1. Introduction
During the past few years, worldwide shrimp production has encountered limited development and production owing to, among other factors, viral and bacterial outbreaks [
1]. Many methods have been used to treat or prevent the development of new diseases, such as chemical and antibiotic compounds. However, the harmful potential effect of synthetic drugs on animals and the environment made these compounds problematic. Thus, to avoid their use, alternative methods have been studied, for instance, good handling practices of production and the enhancement of the immunological resistance of shrimps. The balance among host, pathogen and the environment is crucial for the animals that, even though they come into contact with the infection agent, do not develop the disease. Avoiding fluctuations in salinity and temperature are also essential practices in shrimp production, since they are stressful factors and can eventually serve as a stimulus for the development of an outbreak. On the other hand, the development of the immunocompetence of shrimps strengthens their immune system. This can be developed throughout the use of natural substances in diets, namely immunostimulants, such as some plant additives and seaweeds [
2,
3,
4,
5,
6].
Immunostimulant composts can be synthetically produced or obtained through natural sources, for instance through ingestion of microorganisms, such as fungi and bacteria, as well as algae, or even microalgae and seaweed [
7,
8]. Microalgae are already part of the food chain of marine shrimp. They are a source of long-chain fatty acid and are extremely important for penaeid shrimp, since their capacity to synthetize those fatty acids is limited [
9,
10,
11,
12,
13,
14]. Fatty acids are part of the group of lipids and are important because they are components of cell membranes and a source of energy. Studies with immunostimulants have demonstrated their positive effects on the growth and modulation of the immune system of Pacific white shrimp [
15,
16,
17].
Microalgae from the
Nannochloropsis genus are well known in aquaculture nutrition and play an important role in live feed enrichment because they are rich in lipids and contain high levels of eicosapentaenoic fatty acid. In addition, some species are also rich in carotenoids, such as
N. salina and
N. oculata [
18,
19,
20]. The benefits of
Nannochloropsis spp. in shrimp feeding are already well-studied, and they range from improving growth performance and survival to enhancing fatty acid content in shrimp flesh [
21,
22,
23]. These microalgae contain β 1-3 glycans in their cellulose cell wall. After analyzing some important immunological parameters for Pacific white shrimp, some studies have suggested that this cell wall component is responsible for shrimp immunostimulation [
24,
25,
26,
27,
28,
29,
30].
Immunological parameters such as hemograms, hemolymph coagulation time, phenoloxidase (PO) activity, the agglutination titer of plasma, plasma total protein concentration, and reactive oxygen species production (ROS), are used to verify the health of crustaceans. ROS is a common measurement of immunostimulation in shrimp [
24,
25,
26,
27,
28,
30,
31,
32,
33]. Among all ROS, superoxide anions are especially active against pathogens, owing to the occurrence of oxygen consumption, known as respiratory “burst”, during the production of these molecules, which inhibits growth and finally destroys the intruding pathogen [
1,
34].
This work aimed to evaluate Nannochloropsis spp. as a feed additive in the diet of Pacific white shrimp feeds for their effect on midgut microbiology, thermal shock resistance, and immune defense.
4. Discussion
Nannochloropsis spp. are microorganisms with the capacity to produce many beneficial compounds for animals. For instance, they have a high capacity to produce fatty acids (mainly those from the n-3 series), and they present immunostimulants, such as b-glucans, which can interact with the immune system, in either vertebrates or invertebrates [
29,
46]. The benefits of
Nannochloropsis spp. in shrimp were demonstrated in some works as increasing growth performance, survival and stimulation of the immune system. Thus, in the present work, the effects
Nannochloropsis spp. As a feed additive for Pacific white shrimp were assessed.
Digestibility of the microalgae meal was tested, and it was high. In general, protein digestibility, as well as that of some very important fatty acids, such as arachidonic, linoleic, linolenic and eicosapentaenoic, was respectively high. The digestibility of lipids and the eicosapentaenoic fatty acid was also high. This result is very important because it demonstrates that these shrimp can utilize nutrients present in the microalgae meal. Therefore, it can be concluded that microalgae meal is an ingredient highly assimilable by animals. Accordingly, it is likely that such connections are due to the fact that this feeding source is already part of the food chain of shrimp in the natural environment.
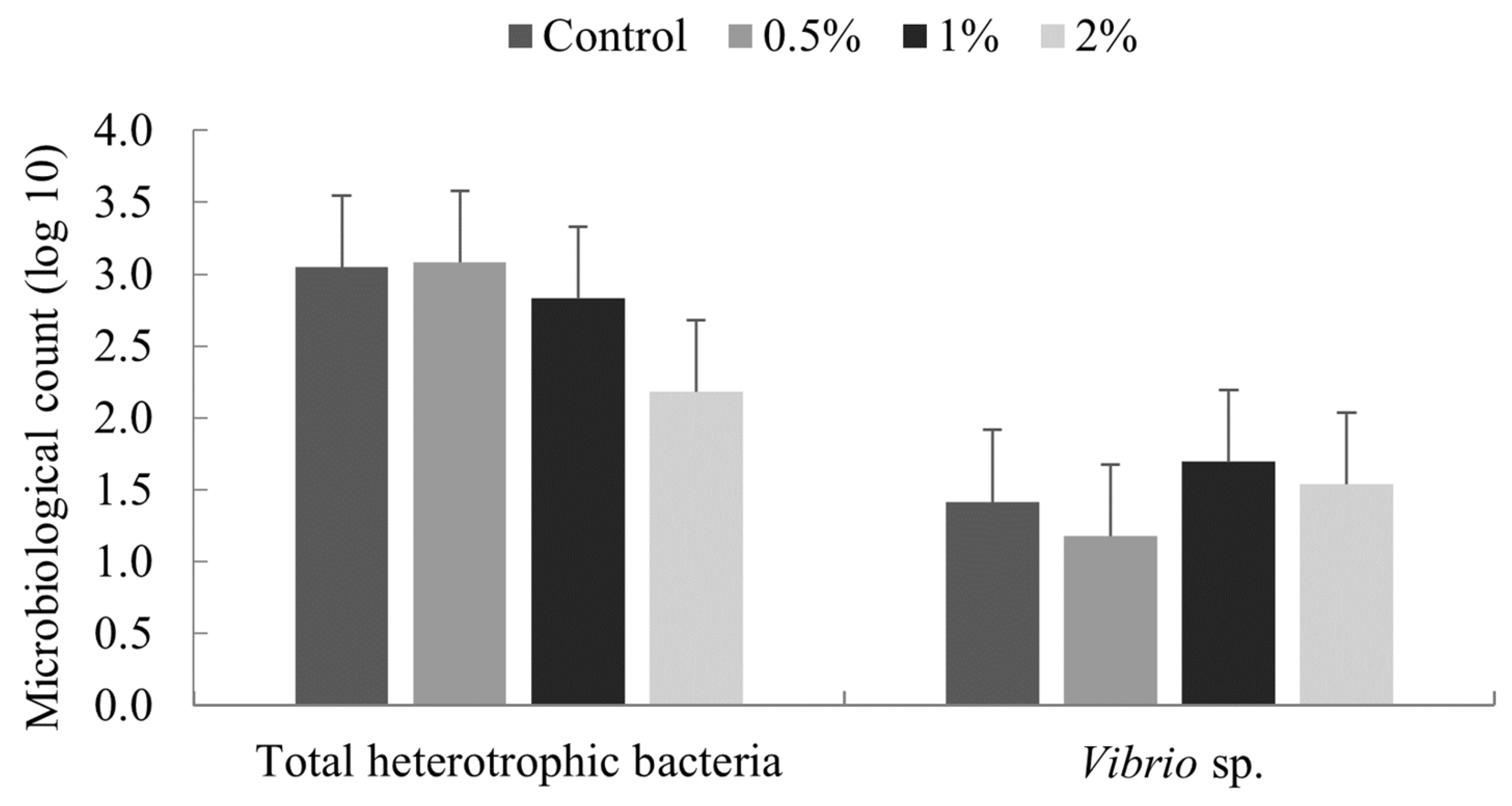
The addition of microalgae meal in diets did not interfere with bacterial midgut microbiology of the intestinal tract; nor did it alter some immunological parameters, such as total hemocyte counts, protein concentration, phenol-oxidase activity, and serum agglutination titer. However, statistical differences were found in the thermal shock challenge and reactive oxygen species (ROS) production.
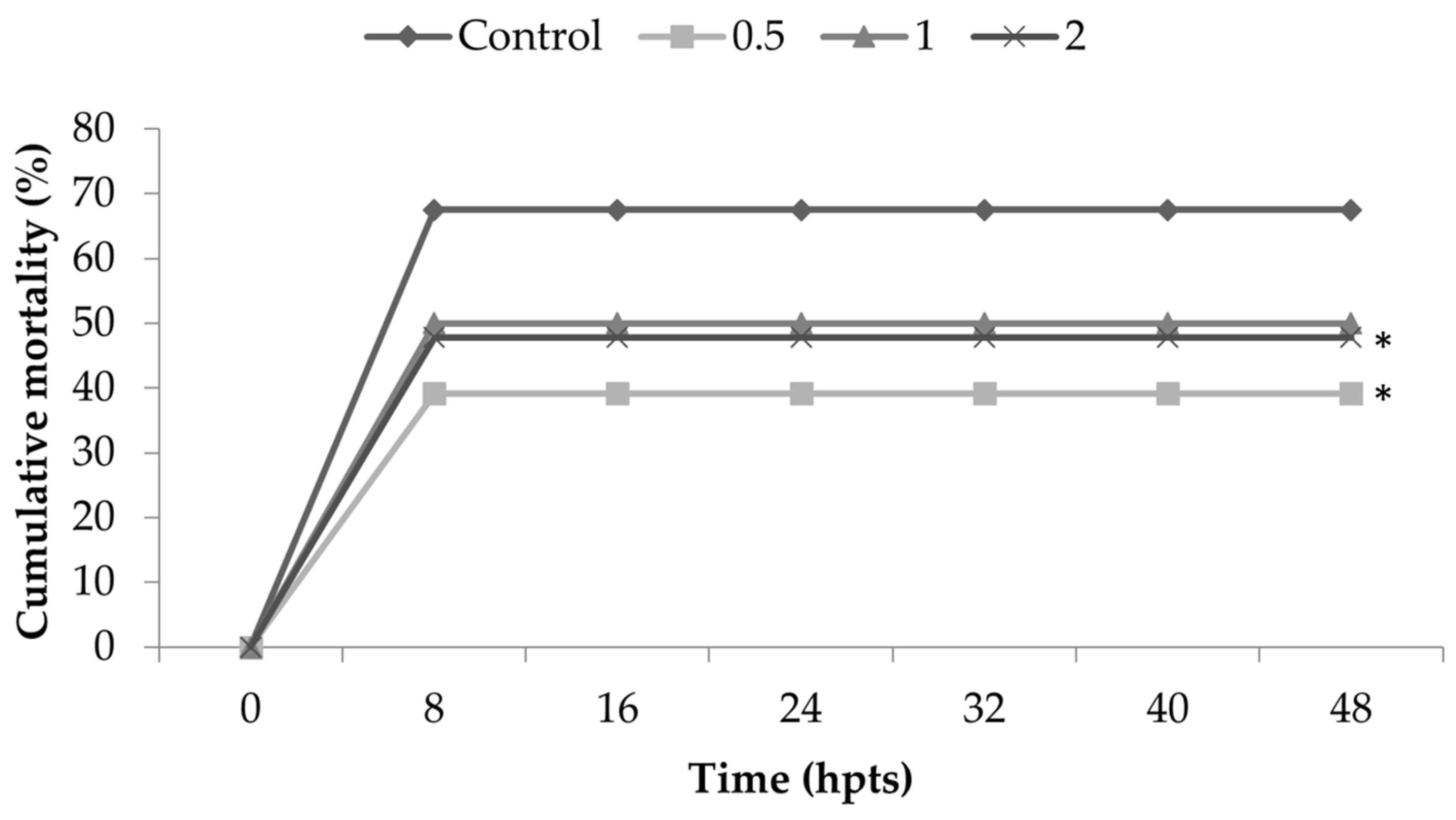
Thermal shock challenge provided the largest set of significant differences in the present study. The inclusion of
Nannochloropsis spp. in the diets of shrimp induced thermal shock resistance, as shown by the lower mortality of groups with 0.5% and 2% inclusion when compared to control groups. Even the treatment 1% did not show a significant difference from control; the mortality was lower, and the p value was 0.064—therefore, this was close to the significant difference. This finding has already been observed in shrimps after being fed with
Sargassum filipendula macroalgae [
47,
48], where resistance to thermal shock from shrimp was also observed, as in the present study, with also showed
Nannochloropsis spp. to be an immunostimulant which strengthened the immune system.
Marine shrimp are ectothermic organisms with no control over internal temperature; therefore, they are highly sensitive to temperature shifts in the rearing environment. Thus, low temperatures may lead to physiological changes, such as membrane fluidity loss, protein integrity loss, and oxidative stress [
49,
50]. Some studies have shown that some mechanisms of shrimp to regulate low temperature conditions, such as the regulation of lipid metabolism and modification of ionic transport, are impaired by the reduction in membrane fluidity, which is also very harmful to cellular functioning, as it leads to problems with cellular functioning and immobilizes transmembrane proteins [
49,
51,
52,
53]. Many biochemical mechanisms can increase the fluidity of the membrane, preventing its harmful effects. Among these mechanisms, it is known that the increase in fatty acids and cholesterol, unsaturated fatty acids and the restructuration their respective polar groups, as well as the elevation of long chain fatty acids, increase the fluidity of the membrane [
53]. It has been described in the literature that unsaturated fatty acids are incorporated in cell membrane phospholipids in situations of cold exposition, leading to modifications in membrane fluidity. This mechanism is common in animals that have resistance to thermal variations, such as insects, crustaceans, plants, and microorganisms [
49,
50,
53,
54,
55,
56]. Thus, the data of the present study demonstrate that the presence of unsaturated fatty acids of n-3 series in
Nannochloropsis spp. microalgae added in the diet could have been metabolized and helped to maintain membrane fluidity. The resistance to the thermal shock, as determined in this study, seems to prove this theory, as well as the high digestibility of fatty acids. Similar results were also demonstrated by Schleder et al. (2017) [
48] through MALDI-TOF MS analysis, where the inclusion of
Sargassum filipendula in the diet led to the increase in membrane fluidity and antimicrobial defense, also changing energetic and hemocyte lipid metabolism.
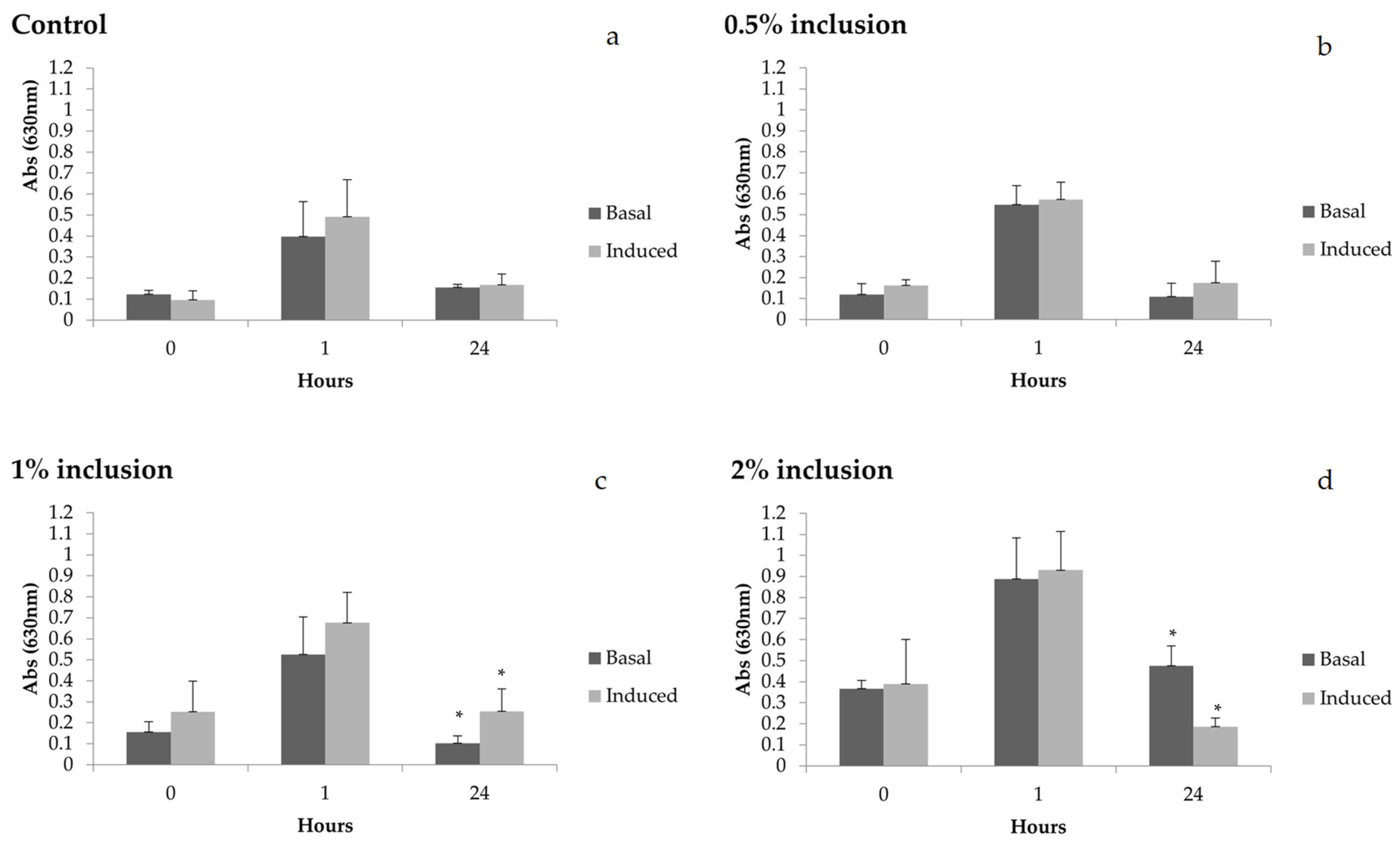
In the present work, changes in the immune status of animals could be measured by evaluating reactive oxygen species (ROS) in shrimp hemocytes. Before thermal shock, we saw an increase of about two-fold (
p ≤ 0.05) in the production of superoxide anion in the 2% treatment when compared to other groups. It is known that superoxide anion production is the first component produced in the ROS cascade during respiratory burst; therefore, this activation is more commonly observed during phagocytosis. Many studies suggest that b-glucans are immunostimulants and help in the prevention of disease in crustaceans [
24,
57], either by in vivo immune stimulation (for immersion and by feed additives) or in vitro, generating cellular responses through, for instance superoxide anions production [
24,
27]. In the present study, a crescent level of immunological stimulus was observed among treatments with 0, 0.5 and 1% of microalgae inclusion. On the other hand, animals fed 2% inclusion presented higher levels of immune stimulation, or ROS. This finding corroborates other studies demonstrating the increase in ROS in immunostimulated shrimp [
24], in shrimp infected by viruses, such as Taura syndrome [
58], white spot virus and hypodermal infection and hematopoietic necrosis [
25,
59].
Increased ROS production was also seen 1 h after thermal shock in all experimental groups. This increase could have been caused by hypoxia and reoxygenation that happens in cases of a severe reduction in metabolism during thermal shock and an abrupt return to normal temperature. Some studies have demonstrated that this situation promotes ROS production in the organism [
60,
61,
62], as well as instances of high oxygen saturation [
4]. At 24 h after thermal shock, ROS production was again similar to the that found before the challenge, demonstrating that the animals could return to basal levels. This is very important, because the presence of toxic molecules, such as ROS, in the absence of infection represents energetic overload for the organism and can lead to tissue damage [
63].
After laminarin stimulation, an increase of 2.5-fold in ROS production was observed in hemocytes in the group fed 1% of microalgae 24 h after thermal shock when compared to basal values. This result indicates that this microalgae concentration in the diet could enhance the capacity of hemocytes to produce ROS after stimulation. On the other hand, during the same 24 h period, the shrimp fed 2% microalgae presented a high basal production in which hemocytes decreased ROS production in response to laminarin by 2.6-fold. These results suggest that hemocytes of these animals were already at respiratory burst, even before in vitro induction. It is also possible that the cells of these animals were unable to generate ROS, indicating a possible low immunological competence of these animals during this period. Thus, even though shrimp fed 2% diets had higher ROS values at basal level, this was not accompanied by an increase in hemocyte capacity in producing ROS after laminarin stimulation, as evidenced in the group fed the 1% diet.
No statistical differences were found in some immune parameters, but it was still possible to observe a tendency of the increase in microalgae inclusion in diets and a further decrease at 2% inclusion. This fact suggests an immune stimulation of the animals that can be observed by the decrease in agglutination titer, which is caused by an alteration in stress and infection, as widely reported in the literature. These data corroborate the studies of Maggioni, Andreatta, Hermes, and Barracco (2004) [
45] who demonstrated the same decrease in agglutination titer after unilateral ablation in adult female
Litopenaeus vannamei. Furthermore, the decrease in coagulation capacity of the hemolymph and drop of total hemocyte counts could be associated with infection, stress or immunostimulation [
6,
24,
59]. Under such conditions, the hemocytes migrate and infiltrate the infected tissues [
64]. This event also happens in crustaceans infected with white spot syndrome virus and infectious myonecrosis virus [
2,
59,
65,
66].
It is important to highlight that phenol-oxidase (PO) activity increased in shrimp fed with 2% microalgae and began to decrease later in the experiment. This increase is important because the PO activity system acts in diverse defense situations and immune stimulation, such as phagocytosis, encapsulation and nodule formation which ends in melanized tissue [
28,
59]. This decrease in PO activity and protein concentration corroborates studies that demonstrate shrimp in stress conditions. The initial increase in PO and its following decrease suggest partial suppression of proPO that is likely caused by stress [
6,
27,
65].
The treatments where shrimp were fed diets of 0.5% and 1% inclusion seem to better prepare shrimp against an eventual infection and without much energetic cost, when compared to 2% inclusion. Some authors agree that the increase in total hemocyte count (CTH) provides better protection of shrimp against infections since hemocytes are mainly responsible for immune cellular reactions and are the site of many defense molecules’ expression in the organism. It also has better protection, with an increase in the capacity of hemolymph agglutination, regulation of ROS and the hemocyte capacity of production of ROS after laminarin stimulation [
67,
68,
69].
Researchers believe that having an immune system constantly activated is not beneficial for animals; however, strengthening the immune system, seeking higher immune competence, and, consequently, improving the capacity to fight off infection are all worth pursuing [
63]. Accordingly, a positive outcome of this study is the fact that feeding microalgae for only 15 days led to immune stimulation the shrimp. This result is important because, within a short period of time, it is possible to induce immunostimulation in shrimp though simple feeding and prepare them for possible stress events, such as weather changes, thus avoiding disease outbreaks.
Future studies might test the influence of microalgae inclusion on zootechnical parameters of shrimp and also challenge shrimp with bacteria of the genus Vibrio or white spot syndrome virus to assess if immune-stimulated shrimp respond better after infection.





{kind=link}
{kind=link}
{kind=link}