, E.; Casey, F.; Bradley, D.G. Cattle demographic history modeled from autosomal sequence variation. Philos. Trans. R. Soc. Lond B Biol. Sci. 2010, 365, 2531–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achilli, A.; Olivieri, A.; Pellecchia, M.; Uboldi, C.; Colli, L.; Al-Zahery, N.; Accetturo, M.; Pala, M.; Hooshiar Kashani, B.; Perego, U.A.; et al. Mitochondrial genomes of extinct aurochs survive in domestic cattle. Curr. Biol. 2008, 18, 157–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loftus, R.; MacHugh, D.; Bradley, D.; Sharp, P.; Cunningham, P. Evidence for two independent domestication of cattle. Proc. Natl. Acad. Sci. USA 1994, 91, 2757–2761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture; Scherf, B.D., Pilling, D., Eds.; FAO: Rome, Italy, 2015. [Google Scholar]
- Livestock Census: 20th Livestock Census of India-Key Results; Ministry of Fisheries, Animal Husbandry and Dairying, Department of Animal Husbandry and Dairying: Krishi Bhavan, India, 2019. Available online: https://epashuhaat.gov.in/documents/ProvisionalKeyResultsof20thLivestockCensus.pdf (accessed on 31 May 2022).
- ICAR-NBAGR. Guidelines for Management of Animal Genetic Resources of India; National Bureau of Animal Genetic Resources (Indian Council of Agricultural Research): Karnal, India, 2016; p. 163. [Google Scholar]
- Bradshaw, C.J.A.; Isagi, Y.; Kaneko, S.; Brook, B.W.; Bowman, D.M.J.S.; Frankham, R. Low genetic diversity in the bottlenecked population of endangered non-native banteng in northern Australia. Mol. Ecol. 2007, 16, 2998–3008. [Google Scholar] [CrossRef]
- Al-Atiyat, R.M. Extinction probabilities of Jordan indigenous cattle using population viability analysis. Livest. Sci. 2008, 123, 121–128. [Google Scholar] [CrossRef]
- Ganapathi, P.; Rajendran, R.; Kathiravan, P. Detection of occurrence of a recent genetic bottleneck event in Indian hill cattle breed Bargur using microsatellite markers. Trop. Anim. Health Prod. 2012, 44, 2007–2013. [Google Scholar] [CrossRef]
- Kataria, R.S.; Kathiravan, P.; Bulandi, S.S.; Pandey, D.; Mishra, B.P. Microsatellite based genetic monitoring to detect cryptic demographic bottleneck in Indian riverine buffaloes (Bubalus bubalis). Trop. Anim. Health Prod. 2010, 42, 849–855. [Google Scholar] [CrossRef]
- Glowatzki-Mullis, M.-L.; Muntwyler, J.; Bäumle, E.; Gaillard, C. Genetic diversity measures of Swiss goat breeds as decision-making support for conservation policy. Small Rumin. Res. 2008, 74, 202–211. [Google Scholar] [CrossRef]
- Amirinia, C.; Seyedabadi, H.; Banabazi, M.H.; Kamali, M.A. Bottleneck study and genetic structure of Iranian Caspian horse population using microsatellites. Pak. J. Biol. Sci. 2007, 10, 1540–1543. [Google Scholar] [CrossRef] [Green Version]
- Côté, S.D.; Dallas, J.F.; Marshall, F.; Irvine, R.J.; Langvatn, R.; Albon, S.D. Microsatellite DNA evidence for genetic drift and philopatry in Svalbard reindeer. Mol. Ecol. 2002, 11, 1923–1930. [Google Scholar] [CrossRef] [Green Version]
- Manomohan, V.; Saravanan, R.; Pichler, R.; Murali, N.; Sivakumar, K.; Sudhakar, K.; Raja, K.N.; Periasamy, K. Legacy of draught cattle breeds of South India: Insights into population structure, genetic admixture and maternal origin. PLoS ONE 2021, 16, e0246497. [Google Scholar] [CrossRef]
- FAO. Molecular Genetic Characterization of Animal Genetic Resources; FAO Animal Production and Health Guidelines. No. 9; FAO: Rome, Italy, 2011. [Google Scholar]
- Sambrook, J.; Russell, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbour Laboratory Press: Cold Spring Harbor, NY, USA, 2001. [Google Scholar]
- Grema, M.; Traoré, A.; Issa, M.; Hamani, M.; Abdou, M.; Soudré, A.; Sanou, M.; Pichler, R.; Tamboura, H.H.; Alhassane, Y.; et al. Short tandem repeat (STR) based genetic diversity and relationship of indigenous Niger cattle. Arch. Anim. Breed. 2017, 60, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, J.D.; Chakraborti, S. Nonparametric Statistical Inference; Marcel Dekker Inc.: New York, NY, USA, 1992. [Google Scholar]
- Piry, S.; Luikart, G.; Cornuet, J.M. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Luikart, G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics 1996, 144, 2001–2014. [Google Scholar] [CrossRef]
- Tantia, M.S.; Vijh, R.K.; Mishra, B.; Bharani Kumar, S.T.; Ahlawat, S.P.S. Evaluation of Indian fowl populations for mutation drift equilibrium. Indian J. Anim. Sci. 2006, 76, 937–943. [Google Scholar]
- Kathiravan, P.; Mishra, B.P.; Kataria, R.S.; Sadana, D.K. Evaluation of genetic architecture and mutation drift equilibrium of Marathwada buffalo population in Central India. Livest. Sci. 2009, 121, 288–293. [Google Scholar] [CrossRef]
- DADF. Estimated Livestock Population Breed-Wise Based on Breed Survey 2013; Department of Animal Husbandry, Dairying and Fisheries, Ministry of Agriculture, Government of India: New Delhi, India, 2015. [Google Scholar]
- Singh, P.K.; Sharma, A. Assessment of degree of endangerment of livestock breeds in India. Indian J. Anim. Sci. 2017, 87, 316–323. [Google Scholar]
- Di Rienzo, A.; Peterson, A.C.; Garza, J.C.; Valdes, A.-M.; Slatkin, M.; Freimer, N.B. Mutational processes of simple sequence repeat loci in human populations. Proc. Natl. Acad. Sci. USA 1994, 91, 3166–3170. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Chakraborty, R.; Fuerst, P.A. Infinite allele model with varying mutation rate. Proc. Natl. Acad. Sci. USA 1976, 73, 4164–4168. [Google Scholar] [CrossRef] [Green Version]
- Garza, J.C.; Williamson, E.G. Detection of reduction in population size using data from microsatellite loci. Mol. Ecol. 2001, 10, 305–318. [Google Scholar] [CrossRef]
- Williamson-Natesan, E.G. Comparison of methods for detecting bottlenecks from microsatellite loci. Conserv. Genet. 2005, 6, 551–562. [Google Scholar] [CrossRef]
- Girod, C.; Vitalis, R.; Lebois, R.; Freville, H. Inferring population decline and expansion from microsatellite data: A simulation-based evaluation of the MSvar methods. Genetics 2011, 188, 165–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peery, M.Z.; Kirby, R.; Reid, B.N.; Stoelting, R.; Doucet-Beer, E.; Robinson, S.; Vasquez-Carrillo, C.; Pauli, J.N.; Palsboll, P.J. Reliability of genetic bottleneck tests for detecting recent population declines. Mol. Ecol. 2012, 21, 3403–3418. [Google Scholar] [CrossRef] [PubMed]


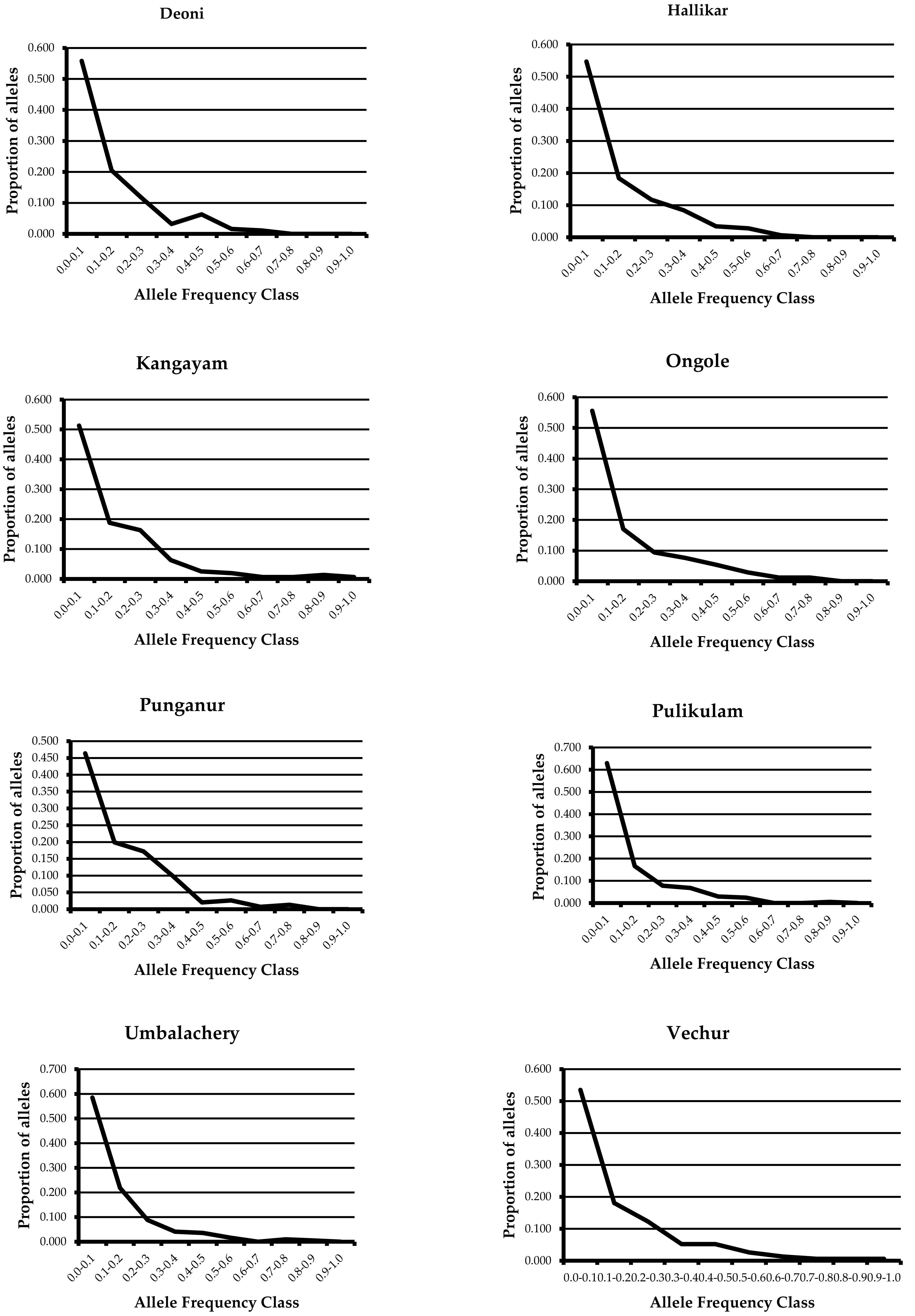{kind=link}
| Breed | n | ko | He | Heq | ||
|---|---|---|---|---|---|---|
| IAM | TPM | SMM | ||||
| Deoni | 47 | 7.46 | 0.721 | 0.689 | 0.741 | 0.784 |
| Hallikar | 36 | 7.00 | 0.728 | 0.695 | 0.740 | 0.778 |
| Kangayam | 50 | 6.25 | 0.662 | 0.619 | 0.676 | 0.725 |
| Ongole | 49 | 6.75 | 0.679 | 0.651 | 0.707 | 0.754 |
| Punganur | 18 | 6.04 | 0.741 | 0.740 | 0.766 | 0.788 |
| Pulikulam | 34 | 8.08 | 0.740 | 0.753 | 0.792 | 0.823 |
| Umbalachery | 33 | 7.58 | 0.719 | 0.730 | 0.770 | 0.801 |
| Vechur | 26 | 6.04 | 0.672 | 0.679 | 0.718 | 0.751 |
| Breed | Infinite Alleles Model (IAM) | Stepwise Mutation Model (SMM) | Two Phase Model (TPM) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of loci with | P-Value | No. of loci with | P-Value | No. of loci with | P-Value | |||||||
| He Deficit | He Excess | He Deficit | He Excess | He Deficit | He Excess | |||||||
| E | O | E | O | E | O | |||||||
| Deoni | 6 | 14.91 | 19 | 0.069 | 21 | 14.91 | 4 | 0.037 | 15 | 14.87 | 10 | 0.000 |
| Hallikar | 7 | 14.93 | 18 | 0.147 | 17 | 14.89 | 8 | 0.005 | 12 | 14.96 | 13 | 0.274 |
| Kangayam | 6 | 14.66 | 19 | 0.056 | 18 | 14.89 | 7 | 0.001 | 13 | 14.83 | 12 | 0.171 |
| Ongole | 8 | 14.79 | 17 | 0.245 | 20 | 14.85 | 5 | 0.000 | 14 | 14.87 | 11 | 0.086 |
| Punganur | 12 | 15.04 | 13 | 0.262 | 17 | 14.91 | 8 | 0.005 | 15 | 14.63 | 10 | 0.048 |
| Pulikulam | 15 | 14.96 | 10 | 0.036 | 20 | 14.78 | 5 | 0.000 | 17 | 15.02 | 8 | 0.004 |
| Umblachery | 10 | 15.02 | 15 | 0.572 | 15 | 14.91 | 10 | 0.037 | 13 | 15.05 | 12 | 0.149 |
| Vechur | 11 | 14.73 | 14 | 0.458 | 17 | 14.78 | 8 | 0.005 | 14 | 14.81 | 11 | 0.090 |
| Breed | Infinite Alleles Model (IAM) | Stepwise Mutation Model (SMM) | Two Phase Model (TPM) | |||
|---|---|---|---|---|---|---|
| T2 | P-Value | T2 | P-Value | T2 | P-Value | |
| Deoni | 1.042 | 0.149 | −8.147 | 0.000 | −2.418 | 0.008 |
| Hallikar | 1.270 | 0.102 | −5.911 | 0.000 | −1.483 | 0.069 |
| Kangayam | 1.830 | 0.034 | −5.038 | 0.000 | −0.776 | 0.219 |
| Ongole | 0.366 | 0.357 | −9.662 | 0.000 | −3.282 | 0.000 |
| Punganur | 0.344 | 0.365 | −4.375 | 0.000 | −1.605 | 0.054 |
| Pulikulam | −1.657 | 0.049 | −13.109 | 0.000 | −6.161 | 0.000 |
| Umbalachery | −0.889 | 0.187 | −9.881 | 0.000 | −4.537 | 0.000 |
| Vechur | −0.809 | 0.209 | −8.130 | 0.000 | −3.727 | 0.000 |
| Breed | Probability for One Tail Test (Heterozygosity Excess) | ||
|---|---|---|---|
| IAM | SMM | TPM | |
| Deoni | 0.015 | 1.000 | 0.831 |
| Hallikar | 0.100 | 0.998 | 0.729 |
| Kangayam | 0.024 | 0.996 | 0.625 |
| Ongole | 0.050 | 0.999 | 0.933 |
| Punganur | 0.213 | 0.998 | 0.909 |
| Pulikulam | 0.755 | 0.999 | 0.998 |
| Umbalachery | 0.604 | 0.993 | 0.943 |
| Vechur | 0.468 | 0.998 | 0.914 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manomohan, V.; Ramasamy, S.; Pichler, R.; Nagarajan, M.; Karuppusamy, S.; Krovvidi, S.; Nachiappan, R.K.; Peters, S.O.; Periasamy, K. Assessment of Mutation Drift Equilibrium and the Occurrence of a Recent Genetic Bottleneck in South Indian Zebu Cattle. Animals 2022, 12, 1838. https://doi.org/10.3390/ani12141838
Manomohan V, Ramasamy S, Pichler R, Nagarajan M, Karuppusamy S, Krovvidi S, Nachiappan RK, Peters SO, Periasamy K. Assessment of Mutation Drift Equilibrium and the Occurrence of a Recent Genetic Bottleneck in South Indian Zebu Cattle. Animals. 2022; 12(14):1838. https://doi.org/10.3390/ani12141838
Chicago/Turabian StyleManomohan, Vandana, Saravanan Ramasamy, Rudolf Pichler, Murali Nagarajan, Sivakumar Karuppusamy, Sudhakar Krovvidi, Raja K. Nachiappan, Sunday O. Peters, and Kathiravan Periasamy. 2022. "Assessment of Mutation Drift Equilibrium and the Occurrence of a Recent Genetic Bottleneck in South Indian Zebu Cattle" Animals 12, no. 14: 1838. https://doi.org/10.3390/ani12141838