
Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox


 and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
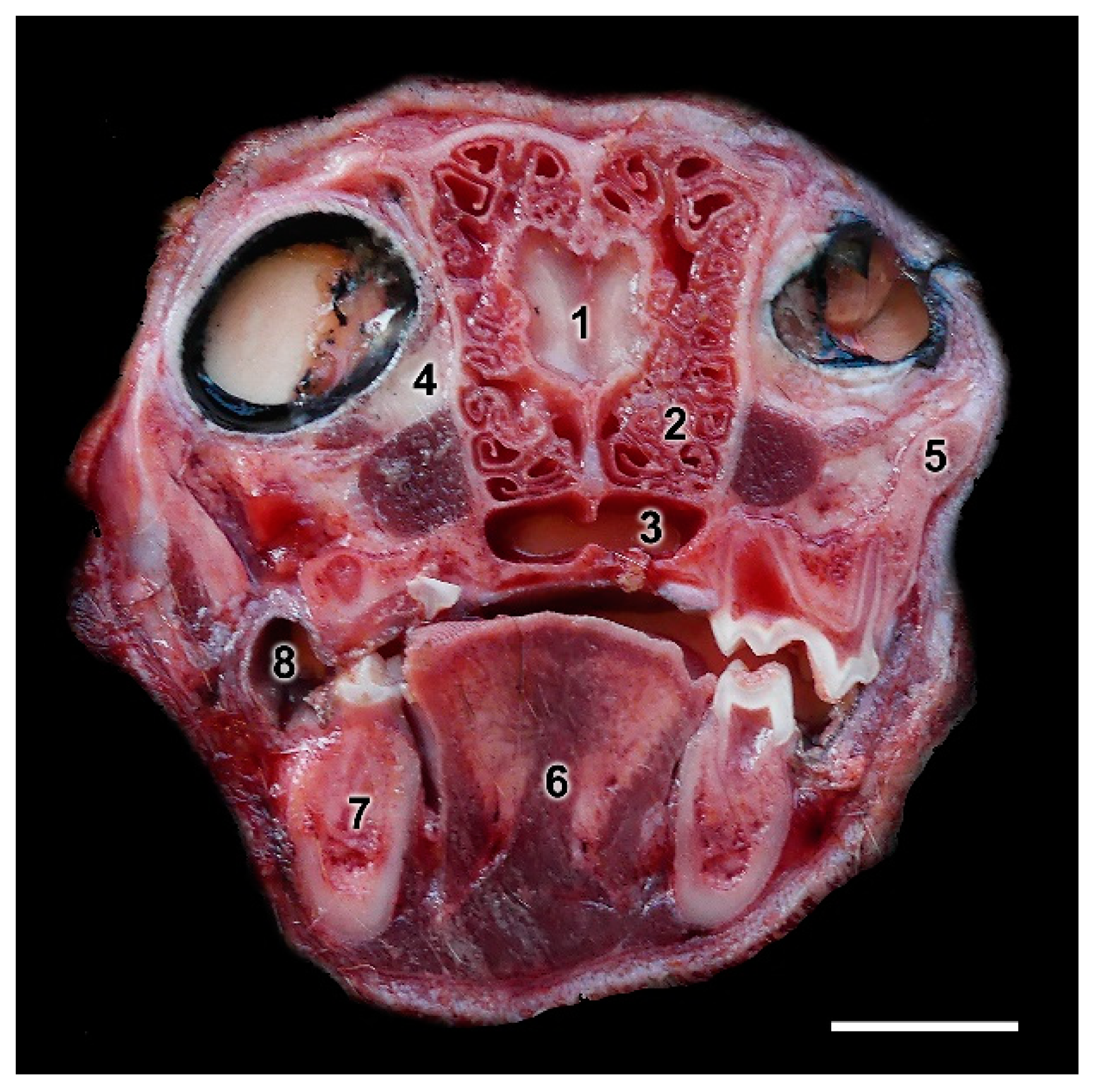
2.1. Samples
2.2. Tolivia Protocol
2.3. Gallego’s Trichrome Protocol
2.4. Immunohistochemical Protocol
2.5. Lectin Histochemistry Protocol
2.5.1. Lycopersicon Esculentum Agglutinin (LEA) Protocol
2.5.2. Ulex Europaeus Agglutinin (UEA) Protocol
2.6. Acquisition of Images
3. Results
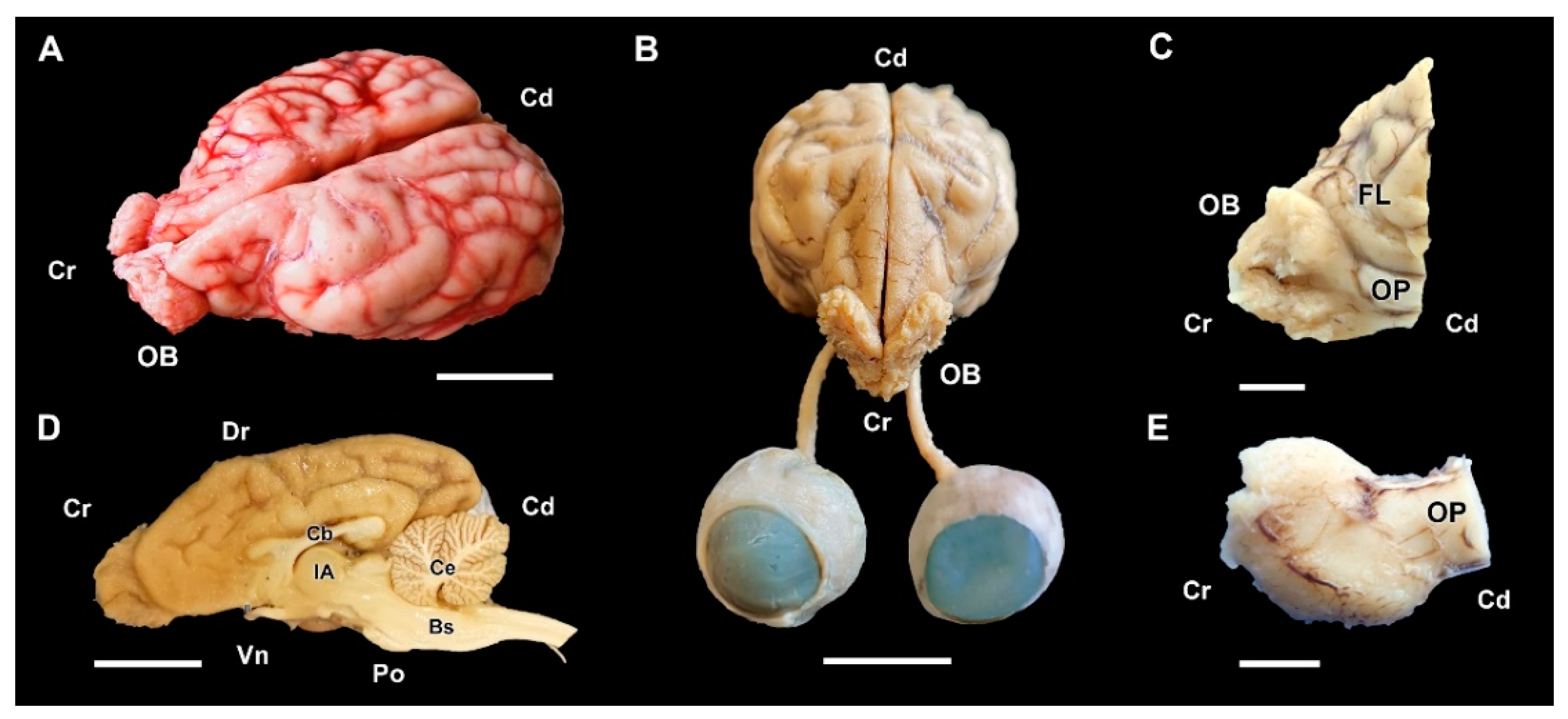
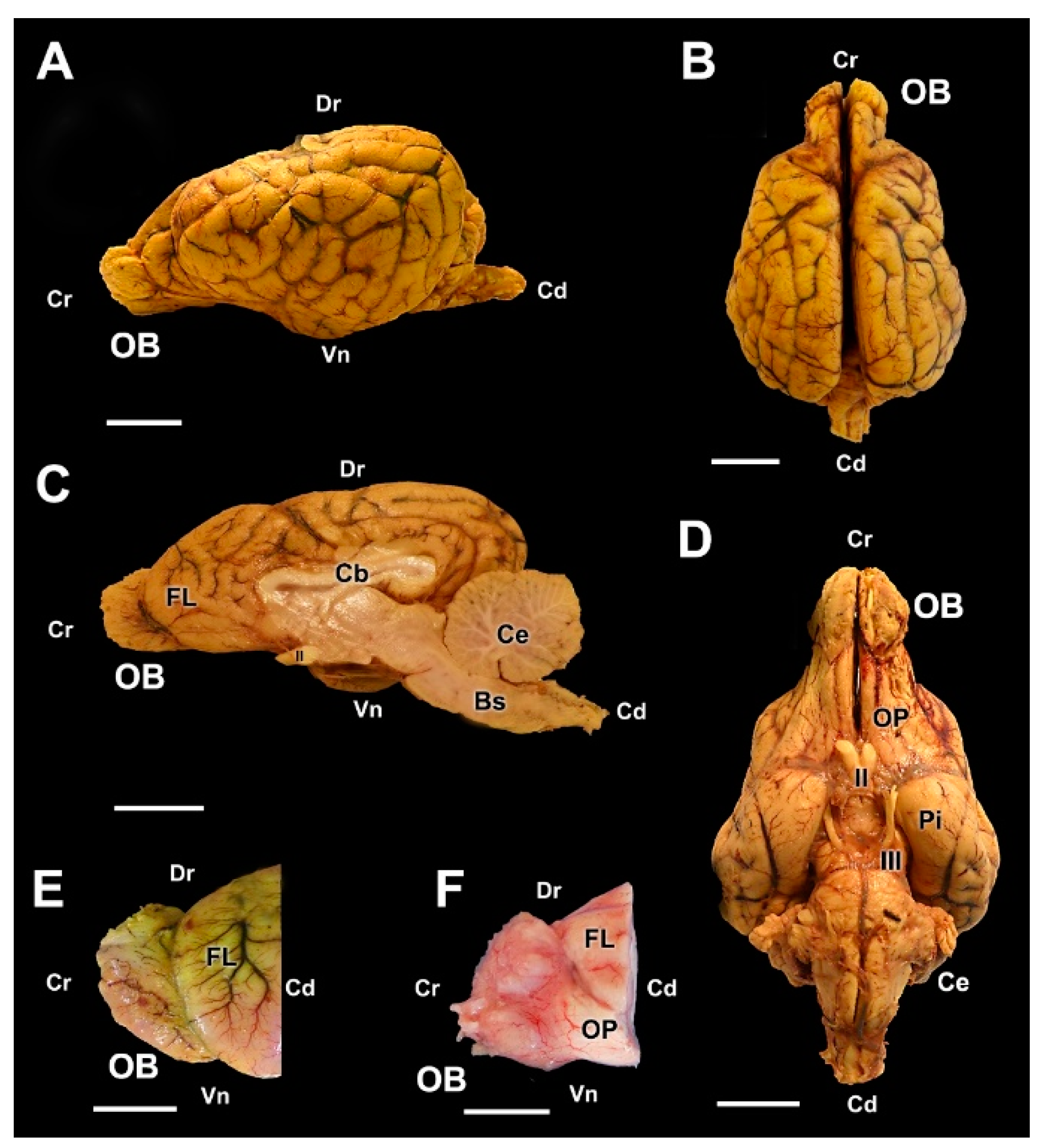
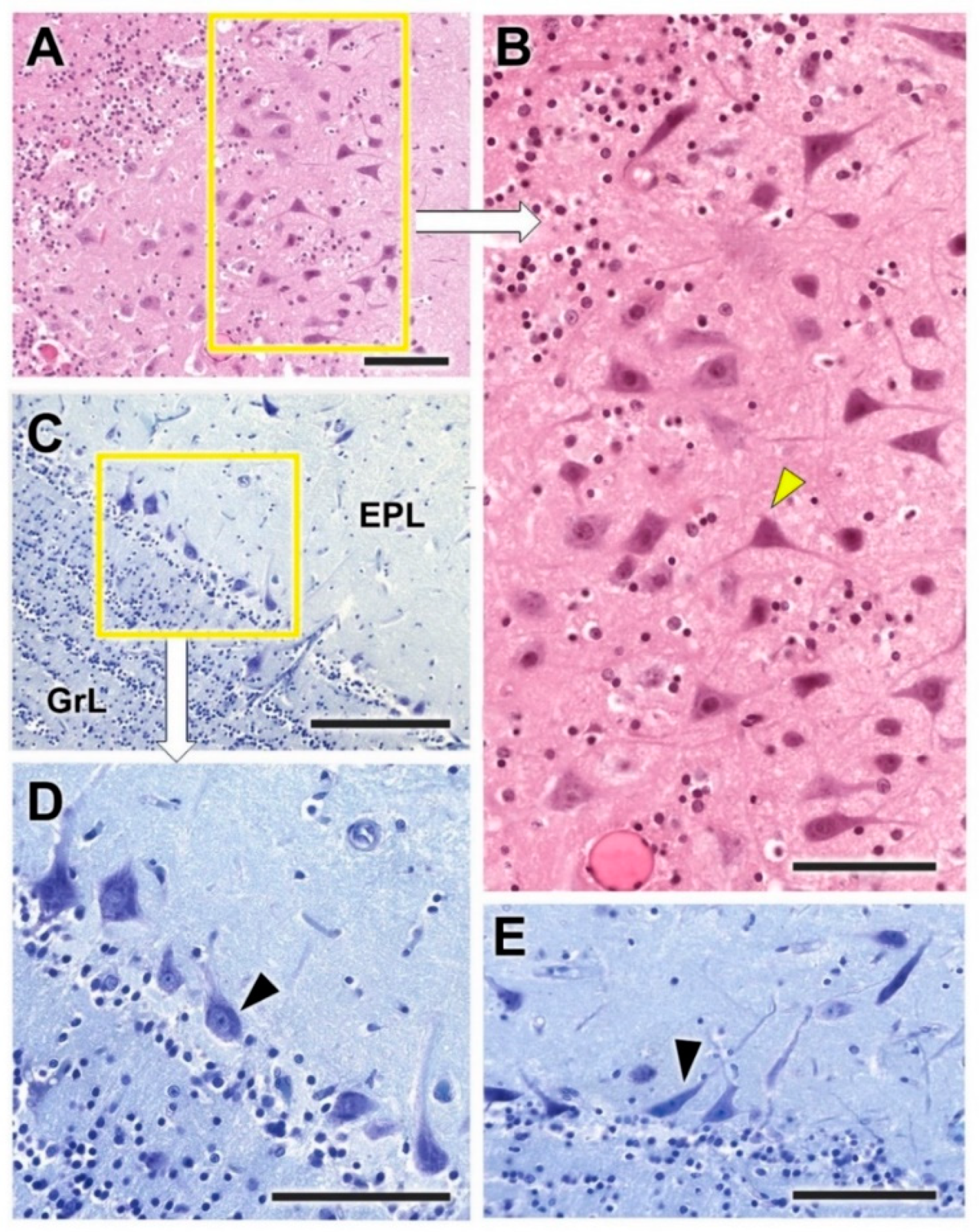
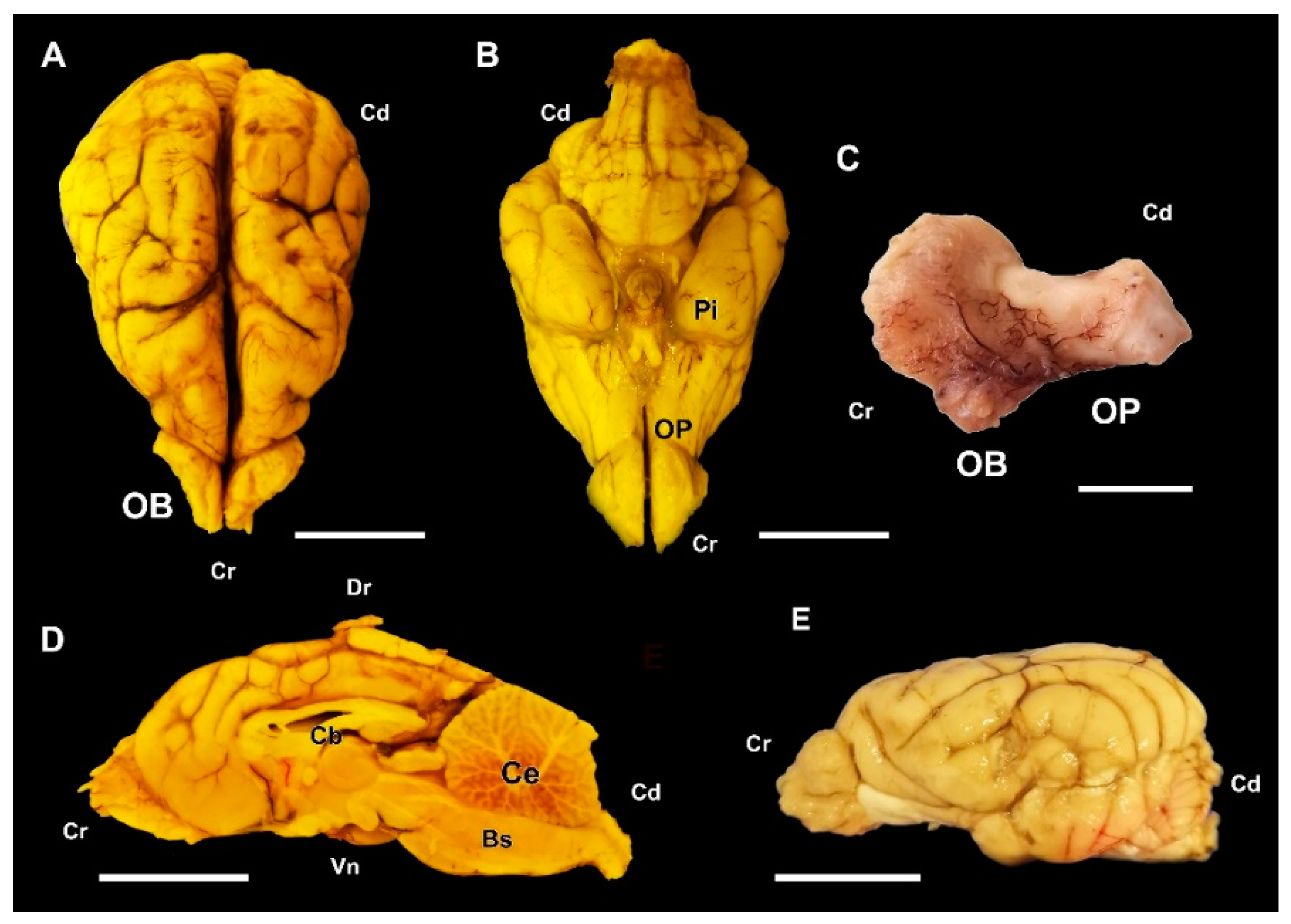
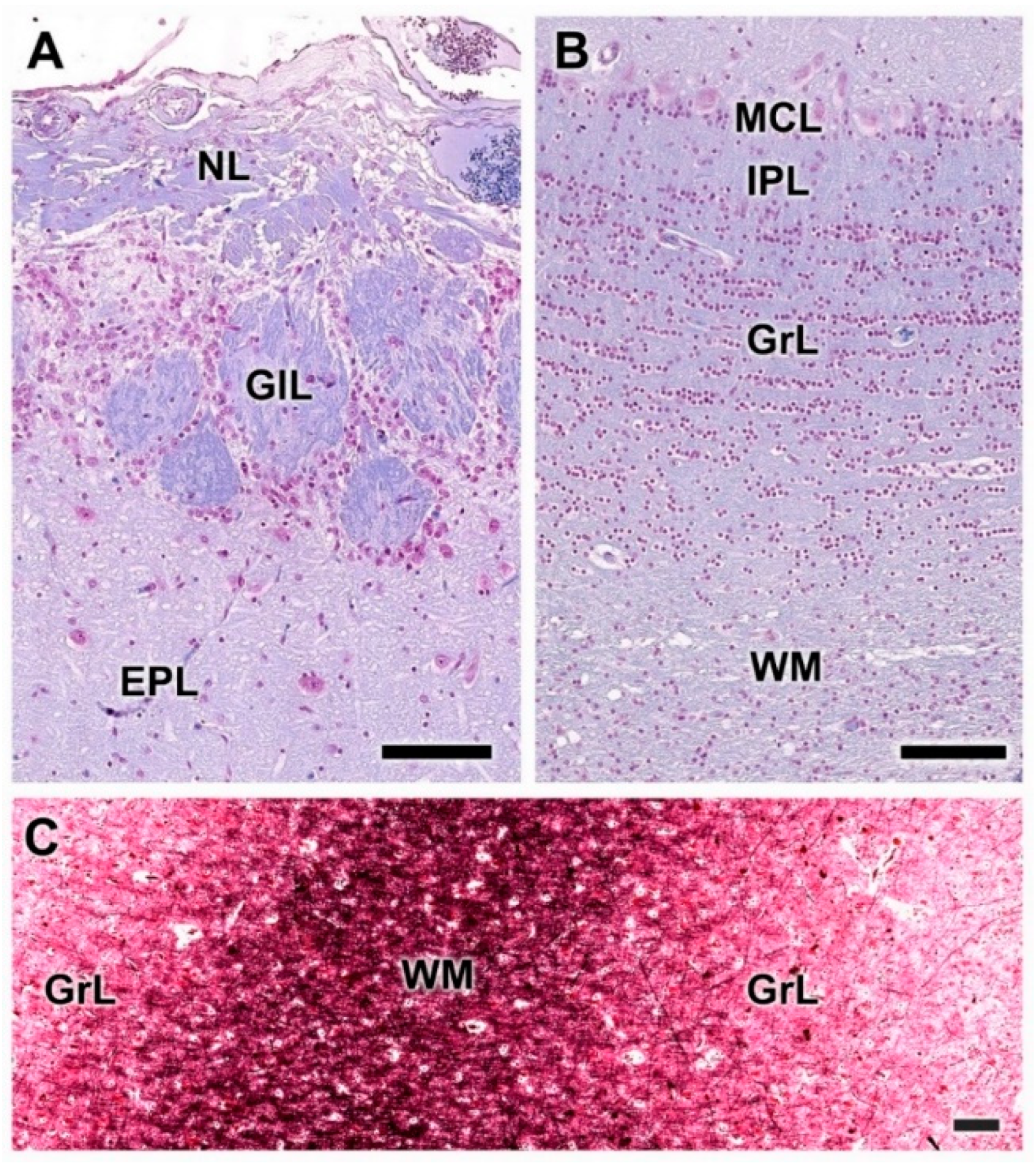
3.1. Dog Olfactory Bulb
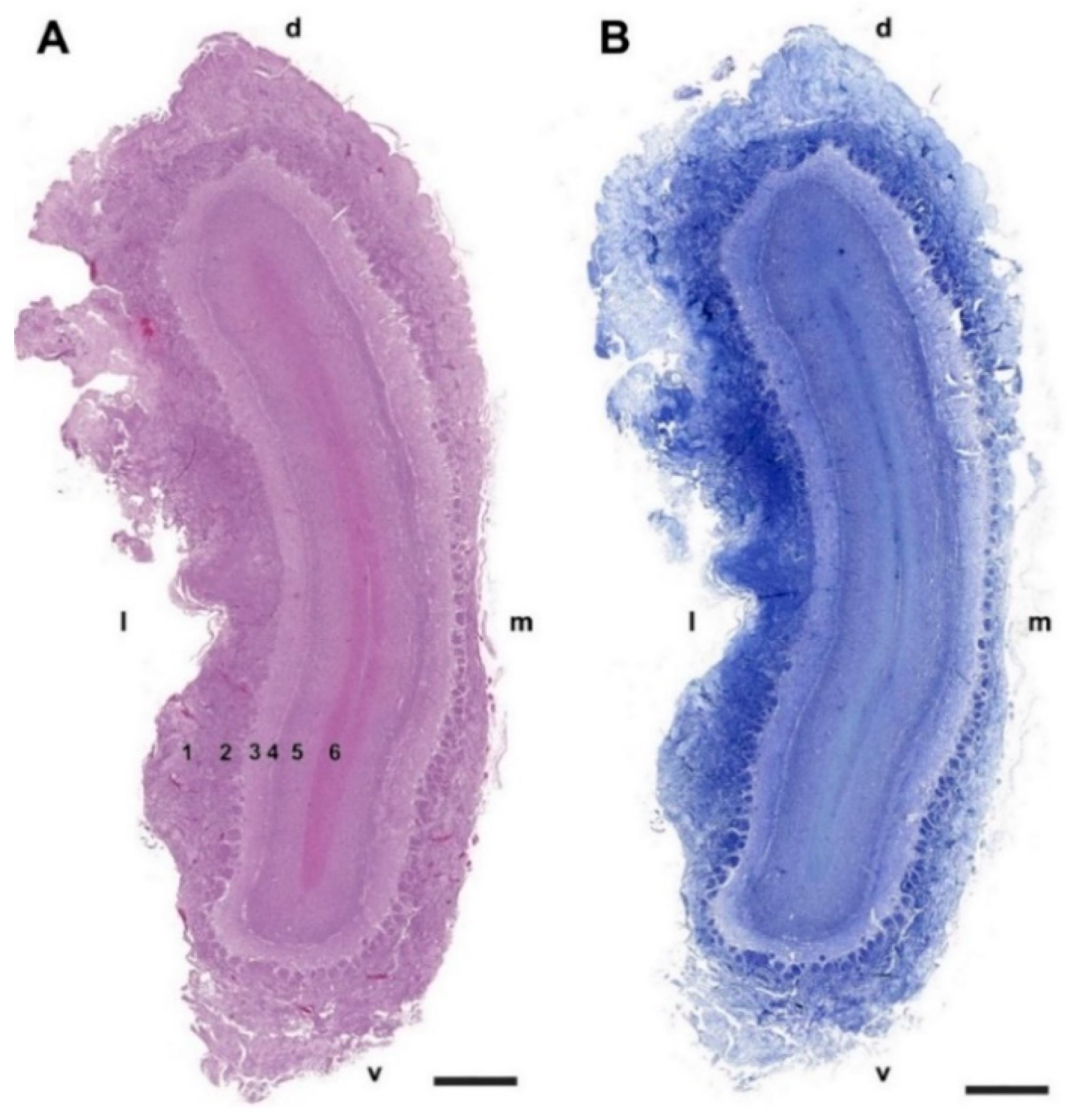
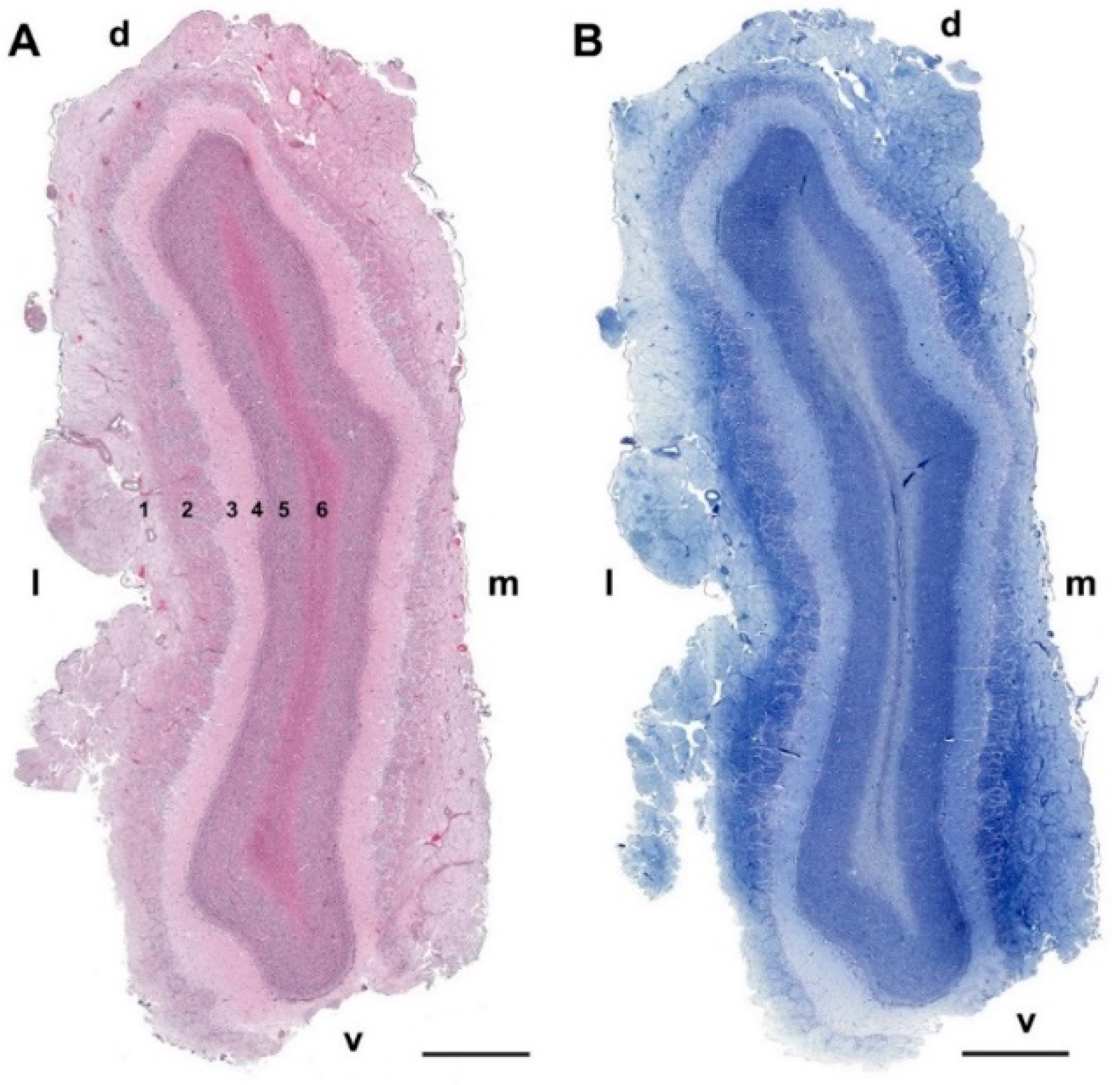
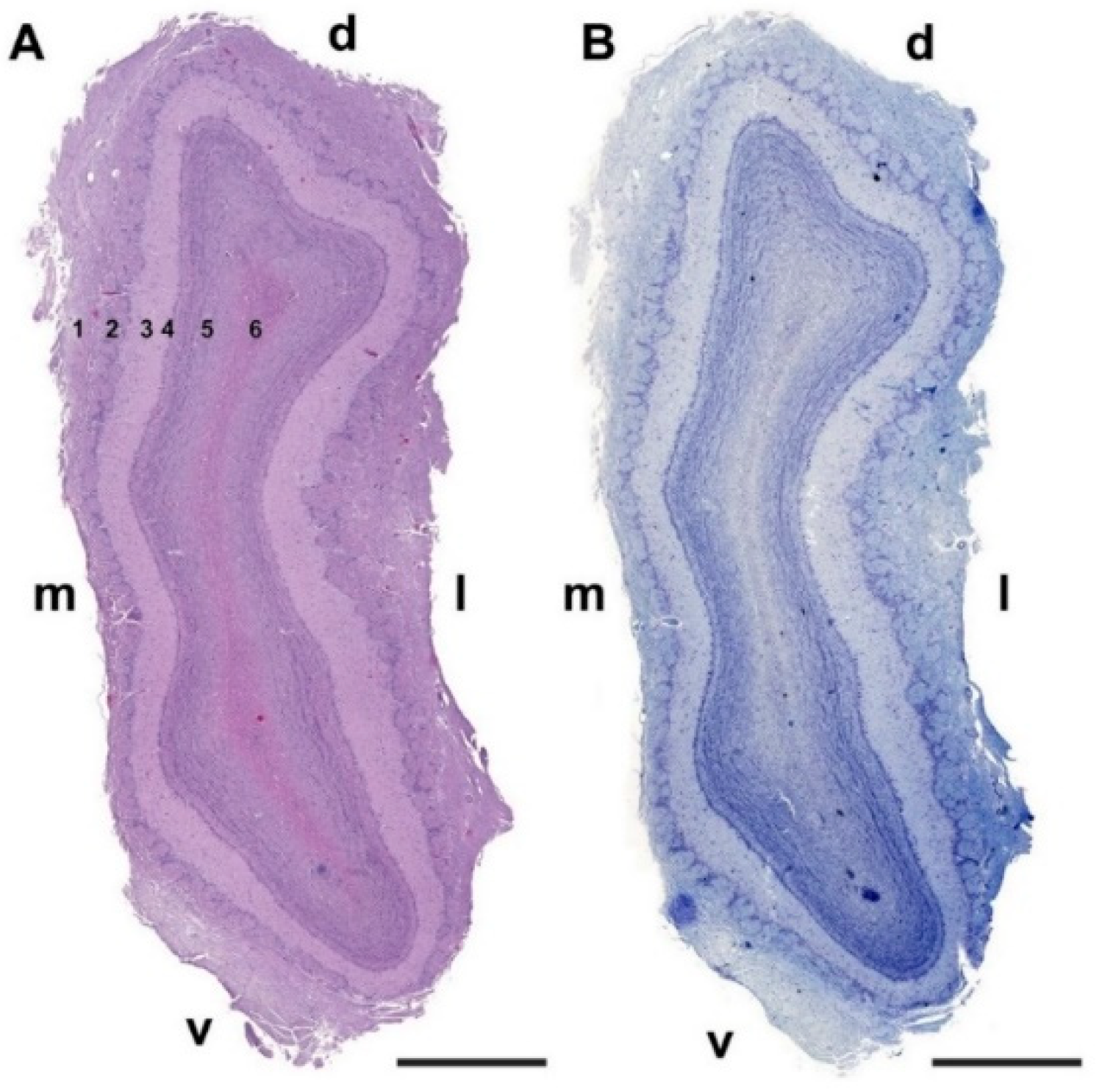
3.1.1. Macroscopic and Microscopic Anatomy
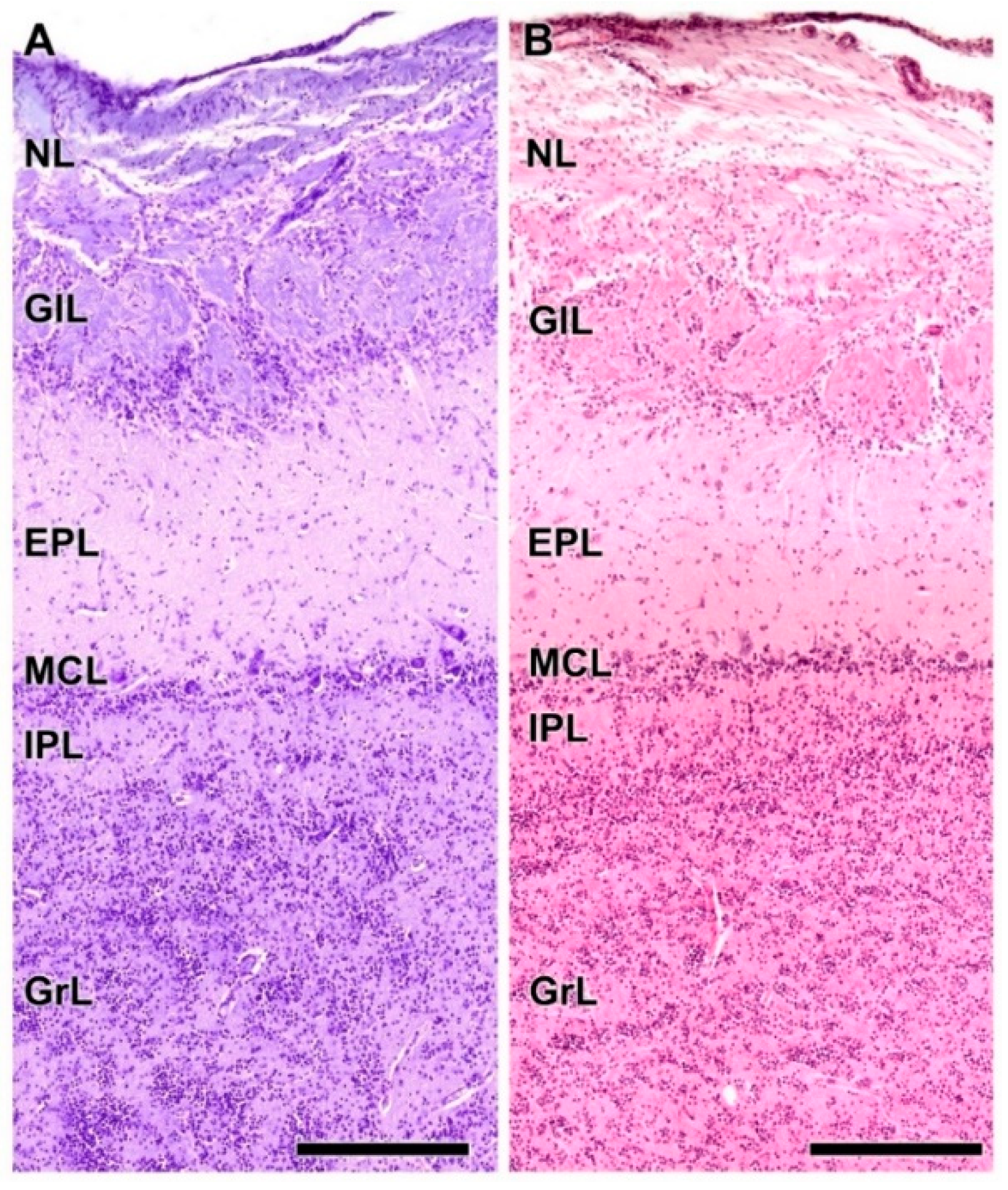
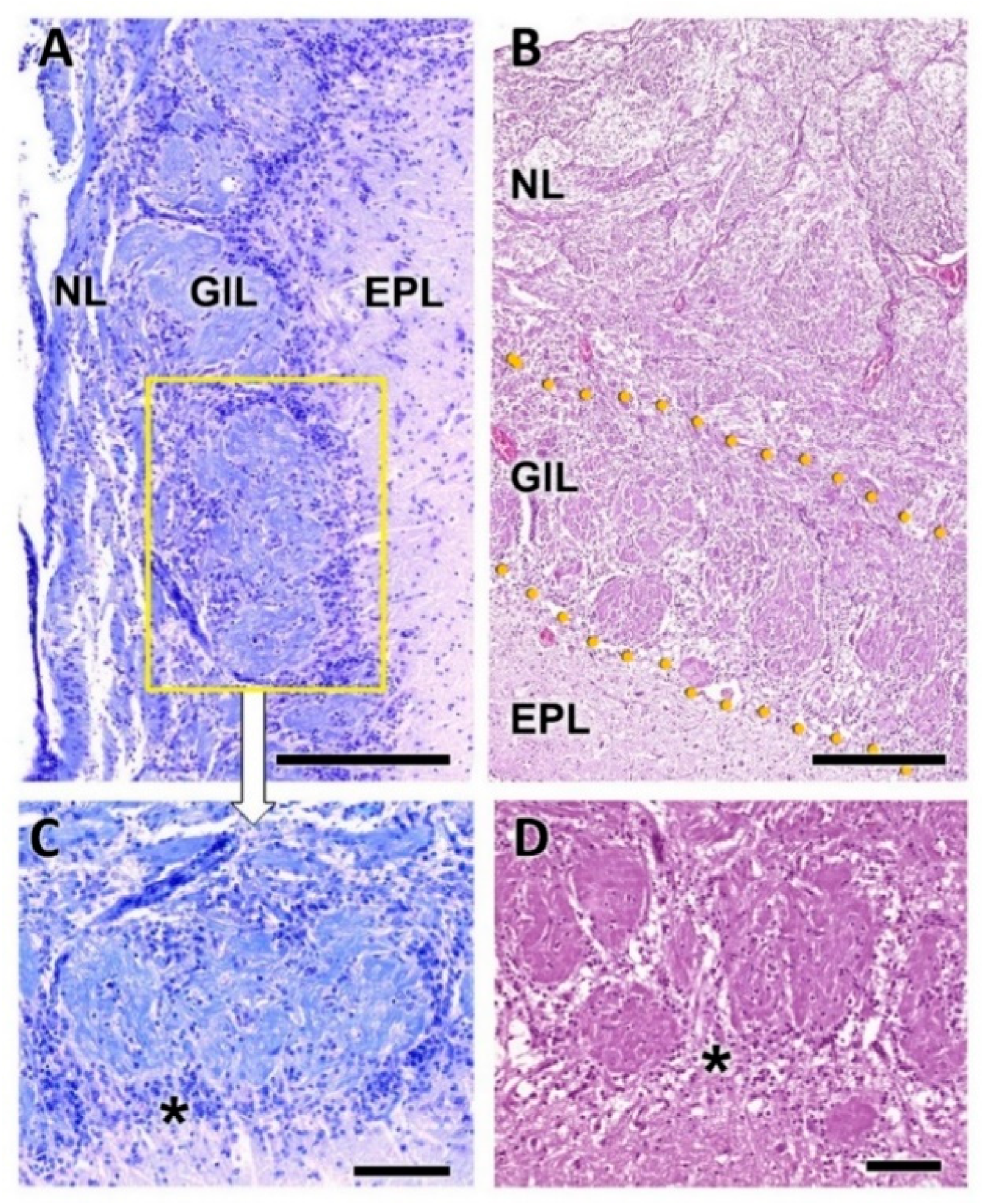
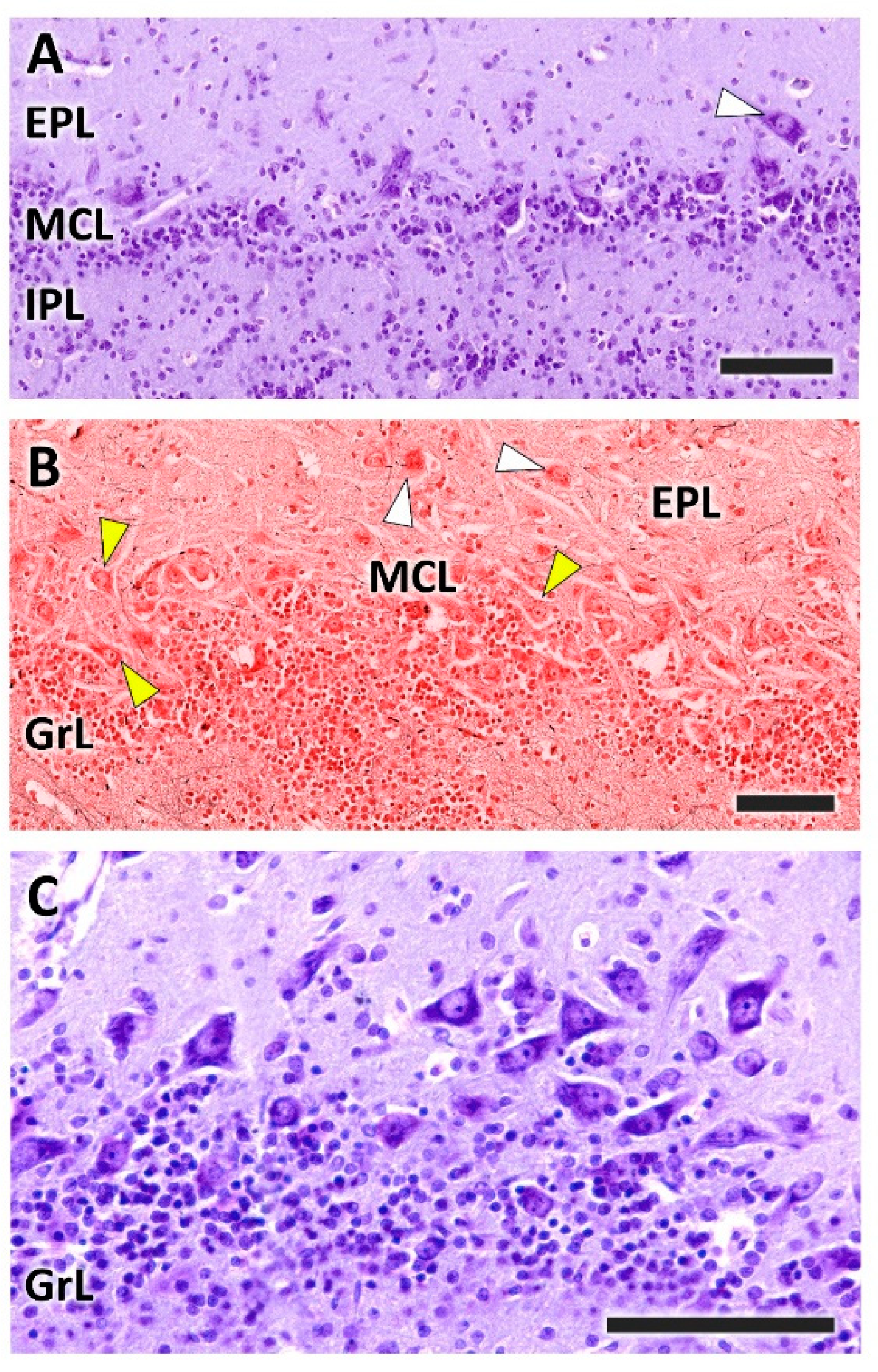
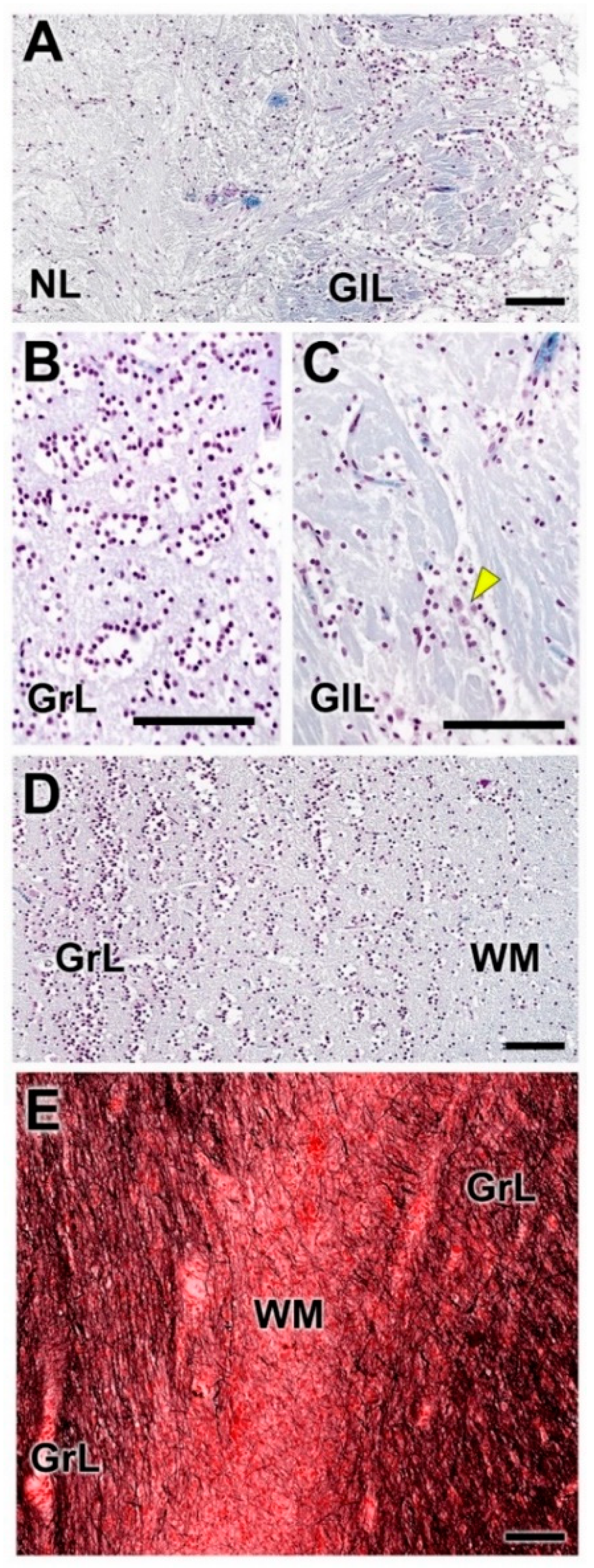
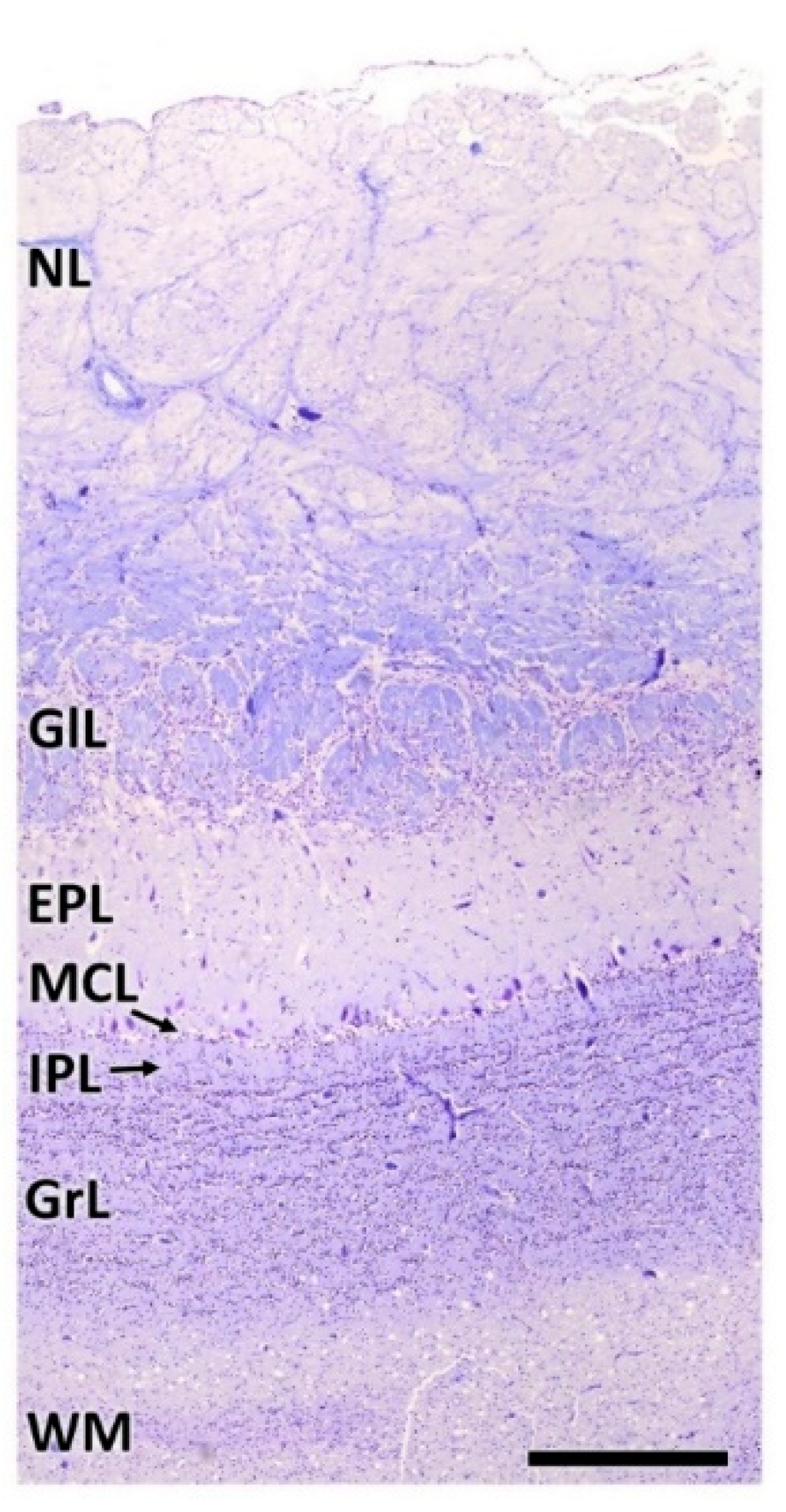
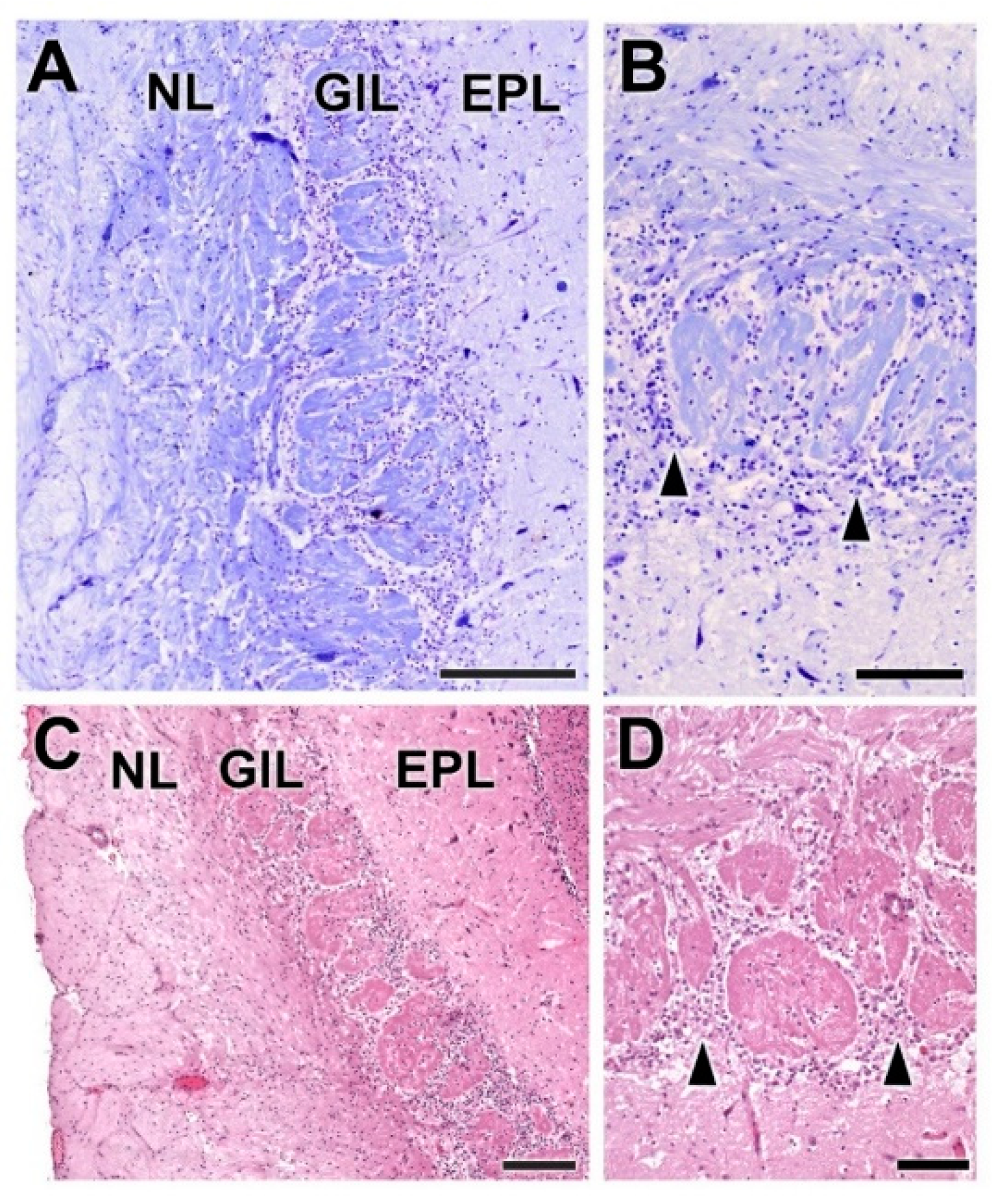
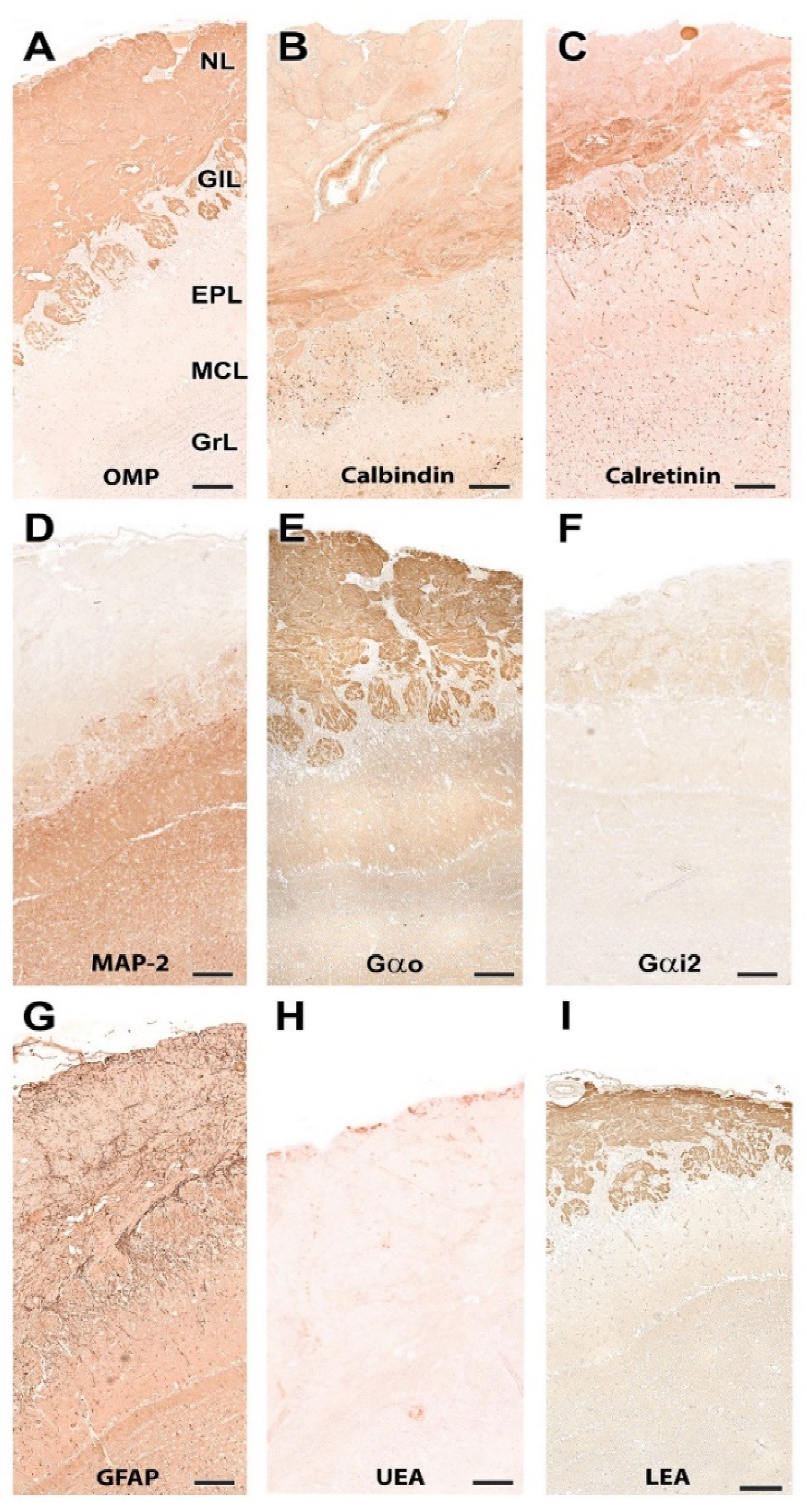
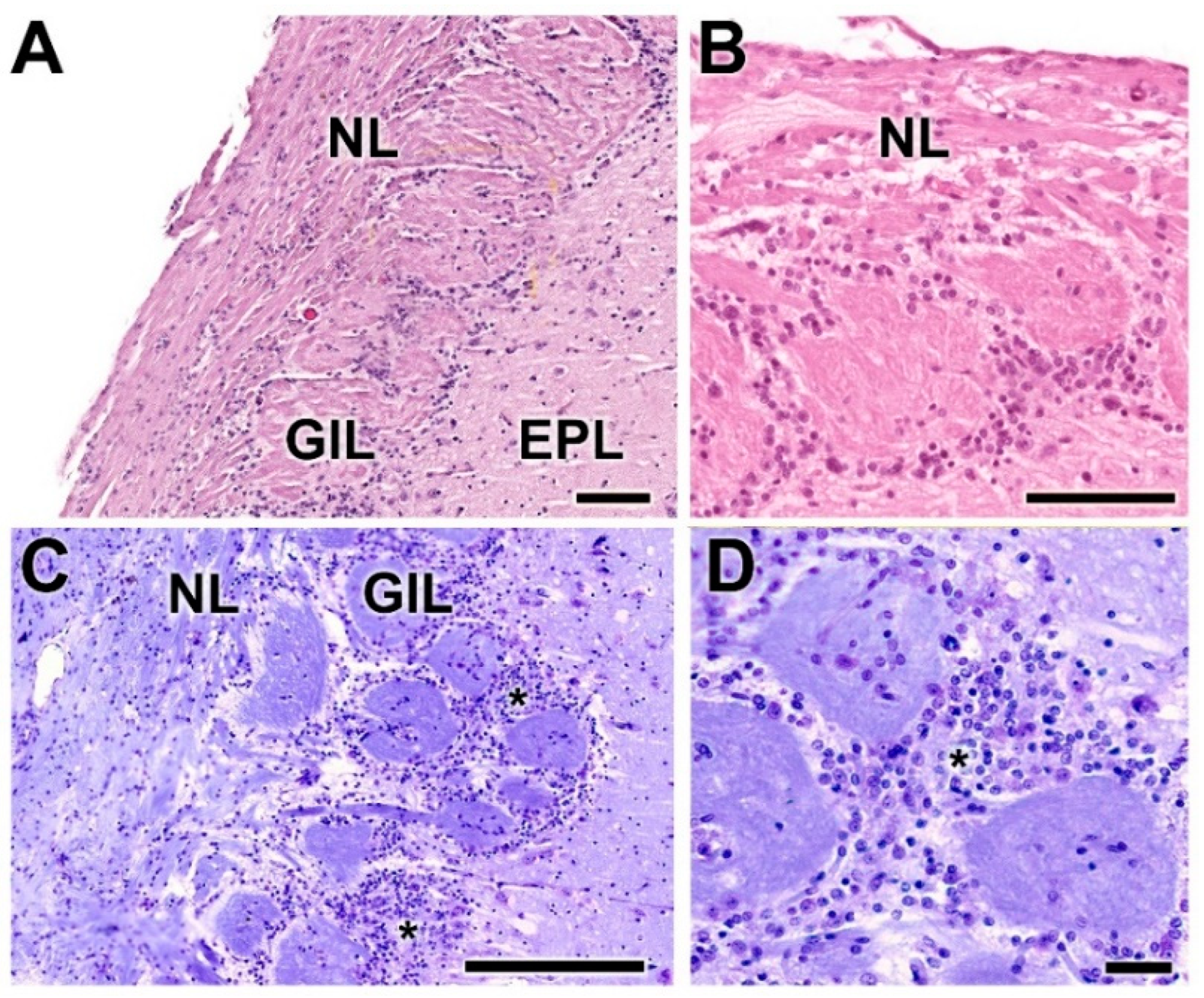
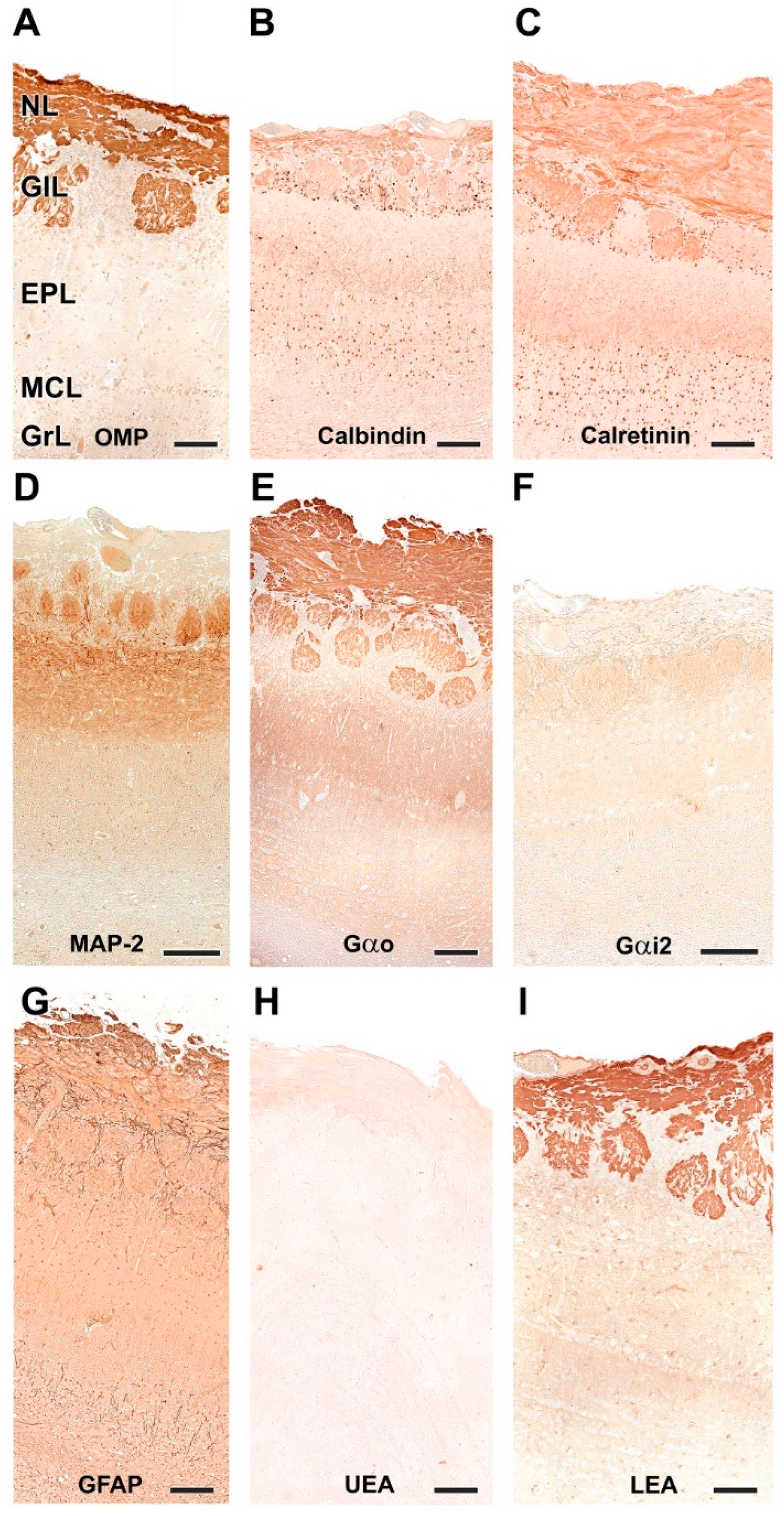
- Nerve layer (NL): formed by the axons of the olfactory nerves that reach the OB.
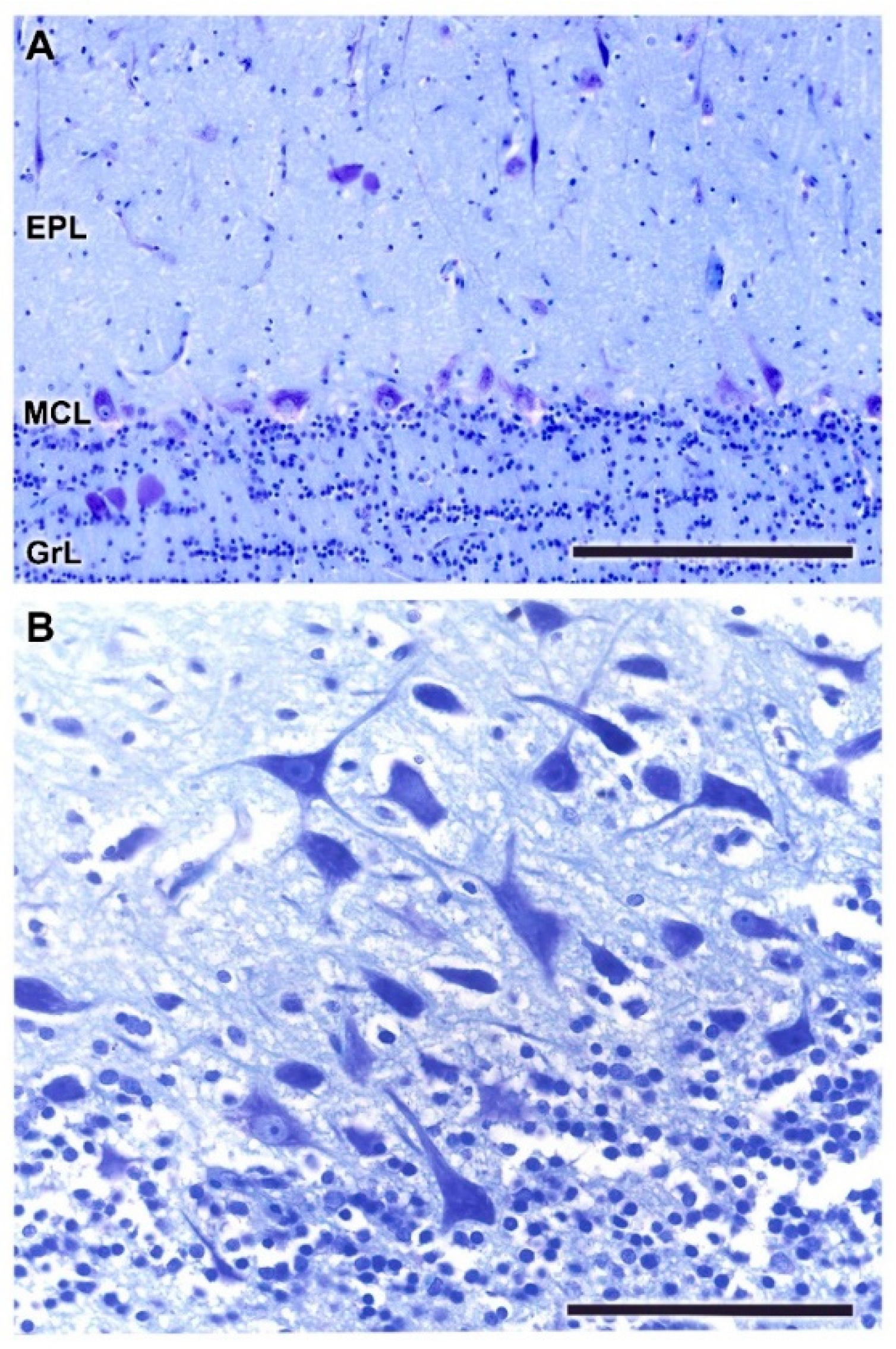
- Glomerular layer (GlL): comprised of glomeruli, spherical structures delimited by periglomerular (PG) cells. They correspond to the synapses of olfactory axons with the dendrites of mitral cells, which are the second neurons in the olfactory pathways.
- External plexiform layer (EPL): a nerve plexus mainly occupied by the dendrites of mitral cells.
- Mitral cell layer (MCL): a linear layer containing the somas of mitral cells.
- Internal plexiform layer (IPL): a thin band of white matter interposed between the mitral and granular layers.
- Granular layer (GrL): composed of large clusters of granule cells, which act as inhibitory neurons in the OB neural circuit.
- White matter (WM): formed by the projecting axons of the OB (Figure 2A,B).
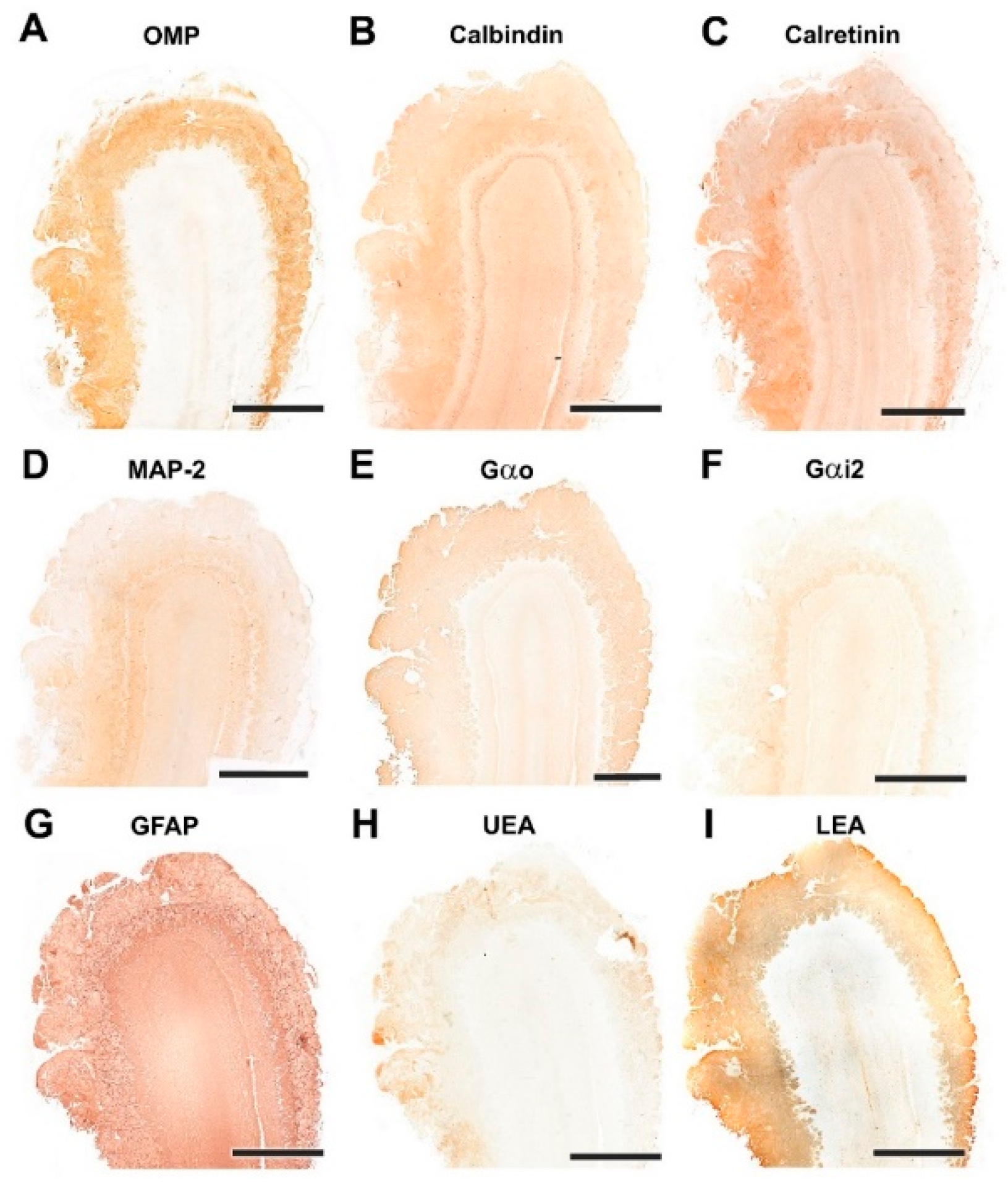
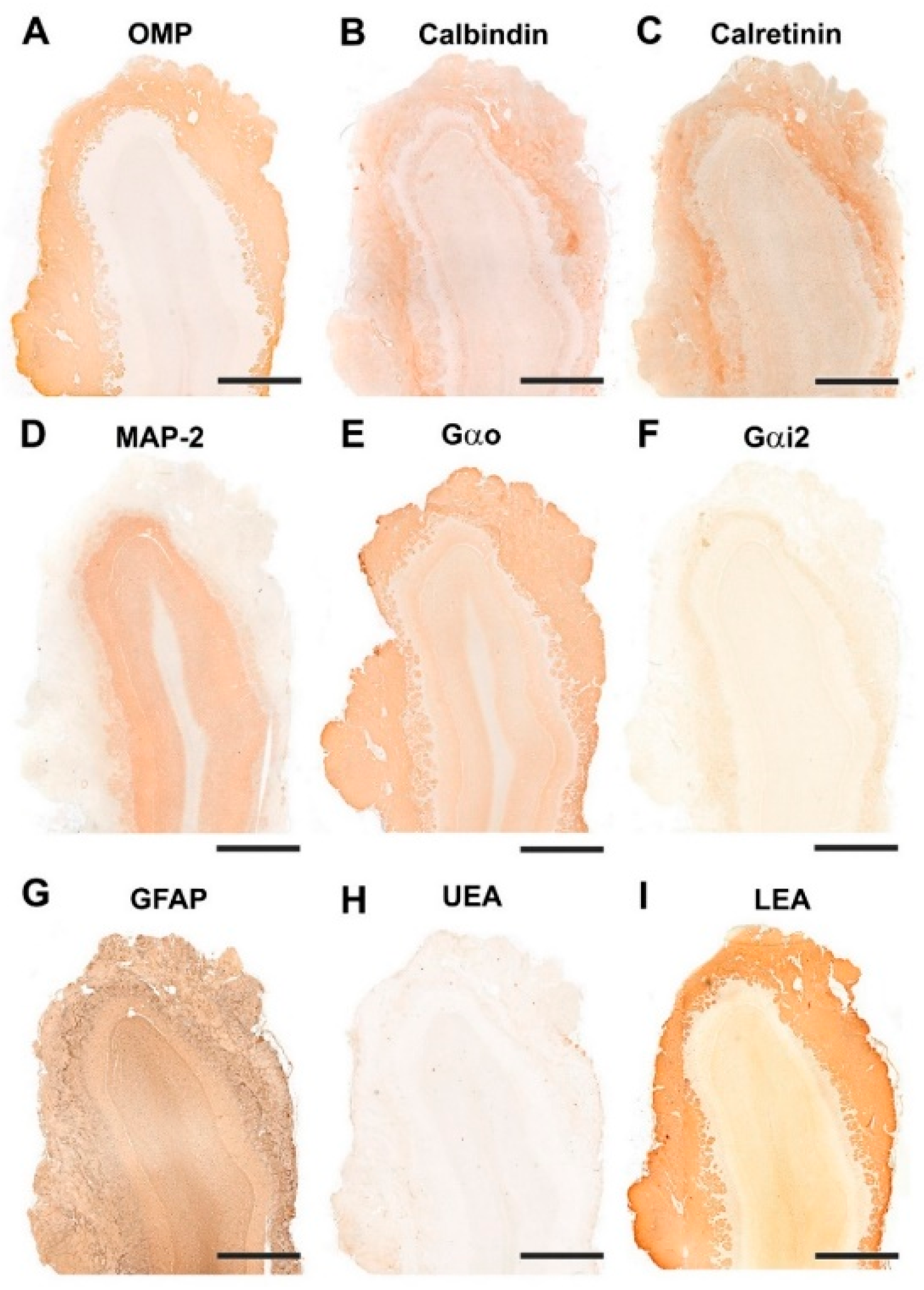
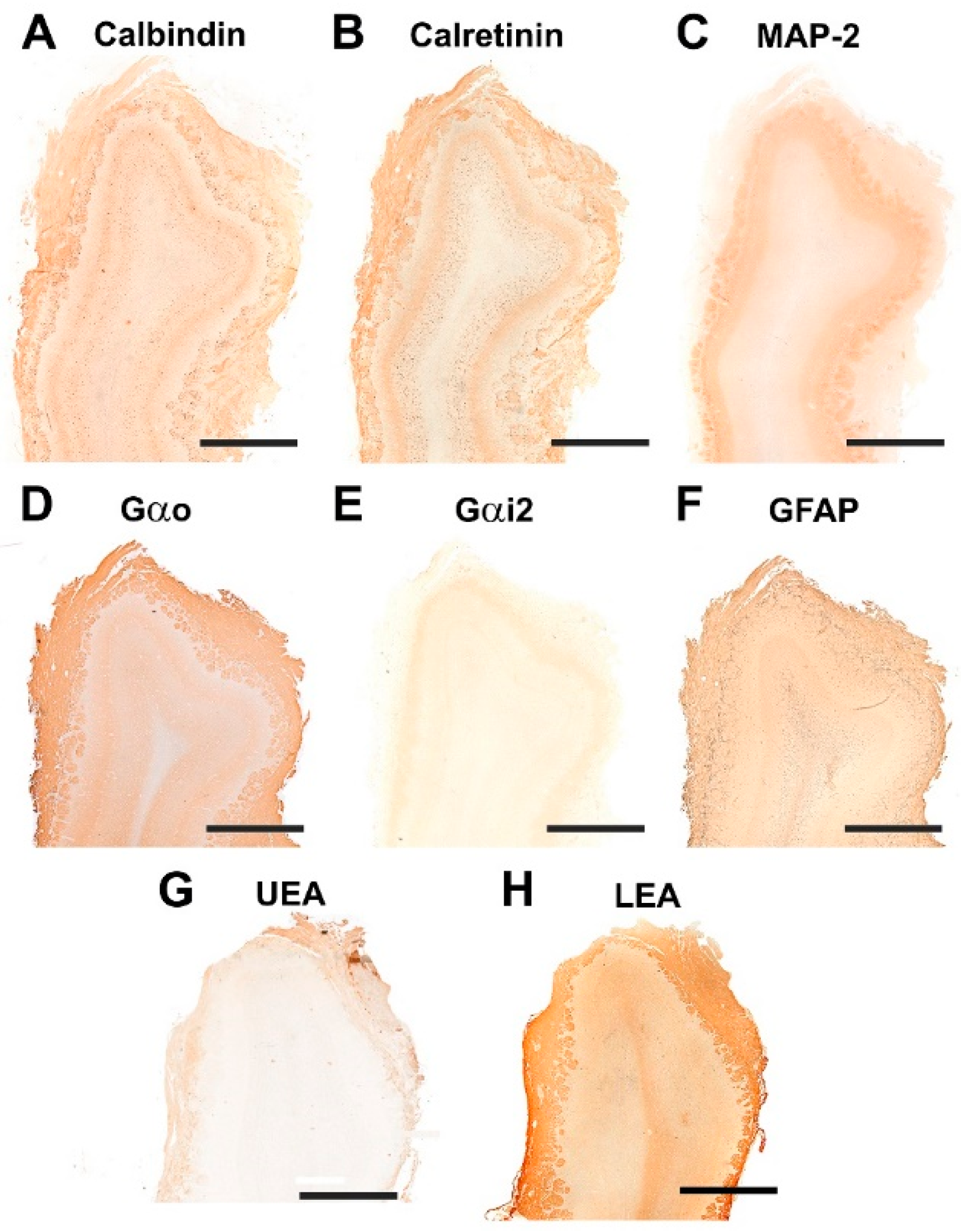
3.1.2. Immunohistochemical and Lectin Histochemical Study
3.2. Wolf Olfactory Bulb
3.2.1. Macroscopic and Microscopic Anatomy
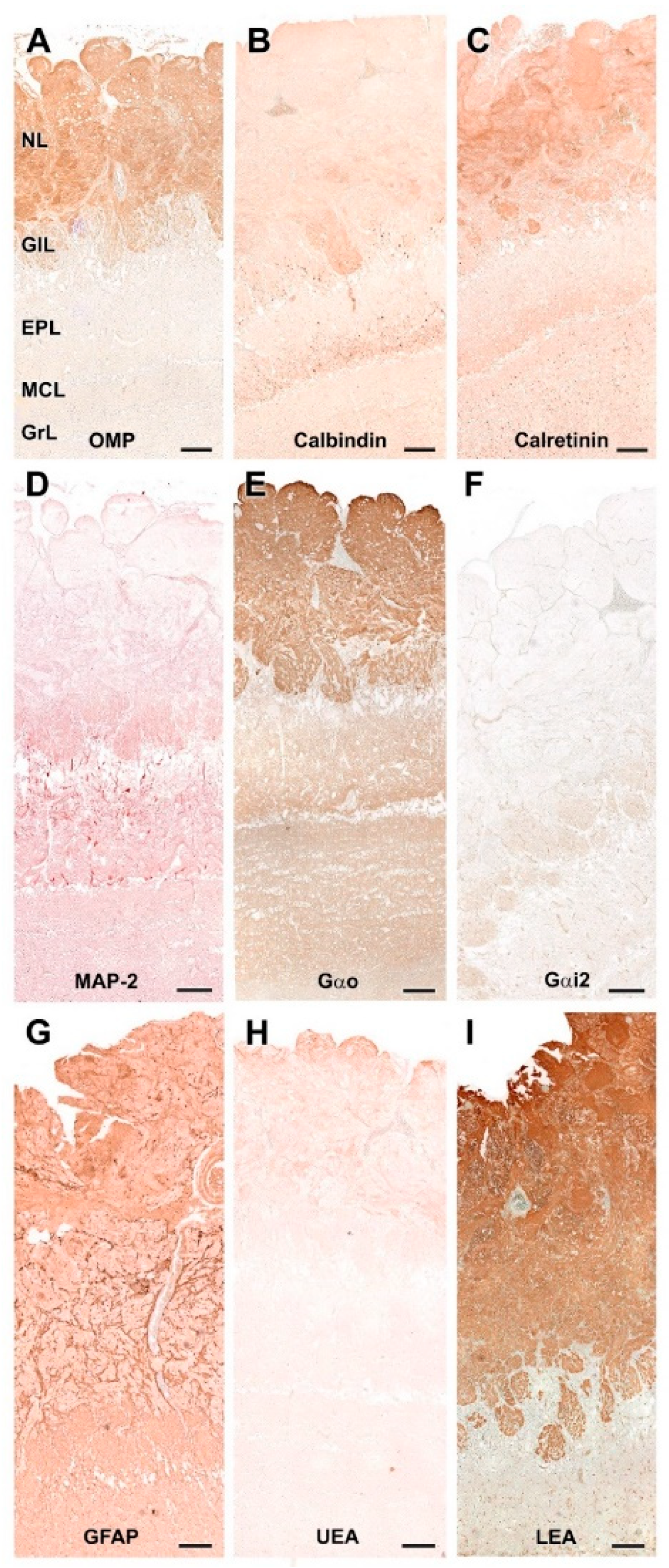
3.2.2. Immunohistochemical and Lectin Histochemical Study
3.3. Fox Olfactory Bulb
3.3.1. Macroscopic and Microscopic Anatomy
3.3.2. Immunohistochemical and Lectin Histochemical Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baum, M.J.; Cherry, J.A. Processing by the main olfactory system of chemosignals that facilitate mammalian reproduction. Horm. Behav. 2015, 68, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Kokocińska-Kusiak, A.; Woszczyło, M.; Zybala, M.; Maciocha, J.; Barłowska, K.; Dzięcioł, M. Canine Olfaction: Physiology, Behavior, and Possibilities for Practical Applications. Animals 2021, 11, 2463. [Google Scholar] [CrossRef] [PubMed]
- Whitlock, K.E.; Palominos, M.F. The Olfactory Tract: Basis for Future Evolution in Response to Rapidly Changing Ecological Niches. Front. Neuroanat. 2022, 16, 831602. [Google Scholar] [CrossRef] [PubMed]
- Touhara, K. Odor discrimination by G protein-coupled olfactory receptors. Microsc. Res. Tech. 2002, 58, 135. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.B.; Walker, J.C.; Cavnar, P.J.; Taylor, J.L.; Pickel, D.H.; Hall, S.B.; Suarez, J.C. Naturalistic quantification of canine olfactory sensitivity. Appl. Anim. Behav. Sci. 2006, 97, 241–254. [Google Scholar] [CrossRef]
- Salazar, I.; Sanchez-Quinteiro, P.; Barrios, A.W.; López Amado, M.; Vega, J.A. Anatomy of the olfactory mucosa. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 164, pp. 47–65. ISBN 978-0-444-63855-7. [Google Scholar]
- Smith, T.D.; Bhatnagar, K.P. Anatomy of the olfactory system. In Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2019; Volume 164, pp. 17–28. ISBN 978-0-444-63855-7. [Google Scholar]
- Arzi, A.; Rozenkrantz, L.; Gorodisky, L.; Rozenkrantz, D.; Holtzman, Y.; Ravia, A.; Bekinschtein, T.A.; Galperin, T.; Krimchansky, B.-Z.; Cohen, G.; et al. Olfactory sniffing signals consciousness in unresponsive patients with brain injuries. Nature 2020, 581, 428–433. [Google Scholar] [CrossRef]
- Ennis, M.; Holy, T.E. Anatomy and Neurobiology of the Main and Accessory Olfactory Bulbs. In Handbook of Olfaction and Gustation; Doty, R.L., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 157–182. ISBN 978-1-118-97175-8. [Google Scholar]
- Torres, M.V.; Ortiz-Leal, I.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Neuroanatomical and Immunohistological Study of the Main and Accessory Olfactory Bulbs of the Meerkat (Suricata suricatta). Animals 2021, 12, 91. [Google Scholar] [CrossRef]
- Powers, J.B.; Winans, S.S. Vomeronasal Organ: Critical Role in Mediating Sexual Behavior of the Male Hamster. Science 1975, 187, 961–963. [Google Scholar] [CrossRef]
- Keverne, E.B. Importance of olfactory and vomeronasal systems for male sexual function. Physiol. Behav. 2004, 83, 177–187. [Google Scholar] [CrossRef]
- Brennan, P.A.; Zufall, F. Pheromonal communication in vertebrates. Nature 2006, 444, 308–315. [Google Scholar] [CrossRef]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures, 2nd ed.; Cambridge University Press: Cambridge, UK, 2013; ISBN 978-1-139-03074-8. [Google Scholar]
- Liberles, S.D. Mammalian Pheromones. Annu. Rev. Physiol. 2014, 76, 151–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salazar, I.; Sánchez Quinteiro, P.; Cifuentes, J.M.; Fernández, P.; Lombardero, M. Distribution of the arterial supply to the vomeronasal organ in the cat. Anat. Rec. 1997, 247, 129–136. [Google Scholar] [CrossRef]
- Kondoh, D.; Tanaka, Y.; Kawai, Y.K.; Mineshige, T.; Watanabe, K.; Kobayashi, Y. Morphological and Histological Features of the Vomeronasal Organ in African Pygmy Hedgehog (Atelerix albiventris). Animals 2021, 11, 1462. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, D.; Watanabe, K.; Nishihara, K.; Ono, Y.S.; Nakamura, K.G.; Yuhara, K.; Tomikawa, S.; Sugimoto, M.; Kobayashi, S.; Horiuchi, N.; et al. Histological Properties of Main and Accessory Olfactory Bulbs in the Common Hippopotamus. Brain. Behav. Evol. 2017, 90, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. The vomeronasal system of the newborn capybara: A morphological and immunohistochemical study. Sci. Rep. 2020, 10, 13304. [Google Scholar] [CrossRef]
- Adams, D.R. The bovine vomeronasal organ. Arch. Histol. Jpn. 1986, 49, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; López-Beceiro, A.; Sanchez-Quinteiro, P. The vomeronasal organ of wild canids: The fox ( Vulpes vulpes ) as a model. J. Anat. 2020, 237, 890–906. [Google Scholar] [CrossRef]
- Nakajima, T.; Sakaue, M.; Kato, M.; Saito, S.; Ogawa, K.; Taniguchi, K. Immunohistochemical and enzyme-histochemical study on the accessory olfactory bulb of the dog. Anat. Rec. 1998, 252, 393–402. [Google Scholar] [CrossRef]
- Salazar, I.; Cifuentes, J.M.; Sánchez-Quinteiro, P. Morphological and Immunohistochemical Features of the Vomeronasal System in Dogs. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2013, 296, 146–155. [Google Scholar] [CrossRef]
- Salazar, I.; Barrios, A.W.; Sánchez-Quinteiro, P. Revisiting the Vomeronasal System From an Integrated Perspective: Revisiting the vomeronasal system. Anat. Rec. 2016, 299, 1488–1491. [Google Scholar] [CrossRef]
- Barrios, A.W.; Sanchez-Quinteiro, P.; Salazar, I. Dog and mouse: Toward a balanced view of the mammalian olfactory system. Front. Neuroanat. 2014, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jawlowski, H. On the bulbus olfactorius and bulbus olfactorius accessorius of some mammals. Lub. Uniw. Marii Curie-Sklodowskiej Roczniki. Annls Dzialc Nauk. Biol. 1956, 10, 67–86. [Google Scholar]
- Miodonski, R. Bulbus olfactorius of the dog (Canis familiaris). Acta Biol. Crac. 1968, 11. [Google Scholar]
- Choi, J.H.; Lee, C.H.; Yoo, K.-Y.; Hwang, I.K.; Lee, I.S.; Lee, Y.L.; Shin, H.-C.; Won, M.-H. Age-related Changes in Calbindin-D28k, Parvalbumin, and Calretinin Immunoreactivity in the Dog Main Olfactory Bulb. Cell. Mol. Neurobiol. 2010, 30, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chengetanai, S.; Bhagwandin, A.; Bertelsen, M.F.; Hård, T.; Hof, P.R.; Spocter, M.A.; Manger, P.R. The brain of the African wild dog. II. The olfactory system. J. Comp. Neurol. 2020, 528, 3285–3304. [Google Scholar] [CrossRef]
- Kruska, D. Mammalian Domestication and its Effect on Brain Structure and Behavior. In Intelligence and Evolutionary Biology; Jerison, H.J., Jerison, I., Eds.; Springer: Berlin/Heidelberg, Germany, 1988; pp. 211–250. ISBN 978-3-642-70879-4. [Google Scholar]
- Grewal, J.S.; Gloe, T.; Hegedus, J.; Bitterman, K.; Billings, B.K.; Chengetanai, S.; Bentil, S.; Wang, V.X.; Ng, J.C.; Tang, C.Y.; et al. Brain gyrification in wild and domestic canids: Has domestication changed the gyrification index in domestic dogs? J. Comp. Neurol. 2020, 528, 3209–3228. [Google Scholar] [CrossRef]
- Bird, D.J.; Jacquemetton, C.; Buelow, S.A.; Evans, A.W.; Van Valkenburgh, B. Domesticating olfaction: Dog breeds, including scent hounds, have reduced cribriform plate morphology relative to wolves. Anat. Rec. 2021, 304, 139–153. [Google Scholar] [CrossRef]
- Bird, D.J.; Murphy, W.J.; Fox-Rosales, L.; Hamid, I.; Eagle, R.A.; Van Valkenburgh, B. Olfaction written in bone: Cribriform plate size parallels olfactory receptor gene repertoires in Mammalia. Proc. R. Soc. B Biol. Sci. 2018, 285, 20180100. [Google Scholar] [CrossRef]
- Ortiz-Leal, I.; Torres, M.V.; Villamayor, P.R.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Can domestication shape canidae brain morphology? The accessory olfactory bulb of the red fox as a case in point. Ann. Anat. Anat. Anz. 2021, 240, 151881. [Google Scholar] [CrossRef]
- Tolivia, J.; Tolivia, D.; Navarro, A. New technique for differential staining of myelinated fibers and nerve cells on paraffin sections. Anat. Rec. 1988, 222, 437–440. [Google Scholar] [CrossRef]
- Villamayor, P.R.; Arana Álvaro, J.; Coppel, C.; Ortiz-Leal, I.; Torres, M.V.; Sanchez-Quinteiro, P.; Sánchez, L. A comprehensive structural, lectin and immunohistochemical characterization of the zebrafish olfactory system. Sci. Rep. 2021, 11, 8865. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P. A detailed morphological study of the vomeronasal organ and the accessory olfactory bulb of cats. Microsc. Res. Tech. 2011, 74, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- de la Rosa-Prieto, C.; Saiz-Sanchez, D.; Ubeda-Bañon, I.; Argandoña-Palacios, L.; Garcia-Muñozguren, S.; Martinez-Marcos, A. Neurogenesis in subclasses of vomeronasal sensory neurons in adult mice. Dev. Neurobiol. 2010, 70, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.H.; Saraswati, M.; Margolis, F.L. Immunolocalization of Bex protein in the mouse brain and olfactory system. J. Comp. Neurol. 2005, 487, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hermanowicz-Sobieraj, B.; Bogus-Nowakowska, K.; Robak, A. Calcium-binding proteins expression in the septum and cingulate cortex of the adult guinea pig. Ann. Anat. Anat. Anz. 2018, 215, 30–39. [Google Scholar] [CrossRef]
- Adrio, F.; Rodríguez-Moldes, I.; Anadón, R. Distribution of glycine immunoreactivity in the brain of the Siberian sturgeon (Acipenser baeri): Comparison with γ-aminobutyric acid. J. Comp. Neurol. 2011, 519, 1115–1142. [Google Scholar] [CrossRef]
- Kotani, T.; Murata, Y.; Ohnishi, H.; Mori, M.; Kusakari, S.; Saito, Y.; Okazawa, H.; Bixby, J.L.; Matozaki, T. Expression of PTPRO in the interneurons of adult mouse olfactory bulb. J. Comp. Neurol. 2010, 518, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Shibata, S.; Cho, K.H.; Kim, J.H.; Abe, H.; Murakami, G.; Cho, B.H. Expression of hyaluronan (hyaluronic acid) in the developing laminar architecture of the human fetal brain. Ann. Anat. Anat. Anz. 2013, 195, 424–430. [Google Scholar] [CrossRef]
- Zufall, F.; Leinders-Zufall, T. Mammalian pheromone sensing. Curr. Opin. Neurobiol. 2007, 17, 483–489. [Google Scholar] [CrossRef]
- Swaney, W.T.; Keverne, E.B. The evolution of pheromonal communication. Behav. Brain Res. 2009, 200, 239–247. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Distinct Evolutionary Patterns between Chemoreceptors of 2 Vertebrate Olfactory Systems and the Differential Tuning Hypothesis. Mol. Biol. Evol. 2008, 25, 1593–1601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meisami, E.; Bhatnagar, K.P. Structure and diversity in mammalian accessory olfactory bulb. Microsc. Res. Tech. 1998, 43, 476–499. [Google Scholar] [CrossRef]
- Grus, W.E.; Zhang, J. Rapid turnover and species-specificity of vomeronasal pheromone receptor genes in mice and rats. Gene 2004, 340, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Sánchez-Quinteiro, P. The risk of extrapolation in neuroanatomy: The case of the mammalian accessory olfactory bulb. Front. Neuroanat. 2009, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Siniscalchi, M. Olfaction and the canine brain. In Canine Olfaction Science and Law; Taylor & Francis Group: Boca Raton, FL, USA, 2016; pp. 31–37. [Google Scholar]
- Kruska, D.C.T. On the Evolutionary Significance of Encephalization in Some Eutherian Mammals: Effects of Adaptive Radiation, Domestication, and Feralization. Brain. Behav. Evol. 2005, 65, 73–108. [Google Scholar] [CrossRef]
- Jardim-Messeder, D.; Lambert, K.; Noctor, S.; Pestana, F.M.; de Castro Leal, M.E.; Bertelsen, M.F.; Alagaili, A.N.; Mohammad, O.B.; Manger, P.R.; Herculano-Houzel, S. Dogs Have the Most Neurons, Though Not the Largest Brain: Trade-Off between Body Mass and Number of Neurons in the Cerebral Cortex of Large Carnivoran Species. Front. Neuroanat. 2017, 11, 118. [Google Scholar] [CrossRef]
- Hecht, E.E.; Kukekova, A.V.; Gutman, D.A.; Acland, G.M.; Preuss, T.M.; Trut, L.N. Neuromorphological Changes following Selection for Tameness and Aggression in the Russian Farm-Fox experiment. J. Neurosci. 2021, 41, 6144–6156. [Google Scholar] [CrossRef]
- Kistner, T.M.; Zink, K.D.; Worthington, S.; Lieberman, D.E. Geometric morphometric investigation of craniofacial morphological change in domesticated silver foxes. Sci. Rep. 2021, 11, 2582. [Google Scholar] [CrossRef]
- Yan, X.; Joshi, A.; Zang, Y.; Assunção, F.; Fernandes, H.M.; Hummel, T. The Shape of the Olfactory Bulb Predicts Olfactory Function. Brain Sci. 2022, 12, 128. [Google Scholar] [CrossRef]
- Ngwenya, A.; Patzke, N.; Ihunwo, A.O.; Manger, P.R. Organisation and chemical neuroanatomy of the African elephant (Loxodonta africana) olfactory bulb. Brain Struct. Funct. 2011, 216, 403–416. [Google Scholar] [CrossRef]
- Dehmelt, L.; Halpain, S. The MAP2/Tau family of microtubule-associated proteins. Genome Biol. 2005, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villamayor, P.R.; Cifuentes, J.M.; Quintela, L.; Barcia, R.; Sanchez-Quinteiro, P. Structural, morphometric and immunohistochemical study of the rabbit accessory olfactory bulb. Brain Struct. Funct. 2020, 225, 203–226. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.S.; Halpern, M.; Ee, P.-L. Lectin histochemical identification of carbohydrate moieties in opossum chemosensory systems during development, with special emphasis on VVA-identified subdivisions in the accessory olfactory bulb. J. Morphol. 1995, 224, 331–349. [Google Scholar] [CrossRef] [PubMed]
- Salazar, I.; Quinteiro, P.S.; Cifuentes, J.M.; Lombardero, M. The Accessory Olfactory Bulb of the Mink, Mustela vison: A Morphological and Lectin Histochemical Study. Anat. Histol. Embryol. J. Vet. Med. Ser. C 1998, 27, 297–300. [Google Scholar] [CrossRef]
- Yang, W.; Choi, Y.; Park, C.; Lee, K.-H.; Ahn, M.; Kang, W.; Heo, S.-D.; Kim, J.; Shin, T. Histological and lectin histochemical studies in the vomeronasal organ of the Korean black goat, Capra hircus coreanae. Acta Histochem. 2021, 123, 151684. [Google Scholar] [CrossRef]
- Kondoh, D.; Kamikawa, A.; Sasaki, M.; Kitamura, N. Localization of α1-2 Fucose Glycan in the Mouse Olfactory Pathway. Cells Tissues Organs 2017, 203, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Villamayor, P.R.; Cifuentes, J.M.; Fdz-de-Troconiz, P.; Sanchez-Quinteiro, P. Morphological and immunohistochemical study of the rabbit vomeronasal organ. J. Anat. 2018, 233, 814–827. [Google Scholar] [CrossRef]
- Salazar, I. The Prenatal Maturity of the Accessory Olfactory Bulb in Pigs. Chem. Senses 2004, 29, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Park, C.; Ahn, M.; Lee, J.-Y.; Lee, S.; Yun, Y.; Lim, Y.-K.; Taniguchi, K.; Shin, T. A morphological study of the vomeronasal organ and the accessory olfactory bulb in the Korean roe deer, Capreolus pygargus. Acta Histochem. 2014, 116, 258–264. [Google Scholar] [CrossRef]
- Torres, M.V.; Ortiz-Leal, I.; Villamayor, P.R.; Ferreiro, A.; Rois, J.L.; Sanchez-Quinteiro, P. Does a third intermediate model for the vomeronasal processing of information exist? Insights from the macropodid neuroanatomy. Brain Struct. Funct. 2021, 227, 881–899. [Google Scholar] [CrossRef]
























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | 1st Ab Species/Dilution | 1st Ab Catalogue Number | Inmunogen | Reference | RRID | 2nd Ab Species/Dilution, Catalogue No. |
|---|---|---|---|---|---|---|
| Anti-Gαo | Rabbit/1:200 | Santa Cruz Biotech. sc-387 | Peptide mapping within a highly divergent domain of rat Gαo | [37] | AB_2111641 | ImmPRESS VR HRP Anti-rabbit IgG Reagent MP-6401-15 |
| Anti-Gαi2 | Rabbit/1:200 | Santa Cruz Biotech. sc-7276 | Peptide mapping within a highly divergent domain of rat Gαi2 | [38] | AB_2111472 | ImmPRESS VR HRP Anti-rabbit IgG Reagent MP-6401-15 |
| Anti-OMP | Goat/1:400 | Wako 544-10001 | Rodent olfactory marker protein | [39] | AB_2315007 | Horse anti-goat IgG 1:250 Vector BA-9500 |
| Anti-CB | Rabbit/1:6000 | Swant CB38 | Rat recombinant calbindin D-28k | [40] | AB_10000340 | ImmPRESS VR HRP Anti-rabbit IgG Reagent MP-6401-15 |
| Anti-CR | Rabbit/1:6000 | Swant 7697 | Recombinant human calretinin with a 6-his tag at the N-terminus | [41] | AB_2619710 | ImmPRESS VR HRP Anti-rabbit IgG Reagent MP-6401-15 |
| Anti-MAP-2 | Mouse/1:400 | Sigma M4403 | Rat brain microtubule-associated proteins | [42] | AB_477193 | ImmPRESS VR HRP Anti-mouse IgG Reagent MP-6402-15 |
| Anti-GFAP | Rabbit/1:400 | Dako Z0334 | GFAP from bovine spinal cord | [43] | AB_10013382 | ImmPRESS VR HRP Anti-rabbit IgG Reagent MP-6401-15 |
| UEA | 1:60 | Vector L-1060 | Rabbit 1:50 DAKO P289 | |||
| LEA | 20 µm/mL | Vector B-1175 | Vectastain ABC reagent PK-4000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz-Leal, I.; Torres, M.V.; López-Callejo, L.N.; Fidalgo, L.E.; López-Beceiro, A.; Sanchez-Quinteiro, P. Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox. Animals 2022, 12, 1079. https://doi.org/10.3390/ani12091079
Ortiz-Leal I, Torres MV, López-Callejo LN, Fidalgo LE, López-Beceiro A, Sanchez-Quinteiro P. Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox. Animals. 2022; 12(9):1079. https://doi.org/10.3390/ani12091079
Chicago/Turabian StyleOrtiz-Leal, Irene, Mateo V. Torres, Linda Noa López-Callejo, Luis Eusebio Fidalgo, Ana López-Beceiro, and Pablo Sanchez-Quinteiro. 2022. "Comparative Neuroanatomical Study of the Main Olfactory Bulb in Domestic and Wild Canids: Dog, Wolf and Red Fox" Animals 12, no. 9: 1079. https://doi.org/10.3390/ani12091079