
Canine Smell Preferences—Do Dogs Have Their Favorite Scents?

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animals
2.3. Odor Samples
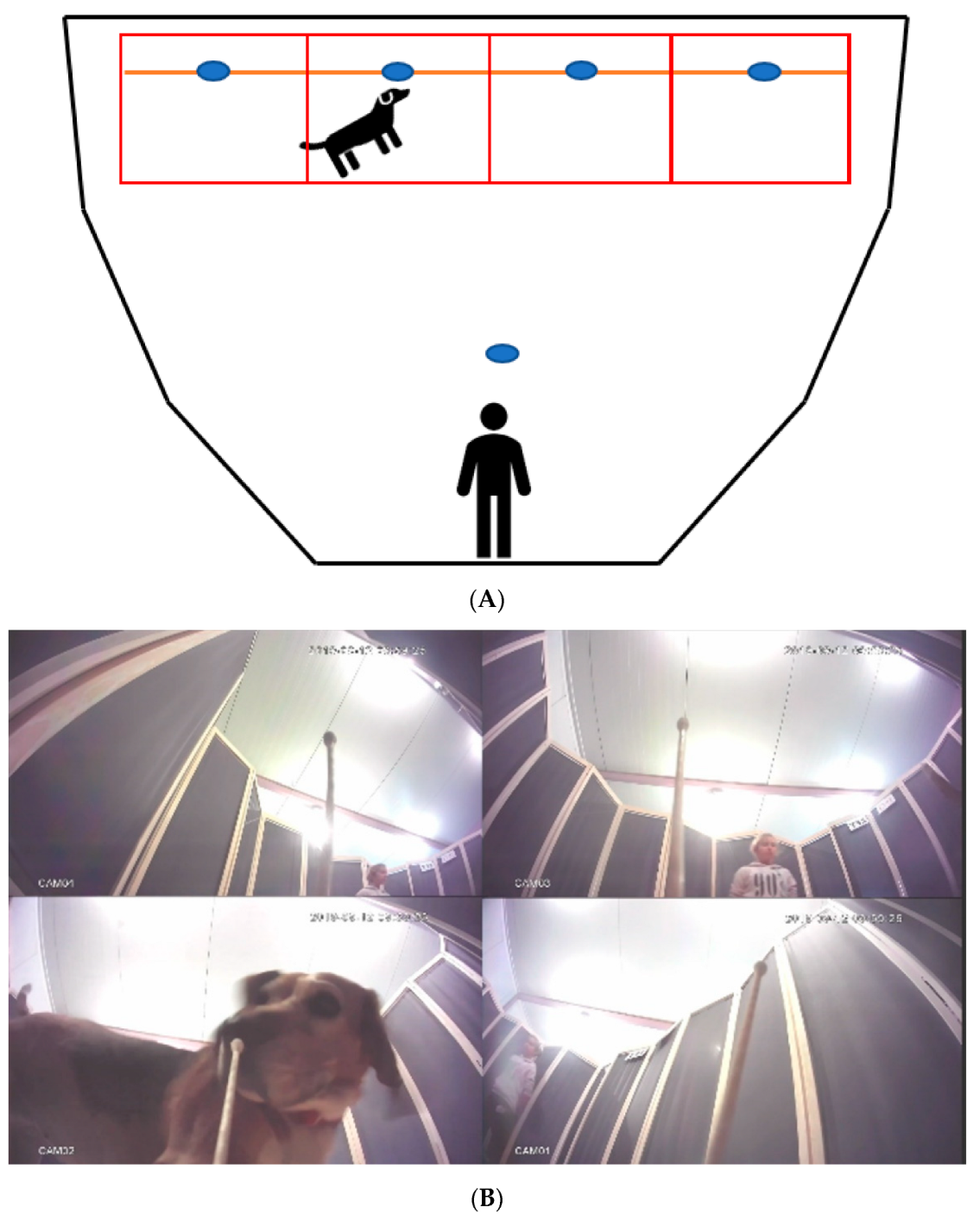
2.4. Experimental Design
2.5. Behavioral Analyses
2.6. Statistical Methods
3. Results
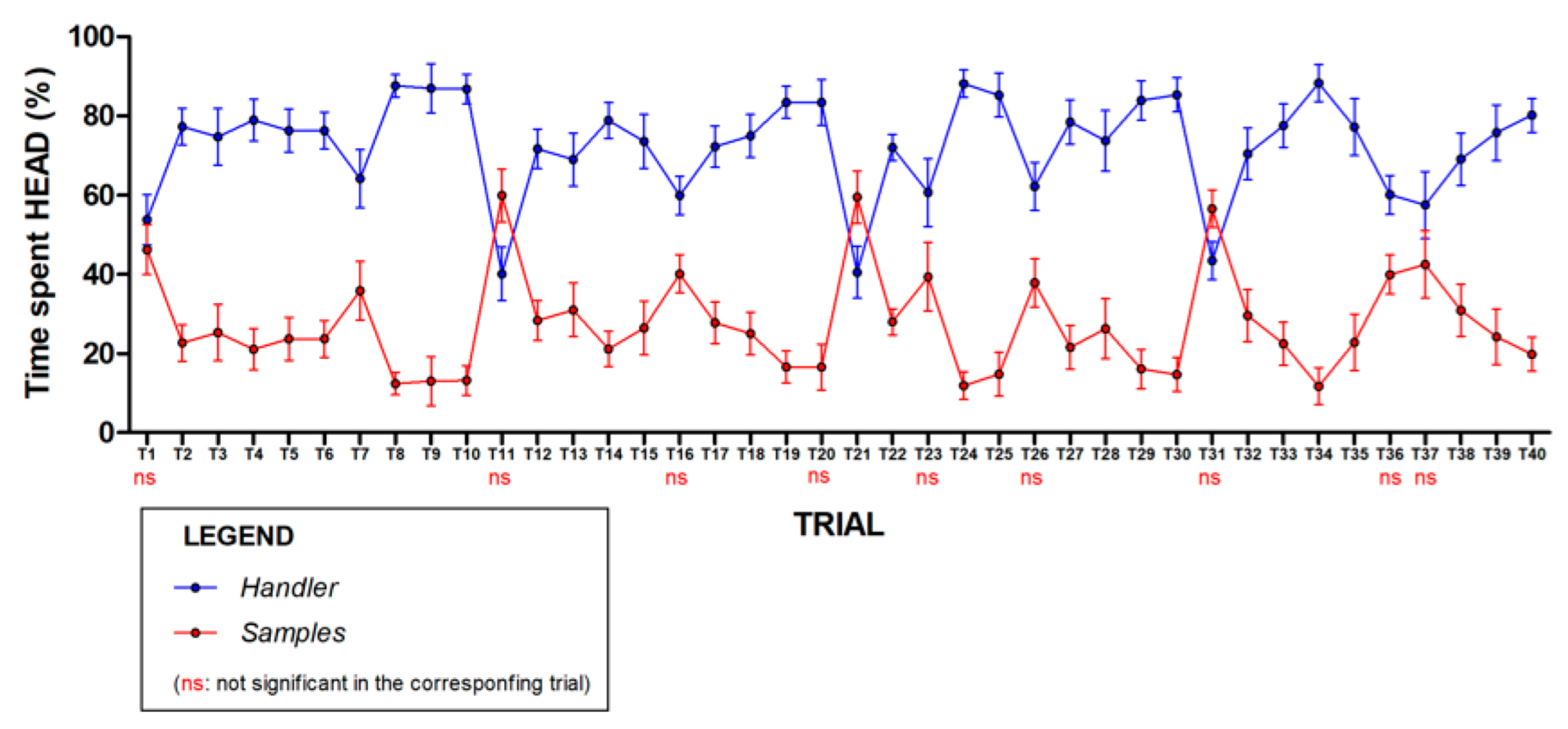
3.1. Analysis 1
3.2. Analysis 2
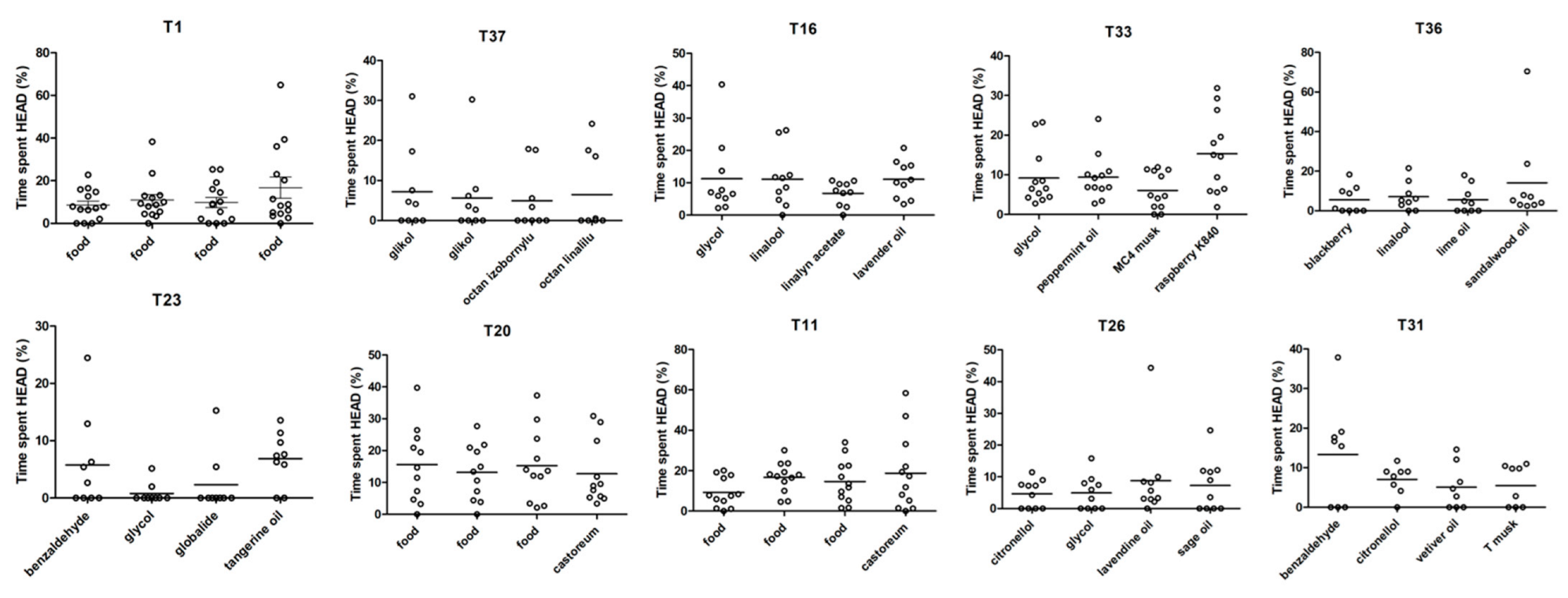
3.3. Analysis 3
3.4. Analysis 4
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jezierski, T.; Ensminger, J.; Papet, L.E. Canine Olfaction Science and Law: Advances in Forensic Science, Medicine, Conservation, and Environmental Remediation; CRC Press: Boca Raton, FL, USA; Taylor & Francis Group: Abingdon, UK, 2016. [Google Scholar]
- Dzięcioł, M.; Podgórski, P.; Stańczyk, E.; Szumny, A.; Woszczyło, M.; Pieczewska, B.; Niżański, W.; Nicpoń, J.; Wrzosek, M.A. MRI Features of the Vomeronasal Organ in Dogs (Canis familiaris). Front. Vet. Sci. 2020, 7, 159. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P. Morphological and immunohistochemical features of the vomeronasal system in dogs. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2013, 1, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Coli, A.; Stornelli, M.R.; Giannessi, E. The dog vomeronasal organ: A review. Dog Behav. 2016, 1, 24–31. [Google Scholar]
- Swaney, W.T.; Keverne, E.B. The evolution of pheromonal communication. Behav. Brain Res. 2009, 2, 239–247. [Google Scholar] [CrossRef]
- Stitzel, S.E.; Aernecke, M.J.; Walt, D.R. Artificial noses. Annu. Rev. Biomed. Eng. 2011, 13, 1–25. [Google Scholar] [CrossRef]
- Horowitz, A.; Hecht, J.; Dedrick, A. Smelling more or less: Investigating the olfactory experience of the domestic dog. Learn. Motiv. 2013, 44, 207–217. [Google Scholar] [CrossRef]
- Bogdan, S. Biologiczne Mechanizmy Zachowania się Ludzi i Zwierząt, 3rd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2007. [Google Scholar]
- Krzymowski, T.; Przała, J.; Dusza, L. Fizjologia Zwierząt Podręcznik dla Studentów Wydziałów Medycyny Weterynaryjnej, Wydziałów Biologii i Hodowli Zwierząt Akademii Rolniczych i Uniwersytetów, 8th ed.; PWRiL: Warszawa, Poland, 2005. [Google Scholar]
- Graham, L.; Wells, D.L.; Hepper, P.G. The influence of olfactory stimulation on the behaviour of dogs housed in a rescue shelter. Appl. Anim. Behav. Sci. 2005, 1, 143–153. [Google Scholar] [CrossRef]
- Carr, W.J.; Loeb, L.S.; Dissinger, M.L. Responses of rats to sex odors. J. Comp. Physiol. Psychol. 1965, 3, 370. [Google Scholar] [CrossRef]
- Freyberg, R.; Ahren, M.-P. A preliminary trial exploring perfume preferences in adolescent girls. J. Sens. Stud. 2011, 3, 237–243. [Google Scholar] [CrossRef]
- Hämmerli, A.; Schweisgut, C.; Kaegi, M. Population genetic segmentation of MHC-correlated perfume preferences. Int. J. Cosmet. Sci. 2012, 2, 161–168. [Google Scholar] [CrossRef]
- Havlicek, J.; Roberts, S.C.; Flegr, J. Women's preference for dominant male odour: Effects of menstrual cycle and relationship status. Biol. Lett. 2005, 3, 256–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granqvist, P.; Vestbrant, K.; Döllinger, L.; Liuzza, M.T.; Olsson, M.J.; Blomkvist, A.; Lundström, J.N. The scent of security: Odor of romantic partner alters subjective discomfort and autonomic stress responses in an adult attachment-dependent manner. Physiol. Behav. 2019, 198, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Groyecka, A.; Pisanski, K.; Sorokowska, A.; Havlíček, J.; Karwowski, M.; Puts, D.; Sorokowski, P. Attractiveness Is Multimodal: Beauty Is Also in the Nose and Ear of the Beholder. Front. Psychol. 2017, 8, 778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guéguen, N. Effect of a perfume on prosocial behavior of pedestrians. Psychol. Rep. 2001, 3, 1046–1048. [Google Scholar] [CrossRef] [PubMed]
- Havlicek, J.; Saxton, T.K.; Roberts, S.C.; Jozifkova, E.; Lhota, S.; Valentova, J.; Flegr, J. He sees, she smells? Male and female reports of sensory reliance in mate choice and non-mate choice contexts. Pers. Individ. Differ. 2008, 6, 565–570. [Google Scholar] [CrossRef]
- Herz, R.S.; Inzlicht, M. Sex differences in response to physical and social factors involved in human mate selection: The importance of smell for women. Evol. Hum. Behav. 2002, 5, 359–364. [Google Scholar] [CrossRef]
- Herz, R.S. Influences of Odors on Mood and Affective Cognition, in Olfaction, Taste, and Cognition; Cambridge University Press: New York, NY, USA, 2002; pp. 160–177. [Google Scholar]
- Allen, C.; Havlíček, J.; Williams, K.; Roberts, S.C. Evidence for odour-mediated assortative mating in humans: The impact of hormonal contraception and artificial fragrances. Physiol. Behav. 2019, 210, 112541. [Google Scholar] [CrossRef]
- Swift-Gallant, A.; Coome, L.; Srinivasan, S.; Monks, D.A. Non-neural androgen receptor promotes androphilic odor preference in mice. Horm. Behav. 2016, 83, 14–22. [Google Scholar] [CrossRef]
- Wei, Q.; Zhang, H.; Ma, S.; Guo, D. Sex- and age-related differences in c-fos expression in dog olfactory bulbs. Acta Zool. 2017, 4, 370–376. [Google Scholar] [CrossRef]
- Amdam, G.V.; Hovland, A.L. Measuring Animal Preferences and Choice Behavior. Nat. Educ. Knowl. 2011, 3, 74. [Google Scholar]
- Smotherman, W.P. Odor aversion learning by the rat fetus. Physiol. Behav. 1982, 5, 769–771. [Google Scholar] [CrossRef]
- Bilkó, A.; Altbäcker, V.; Hudson, R. Transmission of food preference in the rabbit: The means of information transfer. Physiol. Behav. 1994, 5, 907–912. [Google Scholar] [CrossRef]
- Arnould, C.; Schaal, B.; Orgeur, P. Olfactory Preferences in Newborn Lambs: Possible Influence of Prenatal Experience. BEH 1995, 5, 351–365. [Google Scholar] [CrossRef]
- Hepper, P.G.; Wells, D.L. Perinatal Olfactory Learning in the Domestic Dog. Chem. Senses 2006, 3, 207–212. [Google Scholar] [CrossRef] [Green Version]
- Shirasu, M.; Touhara, K. The scent of disease: Volatile organic compounds of the human body related to disease and disorder. J. Biochem. 2011, 3, 257–266. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Bronstad, P.M. Female body odour is a potential cue to ovulation. Proc. R. Soc. London. Ser. B Biol. Sci. 2001, 268, 797–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woszczyło, M.; Jezierski, T.; Szumny, A.; Niżański, W.; Dzięcioł, M. The role of urine in semiochemical communication between females and males of domestic dog (Canis familiaris) during estrus. Animals 2020, 11, 2112. [Google Scholar] [CrossRef]
- Sorokowska, A.; Sorokowski, P.; Szmajke, A. Does Personality Smell? Accuracy of Personality Assessments Based on Body Odour. Eur. J. Pers. 2012, 5, 496–503. [Google Scholar] [CrossRef]
- Sorokowska, A. Assessing Personality Using Body Odor: Differences between Children and Adults. J. Nonverbal Behav. 2013, 3, 153–163. [Google Scholar] [CrossRef] [Green Version]
- Sorokowska, A.; Sorokowski, P.; Havlíček, J. Body Odor Based Personality Judgments: The Effect of Fragranced Cosmetics. Front. Psychol. 2016, 7, 530. [Google Scholar] [CrossRef] [Green Version]
- Olsson, M.J.; Lundström, J.N.; Kimball, B.A.; Gordon, A.R.; Karshikoff, B.; Hosseini, N.; Lekander, M. The Scent of Disease: Human Body Odor Contains an Early Chemosensory Cue of Sickness. Psychol. Sci. 2014, 3, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Moshkin, M.; Litvinova, N.; Litvinova, E.A.; Bedareva, A.; Lutsyuk, A.; Gerlinskaya, L. Scent Recognition of Infected Status in Humans medicine. J. Sex. Med. 2011, 12, 3211–3218. [Google Scholar]
- Alasaad, S.; Permunian, R.; Gakuya, F.; Mutinda, M.; Soriguer, R.C.; Rossi, L. Sarcoptic-mange detector dogs used to identify infected animals during outbreaks in wildlife. BMC Vet. Res. 2012, 1, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, K.M.; Cotton, S.J.; Sandeman, R.M. The use of detector dogs in the diagnosis of nematode infections in sheep feces. J. Vet. Beh. 2008, 1, 25–31. [Google Scholar] [CrossRef]
- De Groot, J.H.B.; Smeets, M.A.; Rowson, M.J.; Bulsing, P.J.; Blonk, C.G.; Wilkinson, J.E.; Semin, G.R. A Sniff of Happiness. Psychol. Sci. 2015, 6, 684–700. [Google Scholar] [CrossRef]
- Fialová, J.; Roberts, S.C.; Havlíček, J. Is the Perception of Dietary Odour Cues Linked to Sexual Selection in Humans? In Chemical Signals in Vertebrates 12; Springer: New York, NY, USA, 2012; pp. 161–169. [Google Scholar]
- Havlicek, J.; Lenochova, P. The Effect of Meat Consumption on Body Odor Attractiveness. Chem. Senses 2006, 8, 747–752. [Google Scholar] [CrossRef]
- Zuniga, A.; Stevenson, R.J.; Mahmut, M.K.; Stephen, I.D. Diet quality and the attractiveness of male body odor. Evol. Hum. Behav. 2016, 1, 136–143. [Google Scholar] [CrossRef]
- Konno, A.; Romero, T.; Inoue-Murayama, M.; Saito, A.; Hasegawa, T. Dog Breed Differences in Visual Communication with Humans. PLoS ONE 2016, 11, e0164760. [Google Scholar] [CrossRef] [Green Version]
- Miklosi, A. Evolutionary approach to communication between humans and dogs. Vet. Res. Commun. 2009, 33 (Suppl. 1), 53–59. [Google Scholar] [CrossRef]
- Wasser, S.K.; Hunt, K.E.; Brown, J.L.; Cooper, K.; Crockett, C.M.; Bechert, U.; Monfort, S.L. A generalized fecal glucocorticoid assay for use in a diverse array of nondomestic mammalian and avian species. Gen. Comp. Endocrinol. 2000, 3, 260–275. [Google Scholar] [CrossRef]
- Hare, B.; Call, J.; Agnetta, B.; Tomasello, M. Chimpanzees know what conspecifics do and do not see. Anim. Behav. 2000, 4, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Call, J.; Bräuer, J.; Kaminski, J.; Tomasello, M. Domestic dogs (Canis familiaris) are sensitive to the attentional state of humans. J. Comp. Psychol. 2003, 3, 257–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horowitz, A. Disambiguating the “guilty look”: Salient prompts to a familiar dog behaviour. Behav. Processes 2009, 3, 447–452. [Google Scholar] [CrossRef] [PubMed]
- Schwab, C.; Huber, L. Obey or not obey? Dogs (Canis familiaris) behave differently in response to attentional states of their owners. J. Comp. Psychol. 2006, 3, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, H.J.; Kaminski, J.; Herrmann, E.; Tomasello, M. Understanding of human communicative motives in domestic dogs. Appl. Anim. Behav. Sci. 2011, 3, 235–245. [Google Scholar] [CrossRef] [Green Version]
- Soproni, K.; Miklósi, Á.; Topál, J.; Csányi, V. Dogs’ (Canis familaris) responsiveness to human pointing gestures. J. Comp. Psychol. 2002, 1, 27–34. [Google Scholar] [CrossRef]
- Szetei, V.; Miklósi, Á.; Topál, J.; Csányi, V. When dogs seem to lose their nose: An investigation on the use of visual and olfactory cues in communicative context between dog and owner. Appl. Anim. Behav. Sci. 2003, 2, 141–152. [Google Scholar] [CrossRef]
- Kokocińska-Kusiak, A.; Woszczyło, M.; Zybala, M.; Maciocha, J.; Barłowska, K.; Dzięcioł, M. Canine Olfaction: Physiology, Behavior, and Possibilities for Practical Applications. Animals 2021, 8, 2463. [Google Scholar] [CrossRef]
- Kirkden, R.D.; Pajor, E.A. Using preference, motivation and aversion tests to ask scientific questions about animals’ feelings. Appl. Anim. Behav. Sci. 2006, 1, 29–47. [Google Scholar] [CrossRef]
- Siniscalchi, M.; d’Ingeo, S.; Quaranta, A. Lateralized functions in the dog brain. Symmetry 2017, 5, 71. [Google Scholar] [CrossRef] [Green Version]
- Royet, J.P.; Plailly, J. Lateralization of olfactory processes. Chem. Senses 2004, 8, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Kaleta, T. Samoleczenie u dziko żyjących kręgowców–krótki przegląd zachowań Self-healing in the wild vertebrates—A brief behavioral overview. Życie Wet. 2005, 80, 278–282. [Google Scholar]
- Wells, D.L. Aromatherapy for travel-induced excitement in dogs. J. Am. Vet. Med. 2006, 6, 964–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komiya, M.; Sugiyama, A.; Tanabe, K.; Uchino, T.; Takeuchi, T. Evaluation of the effect of topical application of lavender oil on autonomic nerve activity in dogs. Am. J. Vet. Res. 2009, 6, 764–769. [Google Scholar] [CrossRef]
- Sayorwan, W.; Siripornpanich, V.; Piriyapunyaporn, T.; Hongratanaworakit, T.; Kotchabhakdi, N.; Ruangrungsi, N. The effects of lavender oil inhalation on emotional states, autonomic nervous system, and brain electrical activity. J. Med. Assoc. Thai. 2012, 4, 598–606. [Google Scholar]
- Atsumi, T.; Tonosaki, K. Smelling lavender and rosemary increases free radical scavenging activity and decreases cortisol level in saliva. Psychiatry Res. 2006, 1, 89–96. [Google Scholar] [CrossRef]
- Duranton, C.; Horowitz, A. Let me sniff! Nosework induces positive judgment bias in pet dogs. Appl. Anim. Behav. Sci. 2019, 211, 61–66. [Google Scholar] [CrossRef]
- Herz, R.S.; Eliassen, J.; Beland, S.; Souza, T. Neuroimaging evidence for the emotional potency of odor-evoked memory. Neuropsychologia 2004, 3, 371–378. [Google Scholar] [CrossRef]
- Herz, R.S. Odor-evoked memory. In The Oxford Handbook of Social Neuroscience; Decety, J., John, T., Eds.; Oxford University Press: Oxford, UK, 2011; pp. 265–275. [Google Scholar]
- Quaranta, A.; d’Ingeo, S.; Siniscalchi, M. Odour-Evoked Memory in Dogs: Do Odours Help to Retrieve Memories of Food Location? Animals 2020, 8, 1249. [Google Scholar] [CrossRef]
- Horowitz, A. Smelling themselves: Dogs investigate their own odours longer when modified in an “olfactory mirror” test. Behav. Processes 2017, 143, 17–24. [Google Scholar] [CrossRef]
- Uenoyama, R.; Miyazaki, T.; Hurst, J.L.; Beynon, R.J.; Adachi, M.; Murooka, T.; Miyazaki, M. The characteristic response of domestic cats to plant iridoids allows them to gain chemical defense against mosquitoes. Sci. Adv. 2021, 4, 9135. [Google Scholar] [CrossRef] [PubMed]
- Murtagh, K.; Farnworth, M.J.; Brilot, B.O. The scent of enrichment: Exploring the effect of odour and biological salience on behaviour during enrichment of kennelled dogs. Appl. Anim. Behav. Sci. 2020, 223, 104917. [Google Scholar] [CrossRef]
- Miyazaki, T.; Nakata, K.; Nishimura, T.; Abe, S.; Yamashita, T.; Miyazaki, M. Identification of 2-phenylethanol with a rose-like odor from anal sac secretions of the small Indian mongoose (Herpestes auropunctatus). Biosci. Biotechnol. Biochem. 2018, 2, 232–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhlemann, J.K.; Klempien, A.; Dudareva, N. Floral volatiles: From biosynthesis to function: Floral volatiles. Plant Cell Environ. 2014, 8, 1936–1949. [Google Scholar] [CrossRef]
- Shelbourne, T. The Truth about Wolves and Dogs; Veloce Publishing Ltd.: Dorchester, UK, 2012. [Google Scholar]
- Gable, T.D.; Windels, S.K.; Bruggink, J.G. Estimating biomass of berries consumed by gray wolves. Wildl. Soc. Bull. 2017, 1, 129–131. [Google Scholar] [CrossRef]
- Khrenov, Y.V. Ecology of a wolf. In Moлoдeжь XXI вeкa в нayчнoм, кyльmypнoм u oбpaзoвameльнoм npocmpaнcmвe: нoвыe цeннocmu, вызoвы, nepcneкmuвы; Poccийcкий yнивepcитeт дpyжбы нapoдoв (PУДН): Moscow, Russia, 2017; pp. 16–18. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dog | Breed | Age (Years) | Sex | Neutered |
|---|---|---|---|---|
| D1 | Polish Hunting Spaniel | 1 | F | + |
| D2 | Polish Hunting Spaniel | 1 | M | + |
| D3 | Polish Hunting Spaniel | 6 | F | + |
| D4 | Polish Hunting Spaniel | 3 | F | + |
| D5 | Mixed | 2 | M | + |
| D6 | Mixed | 8 | F | + |
| D7 | Bearded Collie | 9 | M | − |
| D8 | Tibetan Terrier | 14 | M | + |
| D9 | Shih Tzu | 10 | F | − |
| D10 | Polish Hunting Spaniel | 5 | F | − |
| D11 | Beagle | 8 | F | + |
| D12 | Beagle | 8 | F | + |
| D12 | Beagle | 9 | F | + |
| D14 | Beagle | 10 | F | + |
| Sample No. | Odor Sample |
|---|---|
| 1 | peppermint oil |
| 2 | blackberry K756 |
| 3 | isobornyl acetate |
| 4 | vetiver oil |
| 5 | linalool |
| 6 | citronellol |
| 7 | sage oil |
| 8 | musk MC4 |
| 9 | linalyl acetate |
| 10 | melon C186 |
| 11 | tangerine oil |
| 12 | benzaldehyde |
| 13 | globalide |
| 14 | rosewood oil |
| 15 | sandalwood oil |
| 16 | orange oil |
| 17 | rose oil |
| 18 | lime oil |
| 19 | beta pinene |
| 20 | lavandine oil |
| 21 | strawberry K814 |
| 22 | Eugenol |
| 23 | lavender oil |
| 24 | basil oil |
| 25 | raspberry K840 |
| 26 | ambrettolide |
| 27 | beta ionone |
| 28 | blueberry D761/G |
| 29 | geranium oil AT018 |
| 30 | T musk |
| 31 | glycol (negative control) |
| 32 | food—meat (positive control) |
| 33 | castoreum (positive control) |
| Dogs’ Positive Interaction | |||
|---|---|---|---|
| Predictors | Odds Ratios | CI | p |
| (Intercept) | 0.58 | 0.39–0.86 | 0.007 |
| Peppermint oil | 1.72 | 0.90–3.27 | 0.098 |
| Blackberry K756 | 0.91 | 0.47–1.77 | 0.779 |
| Isobornyl acetate | 1.00 | 0.52–1.93 | 1.000 |
| Vetiver oil | 0.62 | 0.31–1.26 | 0.188 |
| Linalool | 1.00 | 0.52–1.93 | 1.000 |
| Citronnellol | 1.00 | 0.52–1.93 | 1.000 |
| Sage oil | 1.44 | 0.76–2.74 | 0.267 |
| Musk MC4 | 0.80 | 0.41–1.56 | 0.511 |
| Linalyl acetate | 0.42 | 0.20–0.90 | 0.026 |
| Melon C186 | 0.82 | 0.39–1.76 | 0.620 |
| Tangerine oil | 0.89 | 0.49–1.64 | 0.712 |
| Benzaldehyde | 0.67 | 0.34–1.34 | 0.261 |
| Globalide | 0.54 | 0.26–1.11 | 0.092 |
| Rosewood oil | 0.91 | 0.47–1.77 | 0.779 |
| Sandalwood oil | 0.75 | 0.38–1.47 | 0.399 |
| Orange oil | 2.46 | 1.28–4.70 | 0.007 |
| Rose oil | 1.57 | 0.83–2.99 | 0.167 |
| Lime oil | 0.67 | 0.34–1.34 | 0.261 |
| Beta pinene | 0.82 | 0.42–1.62 | 0.575 |
| Lavandine oil | 0.75 | 0.38–1.47 | 0.399 |
| Strawberry K814 | 1.57 | 0.83–2.99 | 0.167 |
| Eugenol | 0.82 | 0.42–1.62 | 0.575 |
| Lavender oil | 1.10 | 0.57–2.11 | 0.780 |
| Basil oil | 0.91 | 0.47–1.77 | 0.779 |
| Raspberry K840 | 1.53 | 0.74–3.16 | 0.252 |
| Ambrettolide | 1.20 | 0.67–2.18 | 0.541 |
| Beta ionone | 0.91 | 0.47–1.77 | 0.779 |
| Blueberry D761/G | 0.94 | 0.44–1.98 | 0.869 |
| Geranium oil AT018 | 0.54 | 0.26–1.11 | 0.092 |
| T musk | 0.96 | 0.53–1.76 | 0.902 |
| Food-meat | 3.45 | 2.31–5.16 | <0.001 |
| Castoreum | 6.17 | 2.63–14.47 | <0.001 |
| Random Effects | |||
| σ2 | 3.29 | ||
| τ00 Dog | 0.26 | ||
| ICC | 0.07 | ||
| N Dog | 12 | ||
| Observations | 1920 | ||
| Marginal R2/Conditional R2 | 0.082/0.149 | ||
| Time | |||
|---|---|---|---|
| Predictors | Estimates | CI | p |
| (Intercept) | 3.05 | 2.42–3.69 | <0.001 |
| Peppermint oil | −0.42 | −1.51–0.67 | 0.452 |
| Blackberry K756 | 0.14 | −1.23–1.50 | 0.843 |
| Isobornyl acetate | 1.55 | 0.26–2.84 | 0.019 |
| Vetiver oil | 0.51 | −0.75–1.76 | 0.428 |
| Linalool | 0.59 | −0.74–1.92 | 0.385 |
| Citronnellol | 0.52 | −0.59–1.63 | 0.362 |
| Sage oil | 0.23 | −1.06–1.52 | 0.724 |
| Musk MC4 | 0.28 | −0.92–1.48 | 0.646 |
| Linalyl acetate | 1.60 | 0.34–2.85 | 0.013 |
| Melon C186 | 0.86 | −0.40–2.12 | 0.180 |
| Tangerine oil | 0.44 | −0.63–1.52 | 0.420 |
| Benzaldehyde | 0.41 | −1.05–1.87 | 0.585 |
| Globalide | 1.30 | 0.04–2.56 | 0.043 |
| Rosewood oil | 0.80 | −0.61–2.22 | 0.266 |
| Sandalwood oil | 0.81 | −0.66–2.28 | 0.278 |
| Orange oil | 1.22 | −0.01–2.45 | 0.052 |
| Rose oil | 0.69 | −0.44–1.82 | 0.230 |
| Lime oil | 1.91 | 1.26–2.56 | <0.001 |
| Beta pinene | 1.61 | 0.57–2.65 | 0.002 |
| Lavandine oil | 0.39 | −1.03–1.80 | 0.593 |
| Strawberry K814 | 0.72 | −0.50–1.95 | 0.249 |
| Eugenol | −0.40 | −1.62–0.83 | 0.526 |
| Lavender oil | 0.58 | −0.57–1.73 | 0.323 |
| Basil oil | 1.43 | 0.30–2.56 | 0.013 |
| Raspberry K840 | 0.49 | −0.80–1.78 | 0.460 |
| Ambrettolide | 2.88 | 1.29–4.47 | <0.001 |
| Beta ionone | 1.49 | 0.13–2.86 | 0.032 |
| Blueberry D761/G | −0.01 | −1.47–1.45 | 0.986 |
| Geranium oil AT018 | 0.84 | −0.42–2.10 | 0.189 |
| T musk | 0.86 | −0.47–2.18 | 0.205 |
| Food-meat | 0.17 | −0.86–1.19 | 0.750 |
| Castoreum | −0.31 | −1.42–0.80 | 0.581 |
| Random Effects | |||
| σ2 | 5.83 | ||
| τ00 Dog | 0.46 | ||
| ICC | 0.07 | ||
| N Dog | 12 | ||
| Observations | 775 | ||
| Marginal R2/Conditional R2 | 0.087/0.154 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kokocińska, A.; Woszczyło, M.; Sampino, S.; Dzięcioł, M.; Zybała, M.; Szczuka, A.; Korczyńska, J.; Rozempolska-Rucińska, I. Canine Smell Preferences—Do Dogs Have Their Favorite Scents? Animals 2022, 12, 1488. https://doi.org/10.3390/ani12121488
Kokocińska A, Woszczyło M, Sampino S, Dzięcioł M, Zybała M, Szczuka A, Korczyńska J, Rozempolska-Rucińska I. Canine Smell Preferences—Do Dogs Have Their Favorite Scents? Animals. 2022; 12(12):1488. https://doi.org/10.3390/ani12121488
Chicago/Turabian StyleKokocińska, Agata, Martyna Woszczyło, Silvestre Sampino, Michał Dzięcioł, Mikołaj Zybała, Anna Szczuka, Julita Korczyńska, and Iwona Rozempolska-Rucińska. 2022. "Canine Smell Preferences—Do Dogs Have Their Favorite Scents?" Animals 12, no. 12: 1488. https://doi.org/10.3390/ani12121488