
Feeding Behaviour and Bioavailability of Essential Amino Acids in Shrimp Penaeus monodon Fed Fresh and Leached Fishmeal and Fishmeal-Free Diets


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Formulation, Composition and Macronutrient Digestibility
2.2. Feeding Behaviour Experiments
2.2.1. Feed Intake Calculation
2.2.2. Feed Intake and Appetite Revival after Single and Multiple Deliveries of Food
2.2.3. Feed Intake Response on Leached out Feeds
2.3. Quantifying Nutrient Losses through Feed Leaching
2.4. Haemolymph Amino Acids
2.5. Chemical Composition Analyses
2.6. Statistical Analyses
3. Results
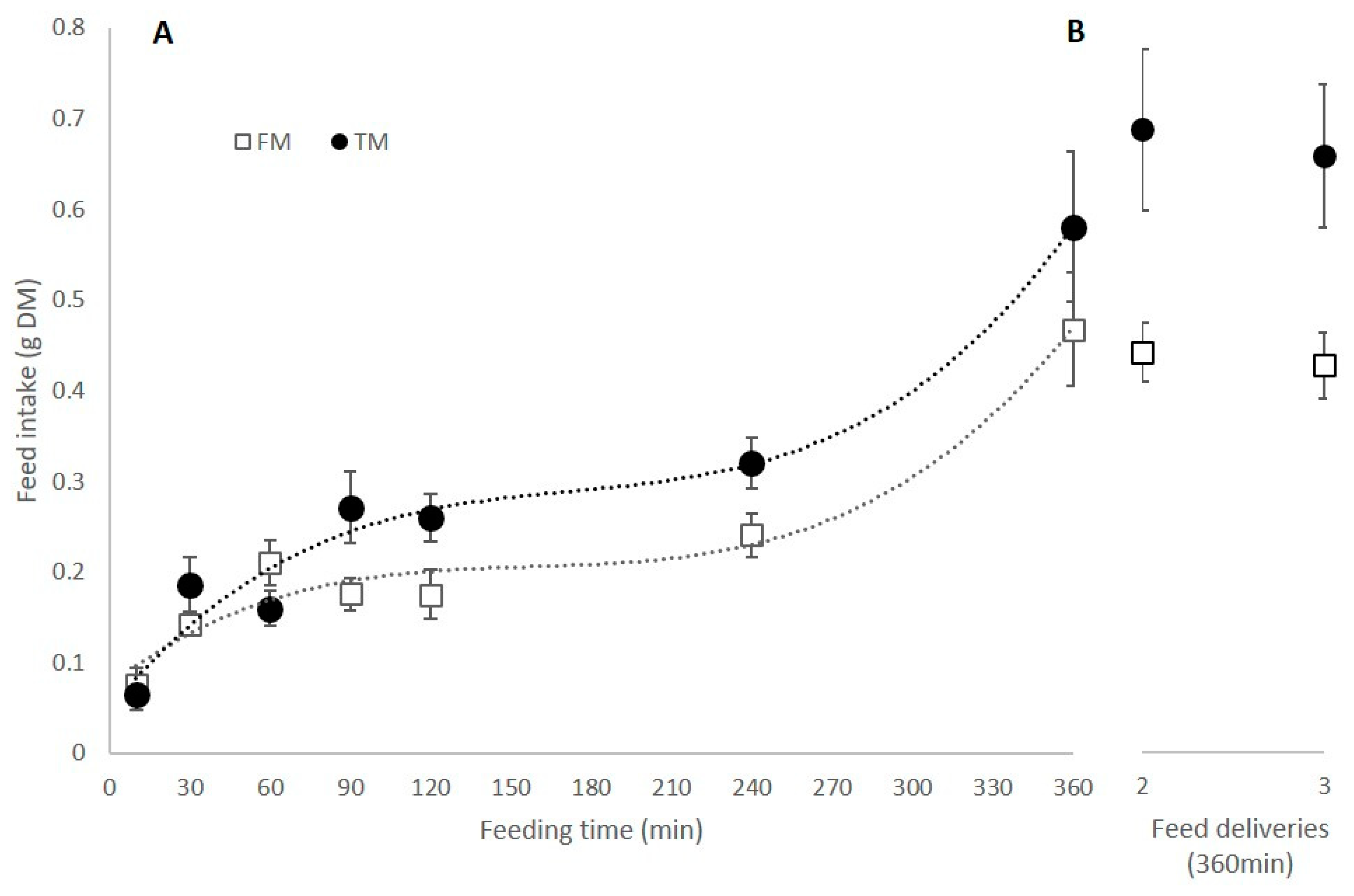
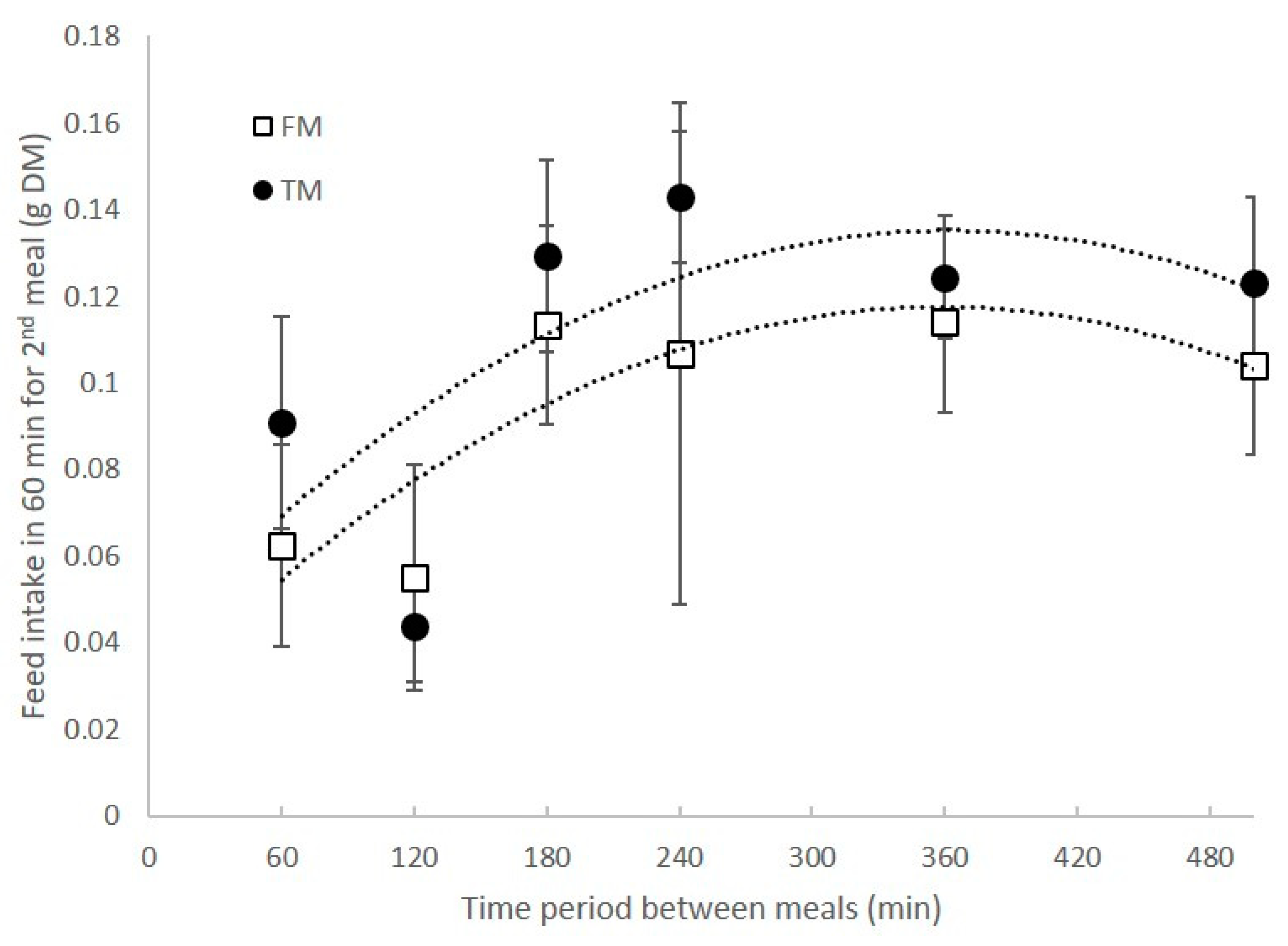
3.1. Feed Intake and Appetite Revival after Single and Multiple Deliveries of Food
3.2. Feed Apparent Digestibility
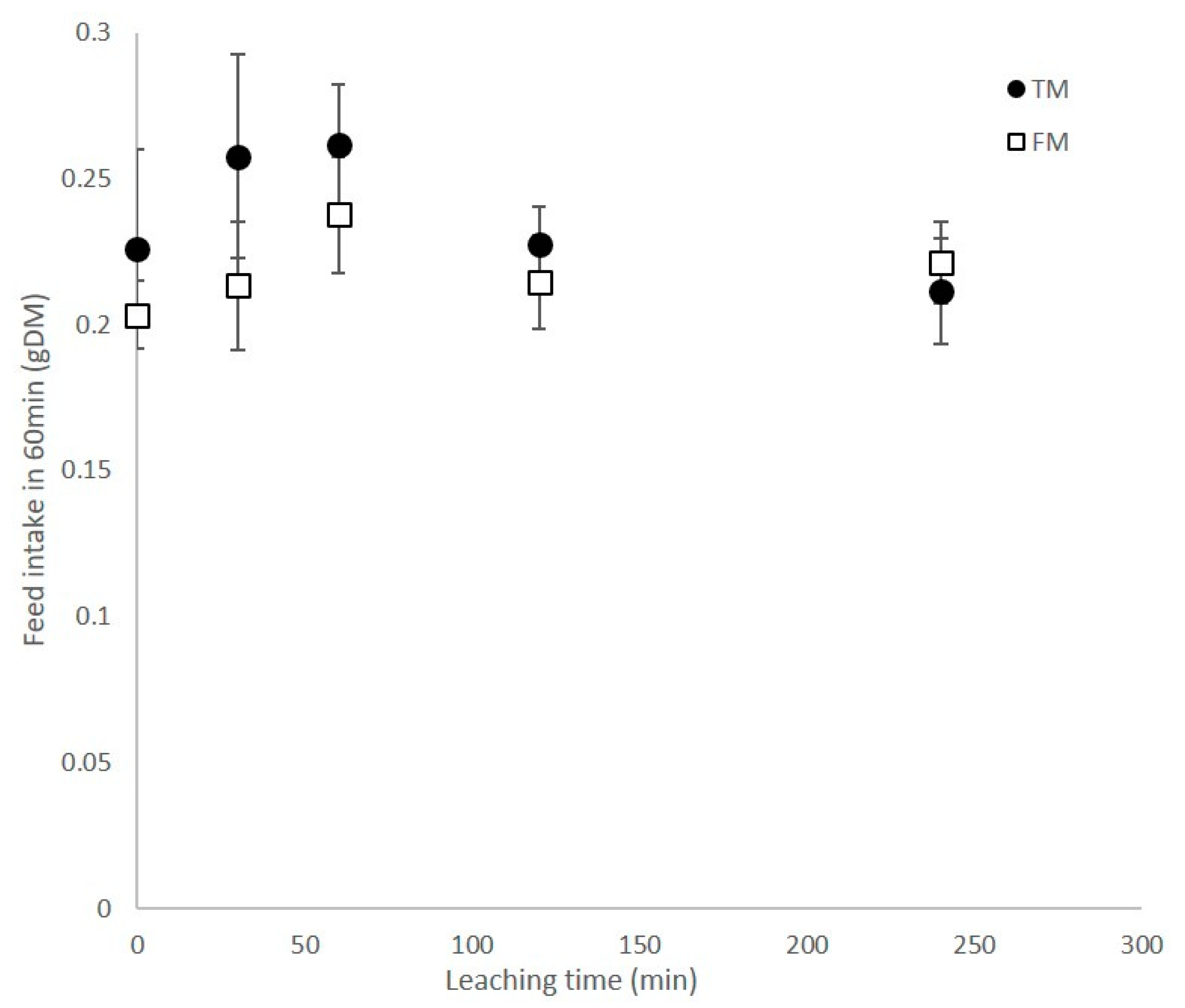
3.3. Feed stability and Leaching of Protein and Amino Acids
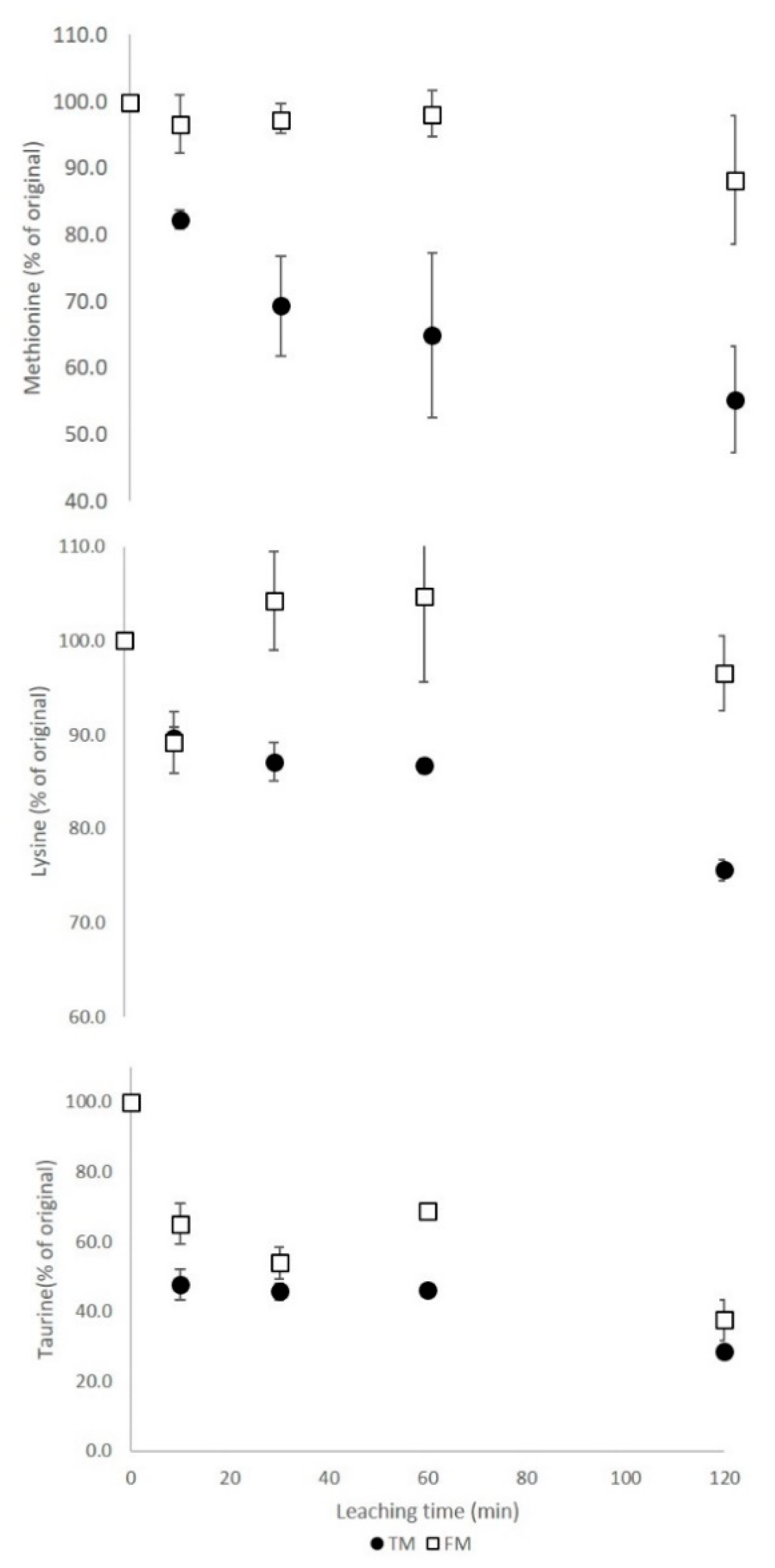
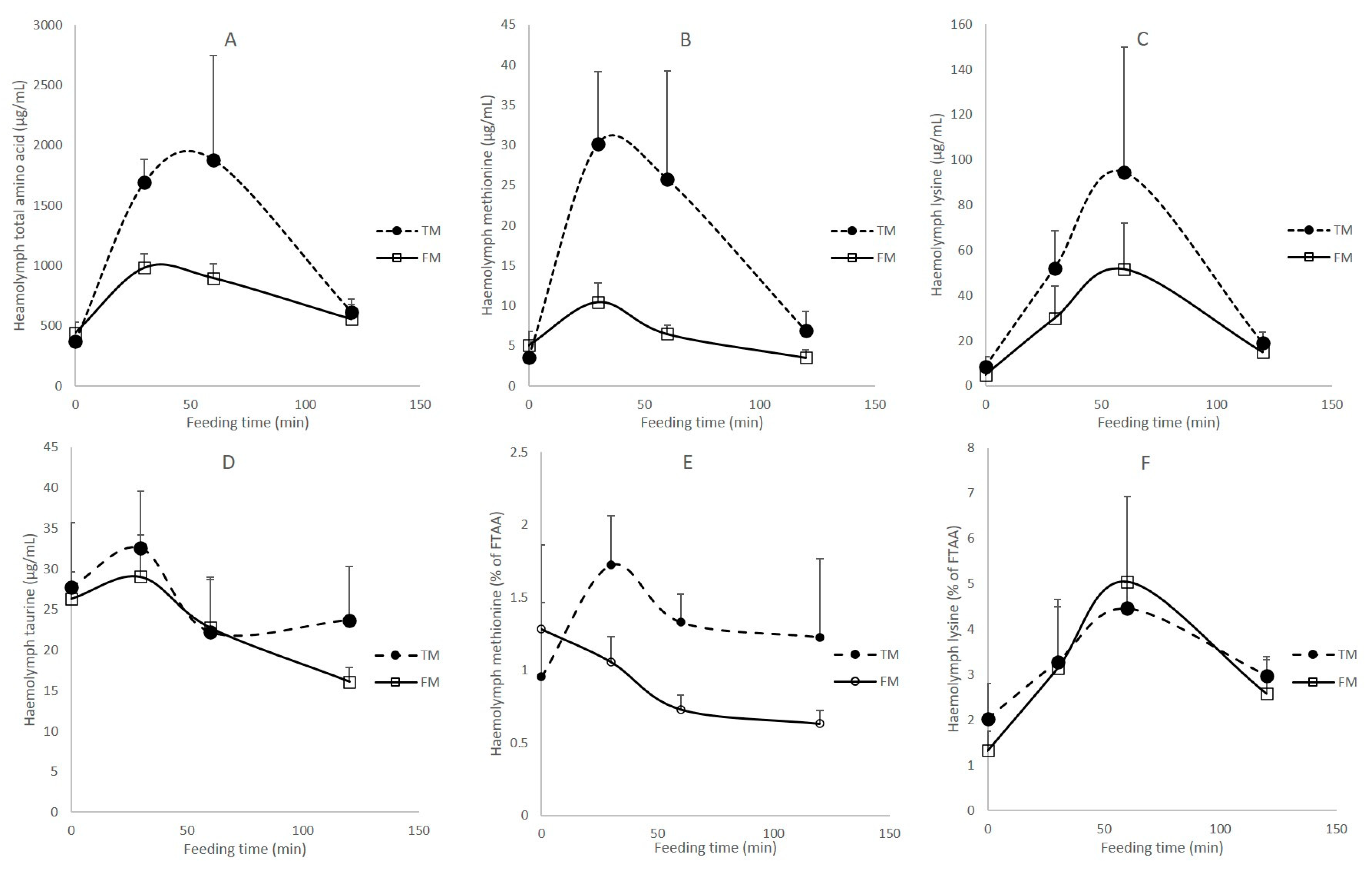
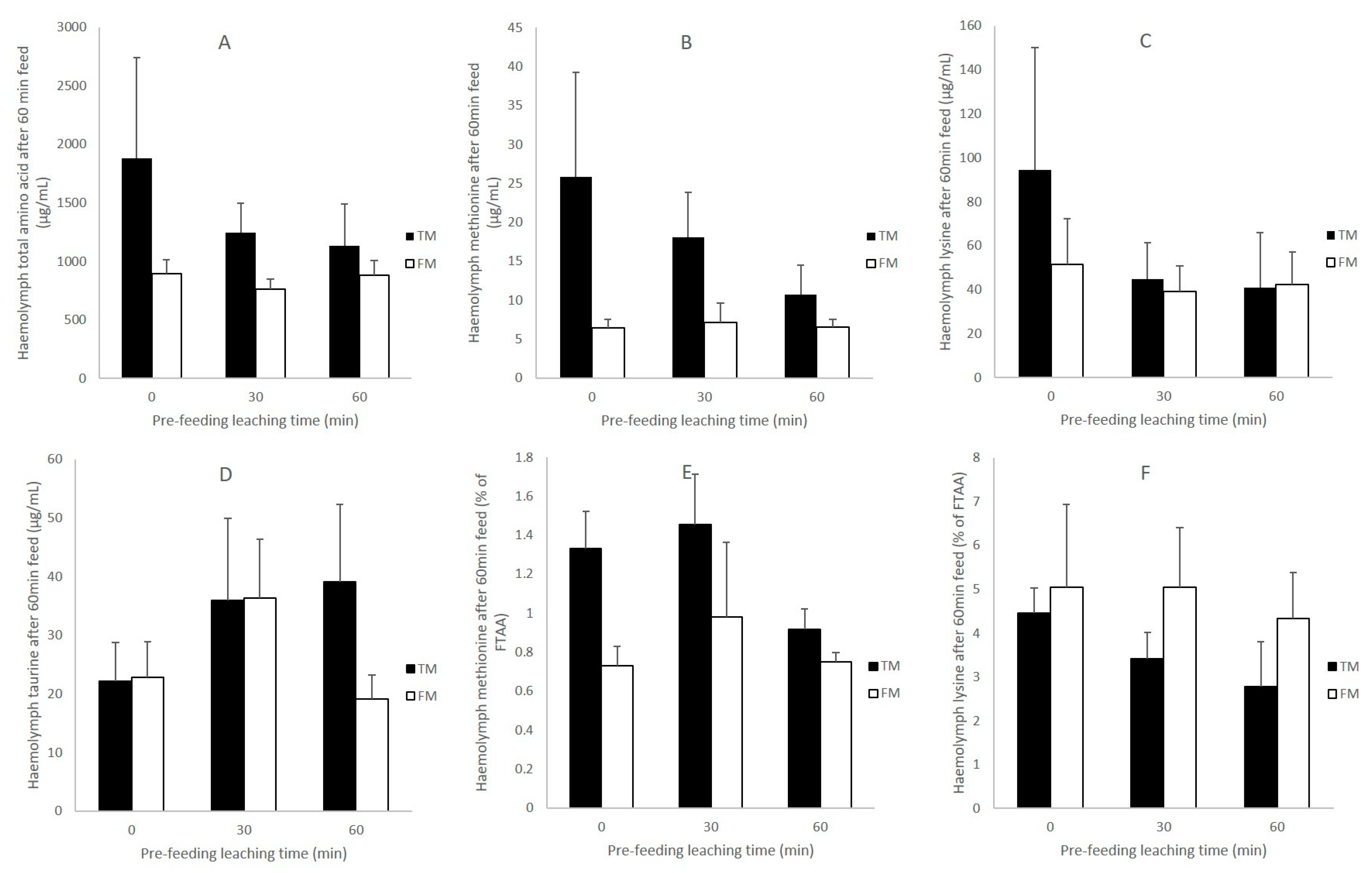
3.4. Haemolymph Amino Acids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvarez, J.S.; Galindo, J.; Fraga, I.; García, T.; Villarreal, H.; Hernández-Llamas, A. Substitution of fishmeal with soybean meal in practical diets for juvenile white shrimp Litopenaeus schmitti (Pérez-Farfante & Kensley 1997). Aquac. Res. 2007, 38, 689–695. [Google Scholar] [CrossRef]
- Cummins, V.C.; Webster, C.D.; Thompson, K.R.; Velasquez, A. Replacement of Fish Meal with Soybean Meal, Alone or in Combination with Distiller’s Dried Grains with Solubles in Practical Diets for Pacific White Shrimp, Litopenaeus vannamei, Grown in a Clear-Water Culture System. J. World Aquac. Soc. 2013, 44, 775–785. [Google Scholar] [CrossRef]
- Glencross, B.; Irvin, S.; Arnold, S.; Blyth, D.; Bourne, N.; Preston, N. Effective use of microbial biomass products to facilitate the complete replacement of fishery resources in diets for the black tiger shrimp, Penaeus monodon. Aquaculture 2014, 431, 12–19. [Google Scholar] [CrossRef]
- Ayisi, C.L.; Hua, X.; Apraku, A.; Afriyie, G.; Kyei, B.A. Recent Studies toward the Development of Practical Diets for Shrimp and Their Nutritional Requirements. HAYATI J. Biosci. 2017, 24, 109–117. [Google Scholar] [CrossRef]
- Cummins, V.C.; Rawles, S.D.; Thompson, K.R.; Velasquez, A.; Kobayashi, Y.; Hager, J.; Webster, C.D. Evaluation of black soldier fly (Hermetia illucens) larvae meal as partial or total replacement of marine fish meal in practical diets for Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2017, 473, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Richard, L.; Surget, A.; Rigolet, V.; Kaushik, S.J.; Geurden, I. Availability of essential amino acids, nutrient utilisation and growth in juvenile black tiger shrimp, Penaeus monodon, following fishmeal replacement by plant protein. Aquaculture 2011, 322–323, 109–116. [Google Scholar] [CrossRef]
- Sudaryono, A.; Tsvetnenko, E.; Evans, L.H. Evaluation of potential of lupin meal as an alternative to fish meal in juvenile Penaeus monodon diets. Aquac. Nutr. 1999, 5, 277–285. [Google Scholar] [CrossRef]
- Simon, C.J.; Truong, H.H.; Noble, T.H.; Osborne, S.A.; Wynne, J.W.; Wade, N.M. Microbial biomass, marine invertebrate meals and feed restriction influence the biological and gut microbiota response of shrimp Penaeus monodon. Aquaculture 2020, 520, 734679. [Google Scholar] [CrossRef]
- Leber, K.M.; Pruder, G.D. Using Experimental Microcosms in Shrimp Research: The Growth-Enhancing Effect of Shrimp Pond Water. J. World Aquac. Soc. 1988, 19, 197–203. [Google Scholar] [CrossRef]
- Amaya, E.A.; Davis, D.A.; Rouse, D.B. Replacement of fish meal in practical diets for the Pacific white shrimp (Litopenaeus vannamei) reared under pond conditions. Aquaculture 2007, 262, 393–401. [Google Scholar] [CrossRef]
- Sookying, D.; Davis, D.A. Pond production of Pacific white shrimp (Litopenaeus vannamei) fed high levels of soybean meal in various combinations. Aquaculture 2011, 319, 141–149. [Google Scholar] [CrossRef]
- Nunes, A.J.P.; Sá, M.V.C.; Browdy, C.L.; Vazquez-Anon, M. Practical supplementation of shrimp and fish feeds with crystalline amino acids. Aquaculture 2014, 431, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Nunes, A.J.P.; Sabry-Neto, H.; da Silva, F.H.P.; de Oliveira-Neto, A.R.; Masagounder, K. Multiple feedings enhance the growth performance and feed efficiency of juvenile Litopenaeus vannamei when fed a low-fish meal amino acid-supplemented diet. Aquac. Int. 2019, 27, 337–347. [Google Scholar] [CrossRef]
- Ullman, C.; Rhodes, M.; Hanson, T.; Cline, D.; Davis, D.A. Effects of Four Different Feeding Techniques on the Pond Culture of Pacific White Shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2019, 50, 54–64. [Google Scholar] [CrossRef] [Green Version]
- Arnold, S.; Smullen, R.P.; Briggs, M.; West, M.; Glencross, B. The combined effect of feed frequency and ration size of diets with and without microbial biomass on the growth and feed conversion of juvenile Penaeus monodon. Aquac. Nutr. 2016, 22, 1340–1347. [Google Scholar] [CrossRef]
- Truong, H.H.; Hines, B.M.; Rombenso, A.N.; Simon, C.J. Feed intake, gastro-intestinal transit and haemolymph free amino acids in the shrimp Penaeus monodon are influenced by marine meal supplementation. Aquaculture 2020, 533, 736171. [Google Scholar] [CrossRef]
- Guo, J.; Duan, M.; Qiu, X.; Masagounder, K.; Davis, D.A. Characterization of methionine uptake and clearance in the hemolymph of Pacific white shrimp Litopenaeus vannamei. Aquaculture 2020, 526, 735351. [Google Scholar] [CrossRef]
- Glencross, B.; Arnold, S.; Irvin, S. Bioactive factors in microbial biomass have the capacity to offset reductions in the level of protein in the diet of black tiger shrimp, Penaeus monodon. Aquaculture 2015, 446, 74–79. [Google Scholar] [CrossRef]
- Rombenso, A.N.; Truong, H.; Simon, C. Dietary butyrate alone or in combination with succinate and fumarate improved survival, feed intake, growth and nutrient retention efficiency of juvenile Penaeus monodon. Aquaculture 2020, 528, 734679. [Google Scholar] [CrossRef]
- Wade, N.; Bourne, N.; Simon, C. Influence of marker particle size on nutrient digestibility measurements and particle movement through the digestive system of shrimp. Aquaculture 2018, 491, 273–280. [Google Scholar] [CrossRef]
- Simon, C.J.; Jeffs, A. The effect of dietary carbohydrate on the appetite revival and glucose metabolism of juveniles of the spiny lobster, Jasus edwardsii. Aquaculture 2013, 384–387, 111–118. [Google Scholar] [CrossRef]
- Obaldo, L.G.; Divakaran, S.; Tacon, A.G. Method for determining the physical stability of shrimp feeds in water. Aquac. Res. 2002, 33, 369–377. [Google Scholar] [CrossRef]
- Berry, S.E.; Simon, C.J.; Foote, A.R.; Jerry, D.R.; Wade, N.M. Evaluation of baseline haemolymph biochemistry, volume and total body energetics to determine an accurate condition index in the black tiger shrimp, Penaeus monodon. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2019, 228, 1–9. [Google Scholar] [CrossRef]
- Davey, J.F.; Ersser, R.S. Amino acid analysis of physiological fluids by high-performance liquid chromatography with phenylisothiocyanate derivatization and comparison with ion-exchange chromatography. J. Chromatogr. B Biomed. Sci. Appl. 1990, 528, 9–23. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Rutherfurd, S.M.; Gilani, G.S. Amino Acid Analysis. Curr. Protoc. Protein Sci. 2009, 58, 11.9.1–11.9.37. [Google Scholar] [CrossRef] [PubMed]
- Ullman, C.; Rhodes, M.A.; Davis, D.A. The effects of feed leaching on the growth of Pacific white shrimp Litopenaeus vannamei in a green-water tank system. Aquac. Res. 2019, 50, 3074–3077. [Google Scholar] [CrossRef]
- Watson, A.M.; Barrows, F.T.; Place, A.R. Leaching of taurine from commercial type aquaculture feeds. Aquac. Res. 2015, 46, 1510–1517. [Google Scholar] [CrossRef]
- Millamena, O.M.; Bautista-Teruel, M.N.; Kanazawa, A. Methionine requirement of juvenile tiger shrimp Penaeus monodon Fabricius. Aquaculture 1996, 143, 403–410. [Google Scholar] [CrossRef]
- Richard, L.; Blanc, P.-P.; Rigolet, V.; Kaushik, S.J.; Geurden, I. Maintenance and growth requirements for nitrogen, lysine and methionine and their utilisation efficiencies in juvenile black tiger shrimp, Penaeus monodon, using a factorial approach. Br. J. Nutr. 2010, 103, 984–995. [Google Scholar] [CrossRef] [Green Version]
- Millamena, O.M.; Bautista-Teruel, M.N.; Reyes, O.S.; Kanazawa, A. Requirements of juvenile marine shrimp, Penaeus monodon (Fabricius) for lysine and arginine. Aquaculture 1998, 164, 95–104. [Google Scholar] [CrossRef]
- Mente, E.; Coutteau, P.; Houlihan, M.; Davidson, I.; Sorgeloos, P. Protein turnover, amino acid profile and amino acid flux in juvenile shrimp Litopenaeus vannamei: Effects of dietary protein source. J. Exp. Biol. 2002, 205, 3107. [Google Scholar]
- Cruz-Ricque, L.; Guillaume, J.; Van Wormhoudt, A. Effect of squid extracts on time course appearance of glucose and free amino acids in haemolymph in Penaeus japonicus after feeding: Preliminary results. Aquaculture 1989, 76, 57–65. [Google Scholar] [CrossRef]
- Reddy, M.S.; Sailaja, M. Changes in hemolymph amino acid profiles during starvation in the penaeid prawn, Penaeus monodon. Proc. Indian Natl. Sci. Acad. Part B Biol. Sci. 1996, 62, 239–246. [Google Scholar]
- Kaushik, S.J.; Dabrowski, K.; Blanc, D. Nitrogen and energy utilization in juvenile carp (Cyprinus carpio) fed casein, amino acids or a protein-free diet. Reprod. Nutr. Dev. 1983, 23, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Murai, T.; Ogata, H.; Hirasawa, Y.; Akiyama, T.; Nose, T. Portal absorption and hepatic uptake of amino acids in rainbow trout force-fed complete diets containing casein or crystalline amino acids. Nippon Suisan Gakkaishi 1987, 53, 1847–1859. [Google Scholar] [CrossRef]
- Cowey, C.B.; Walton, M.J. Studies on the uptake of 14C-amino acids derived from both dietary 14C-protein and dietary 14C-amino acids by rainbow trout. J. Fish Biol. 1988, 33, 293–305. [Google Scholar] [CrossRef]
- Niu, J.; Lemme, A.; He, J.Y.; Li, H.Y.; Xie, S.W.; Liu, Y.J.; Yang, H.J.; Figueiredo-Silva, C.; Tian, L.X. Assessing the bioavailability of the Novel Met-Met product (AQUAVI® Met-Met) compared to DL-methionine (DL-Met) in white shrimp (Litopenaeus vannamei). Aquaculture 2018, 484, 322–332. [Google Scholar] [CrossRef]
- Alam, M.; Teshima, S.; Koshio, S.; Ishikawa, M. Effects of supplementation of coated crystalline amino acids on growth performance and body composition of juvenile kuruma shrimp Marsupenaeus japonicus. Aquac. Nutr. 2004, 10, 309–316. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| g kg−1 | FM | TM |
|---|---|---|
| Fishmeal | 500 | 0 |
| Poultry by-product meal | 0 | 200 |
| Full-fat soybean meal | 0 | 250 |
| Soy protein concentrate (SPC) | 0 | 120 |
| Gluten | 70 | 100 |
| Wheat flour | 395 | 262 |
| Fish oil | 15 | 25 |
| DL-Methionine (Met) | 0 | 10 |
| L-Lysine (Lys) | 0 | 10 |
| Taurine (Tau) | 0 | 3 |
| Soy lecithin | 10 | 10 |
| Cholesterol | 1 | 1 |
| Choline (60% choline chloride) | 5 | 5 |
| Vitamin premix | 2 | 2 |
| Vitamin C (stay C) | 1 | 1 |
| Yttrium oxide | 1 | 1 |
| Astaxanthin | 0.5 | 0.5 |
| Banox E | 0.2 | 0.2 |
| Feed Proximate Composition (on Dry Matter Basis, except for Moisture as Is) | ||
| Moisture (%) | 3.9 | 5.0 |
| Crude protein (CP) (%) | 49.8 | 51.8 |
| Gross energy (KJ g−1) | 21.0 | 21.3 |
| Total lipid (%) | 8.7 | 7.6 |
| Ash (%) | 8.3 | 6.2 |
| Carbohydrate (%) | 33.1 | 34.4 |
| Apparent Digestibility (%) | FM | TM |
|---|---|---|
| Dry matter | 63.2 ± 6.9 | 65.3 ± 4.0 |
| Crude protein | 77.7 ± 3.8 | 82.7 ± 0.1 |
| Gross energy | 82.9 ± 3.1 | 80.5 ± 2.1 |
| Total lipid | 77.2 ± 3.7 | 77.7 ± 2.9 |
| Diet | FM | TM | Stats | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Leaching Time (min) | 0 | 10 | 30 | 60 | 120 | 240 | 0 | 10 | 30 | 60 | 120 | 240 | Diet | Time |
| Water stability (% DM) | 100 | 98.8 | 97.3 | 96.6 | 94.6 | 93.3 | 100 | 97.0 | 95.6 | 95.1 | 90.2 | 87.4 | F = 28.5 *** | F = 33.9 *** |
| Interaction, F = 3.11 * | ||||||||||||||
| CP | 498 | 490 | 495 | 470 | 478 | 486 | 518 | 504 | 485 | 460 | 475 | 473 | ||
| Total AA | 398 | 408 | 403 | 453 | 404 | 413 | 459 | 472 | 481 | 447 | 474 | 459 | ||
| Arg | 22 | 23 | 23 | 25 | 21 | 22 | 30 | 27 | 30 | 29 | 31 | 28 | F = 21.99 *** | |
| His | 10 | 10 | 9 | 14 | 9 | 9 | 9 | 9 | 11 | 10 | 10 | 9 | ||
| Ile | 18 | 18 | 19 | 19 | 18 | 19 | 19 | 20 | 20 | 19 | 20 | 19 | ||
| Leu | 32 | 33 | 32 | 34 | 32 | 32 | 33 | 35 | 36 | 33 | 35 | 33 | ||
| Lys | 28 | 25 | 29 | 29 | 27 | 27 | 30 | 27 | 26 | 26 | 23 | 22 | ||
| Met | 11 | 10 | 10 | 10 | 9 | 10 | 14 | 11 | 10 | 9 | 8 | 7 | F = 3.05 * | |
| Phe | 19 | 20 | 20 | 21 | 19 | 20 | 23 | 24 | 25 | 22 | 25 | 24 | F = 26.8 *** | |
| Thr | 18 | 19 | 18 | 23 | 19 | 20 | 18 | 19 | 19 | 19 | 19 | 18 | ||
| Val | 15 | 16 | 15 | 17 | 15 | 16 | 16 | 17 | 17 | 16 | 17 | 16 | ||
| Sum EAA | 165 | 166 | 168 | 183 | 164 | 167 | 180 | 184 | 185 | 173 | 178 | 171 | ||
| Ala | 23 | 24 | 23 | 27 | 25 | 25 | 21 | 22 | 23 | 22 | 23 | 22 | F = 7.29 ** | |
| Asp | 35 | 36 | 36 | 42 | 36 | 37 | 41 | 43 | 42 | 40 | 43 | 41 | F = 5.49 ** | |
| Cys | 5 | 6 | 6 | 6 | 5 | 6 | 8 | 9 | 9 | 7 | 9 | 8 | F = 15.7 *** | |
| Glu | 83 | 87 | 84 | 86 | 85 | 85 | 102 | 109 | 110 | 101 | 110 | 104 | F = 25.6 *** | |
| Gly | 23 | 24 | 23 | 28 | 23 | 24 | 24 | 25 | 26 | 24 | 26 | 24 | ||
| Pro | 26 | 27 | 27 | 28 | 26 | 27 | 34 | 36 | 38 | 33 | 37 | 34 | F = 51.0 *** | |
| Ser | 18 | 19 | 18 | 33 | 22 | 25 | 23 | 24 | 25 | 23 | 25 | 25 | ||
| Tau | 3 | 2 | 2 | 2 | 1 | 1 | 4 | 2 | 2 | 2 | 1 | 1 | ||
| Tyr | 9 | 10 | 10 | 10 | 10 | 10 | 11 | 12 | 12 | 11 | 13 | 11 | ||
| Sum NEAA | 233 | 241 | 236 | 270 | 240 | 246 | 279 | 288 | 296 | 274 | 296 | 289 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simon, C.J.; Truong, H.; Habilay, N.; Hines, B. Feeding Behaviour and Bioavailability of Essential Amino Acids in Shrimp Penaeus monodon Fed Fresh and Leached Fishmeal and Fishmeal-Free Diets. Animals 2021, 11, 847. https://doi.org/10.3390/ani11030847
Simon CJ, Truong H, Habilay N, Hines B. Feeding Behaviour and Bioavailability of Essential Amino Acids in Shrimp Penaeus monodon Fed Fresh and Leached Fishmeal and Fishmeal-Free Diets. Animals. 2021; 11(3):847. https://doi.org/10.3390/ani11030847
Chicago/Turabian StyleSimon, Cedric J., Ha Truong, Natalie Habilay, and Barney Hines. 2021. "Feeding Behaviour and Bioavailability of Essential Amino Acids in Shrimp Penaeus monodon Fed Fresh and Leached Fishmeal and Fishmeal-Free Diets" Animals 11, no. 3: 847. https://doi.org/10.3390/ani11030847