1. Introduction
African male artiodactyl ungulates commonly investigate the urine of females during the breeding season. This gives males an opportunity to detect chemosensory cues in a female’s urine and then learn the sexual status of the female being investigated. The chemical cues in urine in the proestrus stage may announce that the female will come into full estrus soon, making it worthwhile for the male to follow the female. The typical behavior of a male antelope, such as a gazelle or impala, when pursuing females is to come into contact with freshly eliminated urine, often on the ground, and sample the urine in his oral cavity. Males may then perform the lip-curling flehmen response to analyze the estrous status of the female, which may indicate if she is in proestrus. If estrus is approaching, the male may then guard the female until mating is accomplished. If estrus is not imminent, the male will walk away.
In the case of giraffes, the general details of the giraffe bull’s investigation of the female and the performance of flehmen and, sometimes, the subsequent mating were described by Cynthia Moss [
1] in her classic book, “Portraits in the Wild”. This account drew upon the observations of Ann Innis/Ann Dagg [
2,
3], who continued by describing the bull gently sniffing, nudging, and/or kicking (laufschlag) the female, thereby inducing her to urinate [
4,
5]. Then, the male would collect the urine in his mouth, raise his head, and curl his lip in a flehmen response, sometimes then ejecting some urine from his mouth in a thin stream. Bercovitch and colleagues [
6] assessed how the reproductive behavior varies with the reproductive cycle of the female, documenting that males were more likely to associate with, and sexually investigate, females when they were cycling than when they were either pregnant or acyclic.
Like the vast majority of mammalian species, male giraffes are polygynous, compete for females, and invest little in their offspring. Giraffe males exhibit extreme sexual size dimorphism, and large-necked males gain favored access to estrous females [
7]. The male’s body weight is more than 50% greater than that of the female, and his head and neck continue growing, along with added bone; males have heightened risks of predation [
8]. They are challenged by the difficulty of balancing a cantilevered neck and head upon a relatively slight body [
9]. The mature male also becomes a very dark brown, providing an honest signal of strength and dominance: this is associated with greater mating success [
10]. These dark males may even have a musth period [
11].
Considering the problem of males who need to locate females that provide no prominent signals of when they are in estrus, research attention has focused on the vomeronasal organ (VNO). Moulton’s [
12] description in 1967 mentioned that in mammals, the VNO used in reproduction opens either anteriorly, into the nasal cavity, or into the buccal cavity by way of the nasopalatine canal, with the VNO sensory epithelium connecting to the accessory olfactory bulb by the vomeronasal nerve. Estes, writing in 1972 [
13], expanded our understanding of its role in mammalian reproduction and provided a table (adapted from Schilling 1970) charting the arrangements of the vomeronasal and incisive ducts in different groups of mammals. The table showed most artiodactyls, but not suids, as having openings of the vomeronasal ducts in both the mouth and nose. Giraffes were listed with a question mark, suggested as perhaps having a VNO communication with the nose only.
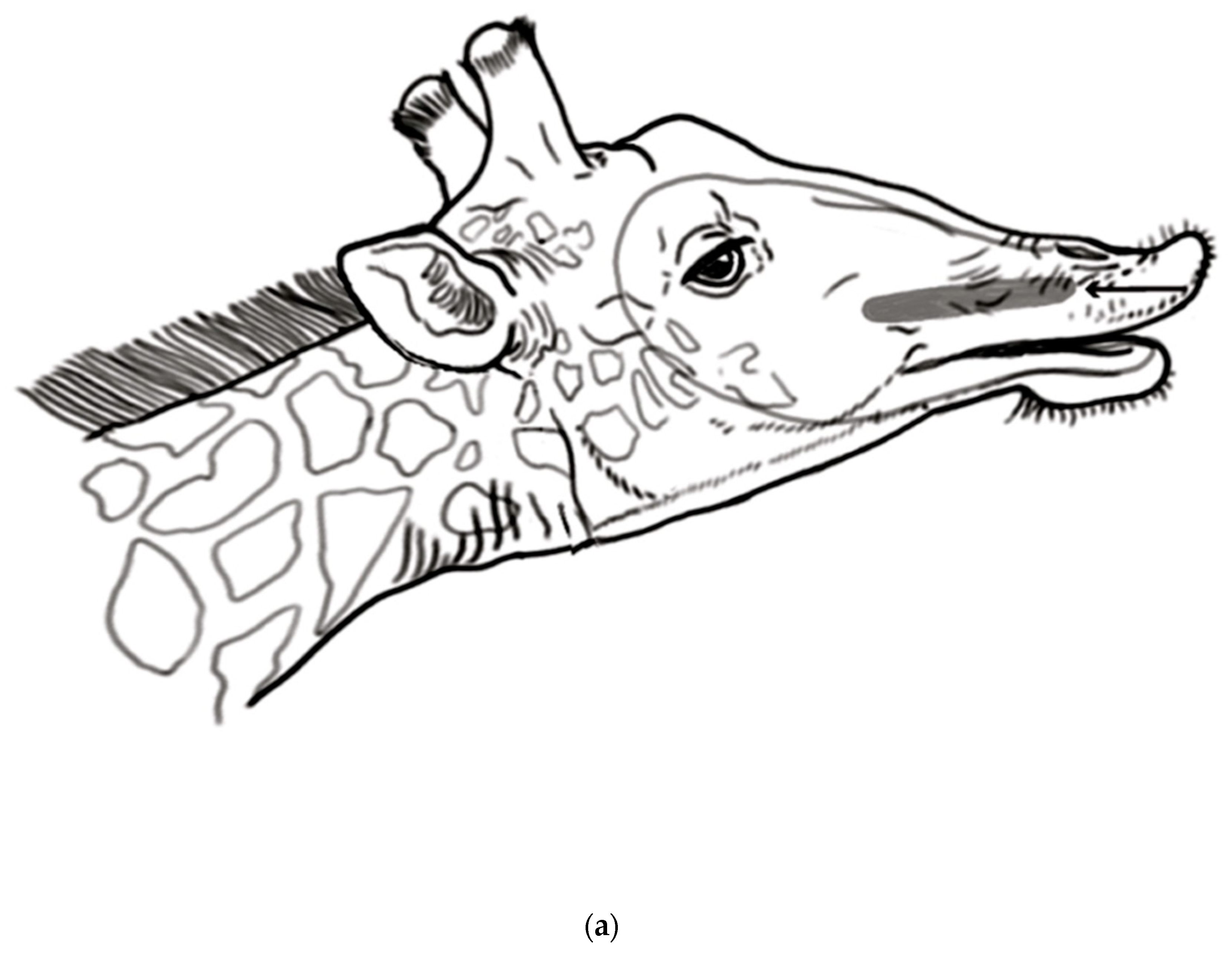
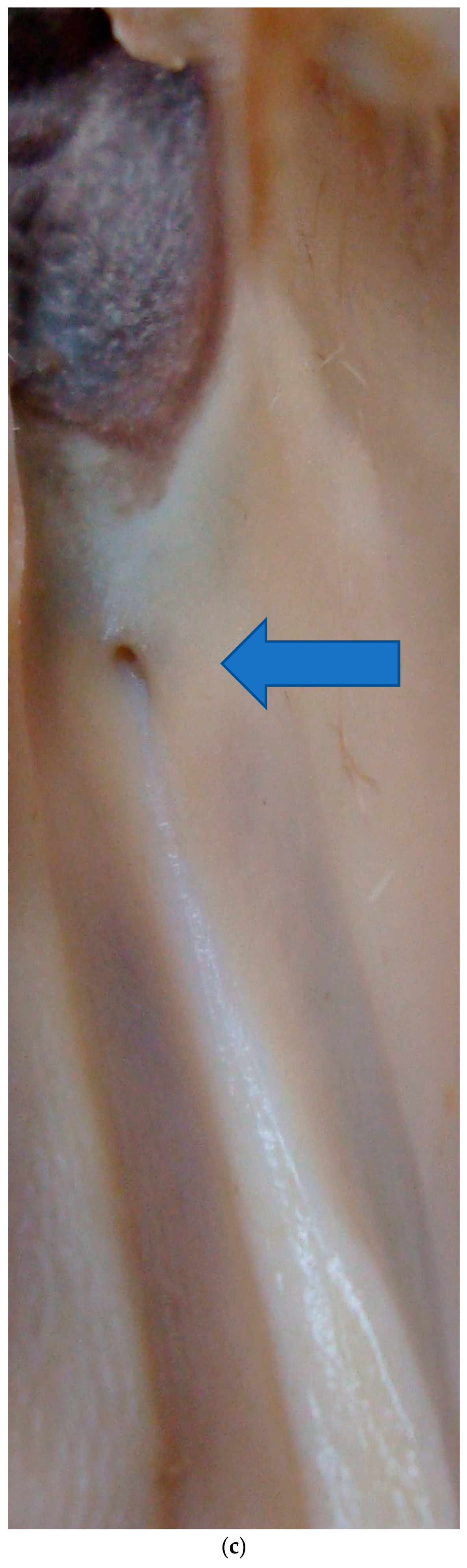
As is common in artiodactyls, a prominent funnel leads from the nasal cavity into the incisive duct that opens to the VNO and continues on to the incisive papilla. The flehmen response facilitates sucking material from the incisive papilla into the VNO. Bulls use their tongues to direct the urine being tested toward the VNO [
14] and dogs have also been shown to actively use their tongues [
15]. In giraffes, the tongue plays an active role in gathering urine from the urine stream; perhaps, male giraffes may also push the females’ urine into the incisive papillae.
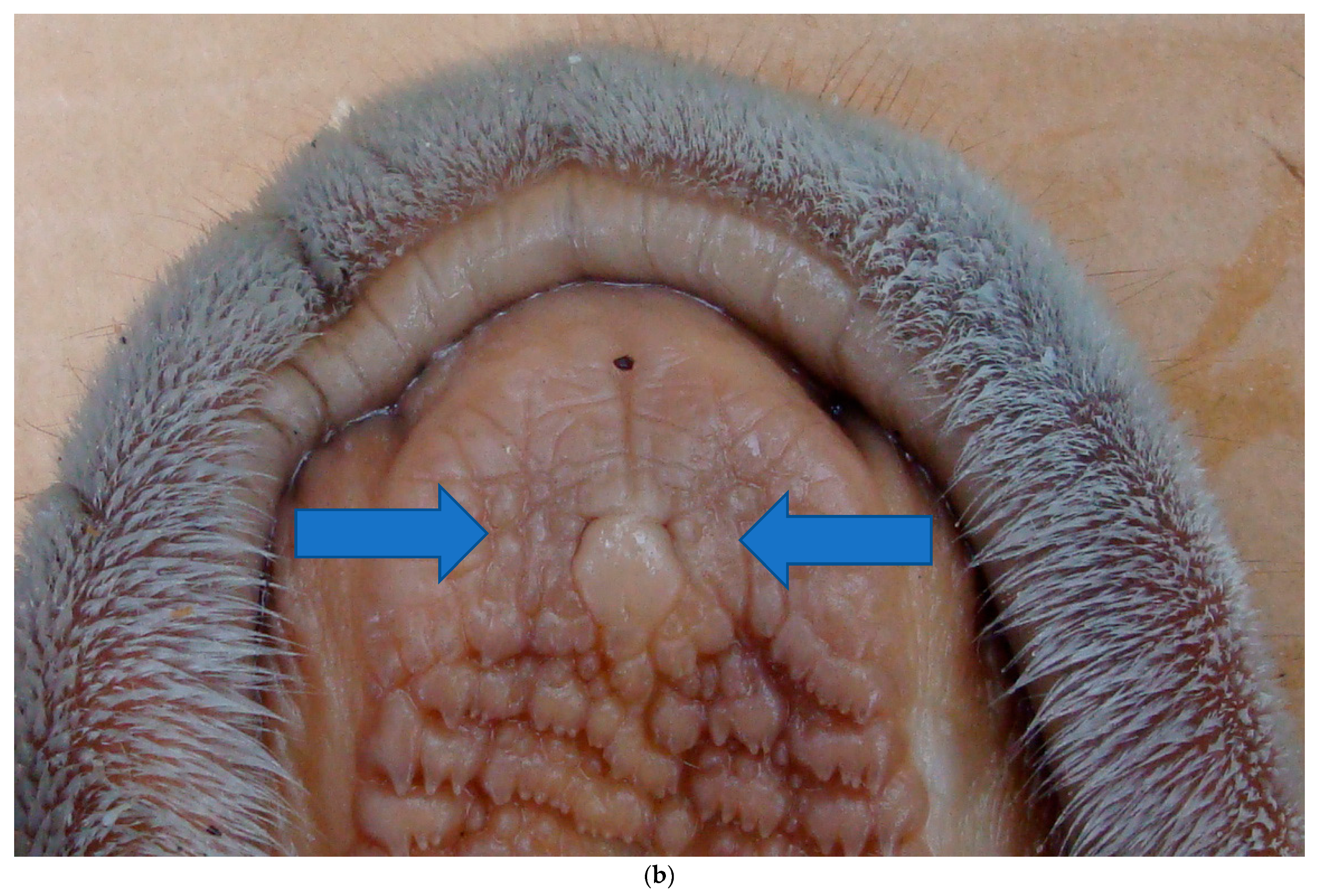
Exactly how the urine is assessed has been investigated somewhat in several African antelopes that have bilateral incisive papillae located on the hard palate, just behind the dental pad; each papilla connects to an incisive duct that leads to the bilateral tubular VNO. The incisive papillae and ducts are involved in transferring fluid-borne material from the oral cavity to the VNO, the bilateral accessory olfactory organs that are specialized for detecting the chemical cues related to proestrus and estrus; these urinary cues are sex pheromones. Flehmen behavior facilitates the transport of the collected urine from the oral cavity into the VNO in numerous artiodactyls. This widespread behavior is manifested in many mammals, including horses, cows, cats, and deer. As shown in a study on goats, the occurrence of flehmen stimulates emptying the VNO and pulling the new material into the VNO from the oral cavity through the incisive duct [
16]. The oral connection leads to the VNO, which is used by the male for the analysis of estrous pheromones in the female’s urine.
In giraffes, the vomeronasal organ includes sensory vomeronasal epithelia covering the medial region of the lumen and cartilage surrounds the soft tissue; veins likely play a role in pumping odorants into the lumen, and the giraffe have distinctive spongy small veins and capillaries that may serve as a secondary pump specific to giraffes [
17].
In most ungulates, there is a nasal connection to the VNO as well as an oral connection, and it has been proposed as a possibility that the nasal route to the VNO may be used for assessing certain plant-oriented chemicals related to detecting the season [
18]. The giraffe is the only ungulate species with a gestation period longer than a year. By breeding throughout the year, with some breeding peaks associated with rainfall, giraffes do not need to focus intently on a particular season.
The typical behavior of a male antelope, such as a gazelle or impala, when pursuing receptive females is to contact freshly eliminated urine, often on the ground, and sample the urine in his oral cavity [
19]. The eland usually acquires urine by intercepting the stream as the female urinates, but they do sometimes sample urine on the ground. After acquiring the female’s urine, a male may then perform the lip-curling flehmen motion to analyze the estrous status of the female, to find if she is in proestrus. If estrus is approaching soon, the male may then guard the female until mating is accomplished. If estrus is not imminent, the male will walk away.
An interesting exception to this pattern was found in our previous work on certain alcelaphine antelopes, including topi, hartebeest, and wildebeest, which lack an incisive papilla and oral connection to the VNO [
20]. Topi and hartebeest were also found to lack interest in female urine and failed to perform flehmen. Thus, these antelopes manifest a different behavior and anatomy than the other antelopes studied.
With our observations at waterholes during the dry season, giraffes sometimes congregated in large numbers and exhibited unusual behaviors. Bone chewing (osteophagia) was a behavior observed in multiple animals for extended periods. A description by Hutson and colleagues describes the typical behavior: “The bone was partly hidden inside the giraffe’s mouth and partly projecting forward from one side of the mouth, while the giraffe’s head was held high, and the lower jaws moved laterally. Profuse saliva dripped from the animal’s lips” [
21] (p. 4140). Until now, this perhaps appeared to be an infrequent behavior; we sought to note how often this behavior was recorded during our field observations. Little is known about osteophagia in giraffes, but in crested porcupines, lactating females engage in this behavior, presumably to bolster their calcium and phosphorus levels [
22].
The attentive responses of giraffes to a giraffe carcass are not described in the research literature: this behavior is first reported here. The high-amplitude, pulsed warning vocalizations by a dark male are also unusual.
The major objective of this study was to observe flehmen behavior and investigate its relationship to VNO anatomy. Given that giraffes breed throughout the year, the older, larger males need to be testing the females efficiently to locate individuals coming into estrus; they need an unambiguous testing method for identifying females of interest. Testing urine on the ground would be risky for a bull giraffe with such a heavy head and long neck, and the urine could also be difficult to link with a specific female. Solely investigating the genitalia and urine directly from the female would be the safest strategy for a bull giraffe and is made more necessary by their heavy anatomy. With serendipitous opportunities, we are presenting here additional and more infrequent behaviors of giraffes.
4. Discussion
The physical structure of the giraffe, with its elongated neck and legs, obviously serves this species very well, allowing them foraging access to highly placed leaves in tall trees. Giraffes have acquired the behavior of splaying the front legs, allowing them access to water, despite being capable of doing without it for long peiods [
10]. The observations reported here, although limited, suggest behavioral and anatomical adaptations to their physical structure to enable the investigation of urinary pheromones that indicate impending estrus in females.
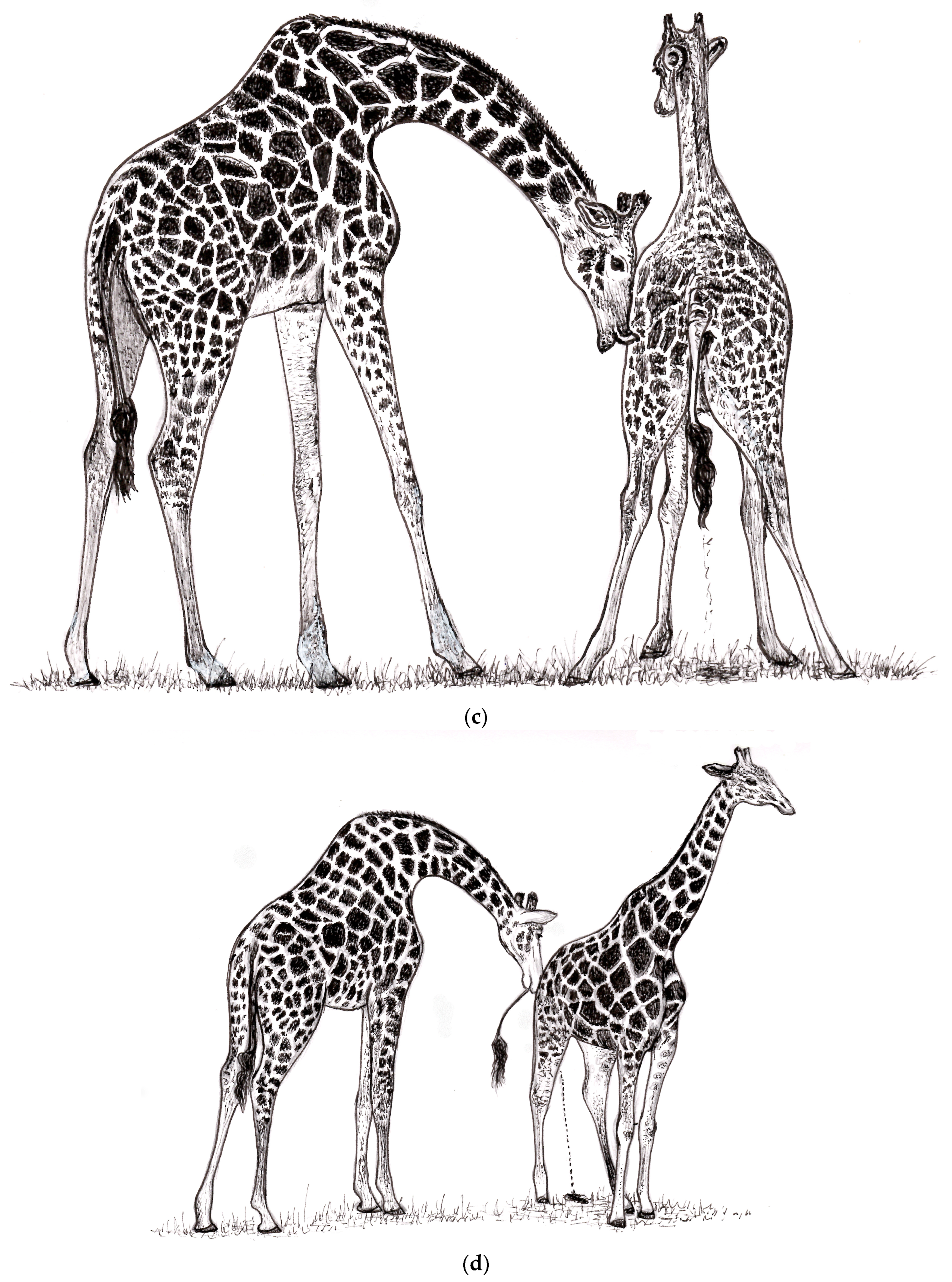
The behavioral adaptation is that a male needs to be vigilant to catch and test female urine while it is being expelled into his mouth for delivery into his VNO. Standing behind a female, perhaps nuzzling or tapping her rump, often triggers urination. Females that are near estrus will wait until a male is near to urinate.
Comparisons with antelopes, which we have studied previously, are interesting. Although oral connections to the VNO were missing or were extremely limited for alcelaphine antelopes, nasal connections were observed (resembling a funnel) between the ventro-rostral end of the nasal cavities and the VNO in all antelopes studied, comprising both non-alcelaphines and alcelaphines, including Thomson’s gazelle, Grant’s gazelle, impala, eland, black wildebeest, blue wildebeest, topi, Hunter’s hartebeest, blesbok, and Coke’s hartebeest [
18].
An adaptation in gross anatomy means that the access to the VNO for the giraffe is primarily through the bilateral incisive ducts, with a much more limited access through the nasal cavity. This VNO connectivity differs from antelopes, such as gazelles, impala, and eland, which have pronounced oral and nasal connections to the VNO [
18]. This proposed adaptation is consistent with a secondary function of the VNO in East African ungulates, which was previously proposed to be intended for detecting the vegetative olfactory cues that indicate season [
18]. Those other ungulates have reproductive cycles that are tuned to weather patterns and seasons each year. In contrast, giraffes have a longer gestation of 448 days [
23], which is not locked into a certain time of year.
For the female giraffe, consorting with the senior males that are so much larger and have large heads can impose some risks. She must stabilize herself, even at the early stage of courting, by widening her stance and bracing her legs. The fact that male giraffes never access urine from the ground resembles the behavior of the large antelope, the eland; for 90% of the time, the male tests the female’s readiness by either using the urine stream or the female’s genitalia [
19]. For such large animals as the eland or giraffe, reaching the ground necessitates great effort and is risky.
As emphasized by Brand [
10], the arid environment of Etosha National Park, especially in the dry seasons, combines with the waterholes to favor large dark male giraffes in terms of their mating success. In our study, these conditions meant that more giraffes were present for much of the time; a higher density of giraffes can also lead to more intense behaviors than are typical—as sometimes occurred in these observations. This helps explain the greater frequency of some of the more unusual behaviors described here. For example, although giraffes were long thought to be mute, they can emit loud warning calls that are sufficient to affect the behavior of other giraffes and cause them to flee. The procession of giraffes investigating the carcass of the dead giraffe is mysterious; were they exploring and expressing reverence for a familiar animal, or prospecting for bones to chew?
Their body size, use of space, and other unique characteristics of giraffes make them challenging to study and understand. This project touches on some additional uniqueness in terms of the coordinating reproductive behavior demonstrated between the male and female giraffes.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}










