1. Introduction
Choline is a vitamin-like nutrient and is essential to the growth of some aquatic animals [
1,
2], and choline chloride is widely used as a source of dietary choline to determine the optimal level of dietary choline for various aquatic animals, such as
Sciaenops ocellatus [
1], hybrid tilapia (
Oreochromis niloticus ×
O. aureus) [
3],
Penaeus monodon [
2], and
Marsupenaeus japonicus [
4],
Cyprinus carpio var. Jian [
5]. Unfortunately, the information on the optimal requirement of dietary choline in crustacean species is fragmental and inconsistent, and the discrepancy may be related to animal species, growth stage, diet ingredient, culture condition, and evaluation criteria [
2,
4]. For example, the growth and survival of
M. japonicus (about 0.004 and 0.01 g initial body weight, respectively) were reportedly improved by feeding on 600 and 1200 mg/kg dietary choline [
6,
7]. However, Deshimaru and Kuroki (1979) had concluded that dietary choline chloride did not affect the growth of
M. japonicus with 0.5 g initial body weight [
8]. In juvenile
P. monodon, the optimum dietary choline requirement is affected by dietary lipid concentration [
9]. Thus, there is a need to accurately determine the choline requirement of the same species.
Dietary choline plays many fundamental functions as a structural component of phosphatidylcholine and acetylcholine, a precursor of the methionine and betaine, and a metabolic methyl donor [
7,
9,
10,
11]. Choline also has many metabolic and physiological functions in aquatic animals. Total lipids in the liver and plasma significantly decreased in
S. ocellatus fed a choline-free diet [
1]. In contrast, the hepatosomatic lipid concentration and hepatopancreatic index are higher in choline-deficient
P. monodon [
9] and
Epinephelus lanceolatus [
12]. The whole-body crude protein is also higher in
P. monodon fed given 4000 and 7000 mg/kg dietary choline [
2]. Several studies have revealed that choline can significantly affect serum biochemical contents such as triglyceride and cholesterol in aquatic animals [
1,
3,
12]. Moreover, dietary choline can regulate the antioxidant system to decrease oxidative damage in juvenile Jian carp [
5] and Nile tilapia [
13]. However, these studies have only focused on choline deficiency but ignored the effect of choline overdose on organism performance, and the molecular mechanisms of these physiological effects are not clear.
The Pacific white shrimp
Litopenaeus vannamei is one of the major penaeid species in aquaculture worldwide [
14]. The choline chloride requirement of
L. vannamei has been estimated without supplemental phospholipid or cholesterol, and the study is limited to the analyses of moisture and lipid content in shrimp [
15]. Dietary cholesterol is needed for optimal growth of
L. vannamei [
16], and the cholesterol-deficient diet can reduce shrimp growth and indirectly affect the optimal choline requirement of
L. vannamei. This study aims to compare the growth performance, whole-body proximate composition, serum metabolites, antioxidant, and digestive enzyme activities in the hepatopancreas of
L. vannamei fed a casein-based diet enriched with different levels of choline chloride. Based on metabolome and transcriptome analyses, we identified the key genes, metabolites, and pathways that are related to the molecular and metabolic mechanisms between shrimp fed choline-deficient and overdosed diets.
2. Materials and Methods
2.1. Experimental Diets
The isonitrogenous (about 42.20% crude protein) and isolipidic (about 7.99% crude lipid) semi-purified diets were formulated with six graded levels of choline chloride (0, 2000, 4000, 6000, 8000, and 12,000 mg/kg diet). Casein was the main protein source in the test diets. Fish oil, soybean oil, lecithin and cholesterol were the source of lipids. The experimental diets were formulated with a choline-free vitamin premix. A mixture of amino acids including betaine, L-glutamic, L-alanine acid, and glycine (Shanghai Sangon Biotech Co., Ltd., Shanghai, China) was used as an attractant [
2]. Butylated hydroxytoluene (BHT), an effective antioxidant, was added at a concentration of 0.005% to prevent lipid oxidation in the diets. After ground and sieved through a 60-mesh sieve, all dry ingredients were mixed thoroughly before oil was added. The choline chloride (reagent grade > 98%; Sangon Biotech, Shanghai, China) was supplemented to the basal diet while reducing the same amount of α-cellulose. Finally, about 300 mL/kg distilled water was added to the mixture to produce a stiff dough, which was wet-extruded into 2.5-mm diameter pellets at an extrusion temperature of less than 50 °C (CD4-1TS extruder, SCUT industrial factory, Guangzhou, China). The pellets were dried by blowing air at room temperature until reaching <10% moisture. Pellets were sieved by a 40-mesh sieve and stored at −20 °C. Ingredients and proximate composition of the basal diet are given in
Table 1. The animal ethics protocol was approved by the Hainan University Experimental Animal Ethics Committee (No. HNUAUCC-2020-00005).
The total choline in diets were detected by the Guangdong Institute of Analysis (Guangzhou, China) according to the national standards of China (GB/T 14924. 11-2001/3.7) using spectrophotometry. Briefly, the total choline in the diets was extracted by the alkali treatment process and purified by Florisil column chromatography, and then it reacted with Reinecke salt to form a pink choline—Reinecke salt complex. After eluting with acetone, this complex has an absorption maximum at 526 nm, and its absorption is proportional to the concentration of choline. The detection limit of choline was 0.1 mg, and the absolute value of the relative deviation of repeated detection results was ≤10%. The actual levels of total choline in the diets were 1082, 2822, 4770, 6450, 9120 and 13,100 mg/kg, respectively. Total choline is the sum of multiple choline compounds, and the choline detected in the basal diet may be choline, phosphatidylcholine, glycerophosphocholine, phosphocholine, and sphingomyelin in ingredients [
17,
18].
2.2. Growth Trial and Sampling
Juvenile L. vannamei were obtained from a shrimp hatchery in Wenchang, China. During a one-month acclimation process, the animals were maintained in an indoor cement pool and fed with a commercial diet (42% crude protein, 8% crude lipid) three times a day (0700, 1200, and 1800), and about three-quarters of the seawater in the pool was renewed once a day. A total of 720 healthy shrimp (1.75 ± 0.09 g) were randomly assigned to 24 cylindrical fiber-glass tanks filled with 100 L sand-filtered seawater. Each treatment was randomly assigned to four replicated tank with 30 shrimp each. Shrimp were fed three times a day at 0700, 1200, and 1800 with a daily ration of 5% body mass for eight weeks. When approximately 60% of seawater in tanks were daily exchanged, meantime the feces, uneaten feed, and exuviae were removed. The dead shrimp were removed, weighed, and recorded immediately. The natural illumination condition was used during the feeding trial and water quality was kept at salinity 30–33‰, temperature 28–30 °C, pH 7.8–8.2, dissolved oxygen 4.8–6.4 mg/L and ammonia-N < 0.05 mg/L.
All shrimp in each tank were fasted for 24 h and then anesthetized in an ice slurry. Shrimp were counted, weighed (0.01 g) and measured the total body length (0.1 cm) to calculate survival (%), condition factor (%), weight gain (%), and specific growth rate (% day−1). Three whole shrimp per tank were randomly selected and kept at −20 °C for the analysis of whole-body composition. Hemolymph samples were extracted from the pericardial cavity of remaining shrimp in each tank using 1 mL disposable sterilized syringes, stored in 1.5 mL Eppendorf tubes at 4 °C for 24 h, and then centrifuged at 4 °C, 5000 r/min for 10 min (3–18KS, Sigma, Osterode am Harz, Germany). The serum samples were stored in 200 μL Eppendorf tubes at −80 °C until biochemical analysis and metabolomic analysis. The hepatopancreas samples were taken from the aforesaid processed shrimp to calculate the hepatosomatic index (%) and were stored at −80 °C until subsequent analysis.
Survival, weight gain, specific growth rate, condition factor, and hepatosomatic index were evaluated using the following formulae:
- (1)
Survival (%) = 100% × final shrimp number/initial shrimp number.
- (2)
Weight gain (%) = 100% × (final weight (g) − initial weight (g))/initial weight (g).
- (3)
Specific growth rate (% day−1) = 100% × (ln (final weight) − ln (initial weight))/days.
- (4)
Condition factor (%) = 100% × final weight (g)/(body length (cm))3.
- (5)
Hepatosomatic index (%) = 100% × wet hepatopancreas weight (g)/wet body weight (g).
2.3. Proximate Composition Analysis
The proximate composition of experimental diets and shrimp whole body were analyzed following the standard methods of the Association of Official Analytical Chemists [
19]. Briefly, the moisture of diets and whole body were determined by drying to a constant weight at 105 °C (WFO-520, EYELA, Tokyo, Japan). The crude protein contents were measured by using the Dumas combustion method (Elementar rapid N exceed, Frankfurt, Germany). Crude lipid contents were determined via petroleum ether extraction using a Soxhlet extractor. The ash contents were determined using a muffle furnace (SX2-4-10N, Yiheng, Shanghai, China) at 550 °C for 8 h.
2.4. Biochemical Assay
The contents of total protein, total cholesterol, glucose, and triacylglycerol in serum samples were determined in eight replicates per diet. Eight hepatopancreases per treatment were weighed and homogenized in the pre-chilled 0.86% saline solution (1:10, w/v) at a frequency of 60 Hz at 4 °C for 30 s (Tissuelyser-24, Shanghai Jingxin Technology, Shanghai, China), centrifuged at 4 °C with 1500× g for 15 min (3-18KS, Sigma, Germany), and the supernatant was collected to measure total protein content, malondialdehyde (MDA) content, the activities of superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), α-amylase (AMS) and lipase (LPS). All the biochemical parameters were determined according to the manufacturer’s instructions using commercial assay kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
2.5. Untargeted Metabolomic Analysis
2.5.1. Sample Preparation
A total of 18 serum samples from shrimp fed 0, 6000, and 12,000 mg/kg choline chloride were obtained for the metabolomic analysis, and six biological replicates were used per group. Firstly, 400 µL of methanol-acetonitrile (1:1, v/v) was mixed with a serum sample (100 µL) and subjected to ultrasound extraction for 30 min (5 °C, 40 kHz). Then the solution was incubated at −20 °C for 30 min and centrifuged at 4 °C with 13,000× g for 15 min. Subsequently, the supernatant was dried under N2 (JXDC-20, Shanghai Jingxin Technology, Shanghai, China) and re-dissolved in 100 µL of acetonitrile-water (1:1, v/v). The re-dissolvent was ultrasonically extracted at 5 °C for 5 min and centrifuged for 5 min at 4 °C with 13,000× g. Finally, the supernatant was added to the sample bottle for detection. A mixture of equal quantities (20 µL per sample) was extracted from all samples and used as a quality control (QC) sample for liquid chromatography-mass spectrometry (LC-MS) analysis.
2.5.2. LC-MS Analysis
The LC-MS-based serum metabolic profiling adopted a Vanquish™ Horizon UHPLC system (Thermo Scientific, Germering, Germany) coupled with a Thermo Scientific™ Q Exactive™ Plus Hybrid Quadrupole-Orbitrap™ mass spectrometer. The parameters of chromatography were as follows: column: Ethylene Bridged Hybrid C18 (100 mm × 2.1 mm, 1.7 μm, Waters, Milford, Massachusetts, USA); gradient mobile phase: (A) deionized water containing 0.1% formic acid, (B) acetonitrile/isopropanol (1:1, v/v) mixture containing 0.1% formic acid; flow rate: 0.4 mL/min; sample injection volume: 2 μL; column temperature: 40 °C. The mobile phase gradient was: 0–3 min, A: 95–80%; 3–9 min, A: 80–5%; 9–13 min, A: 5–5%; 13–13.1 min, A: 5–95%; 13.1–16 min, A: 95–95%. The conditions of MS included the scan ranges (M/Z): 70–1050; Sheath gas flow rate (psi): 40; Aus gas flow rate (psi): 10; Aus gas heater temp (°C): 400; Normalized collision energy (V): 20–40–60; and IonSpray Voltage Floating (V): positive mode (ESI+), +3500; negative mode (ESI−), −2800. A QC sample was inserted for every 6 analytical samples during the experiment to evaluate the stability of the analytical system and assess the reliability of the results.
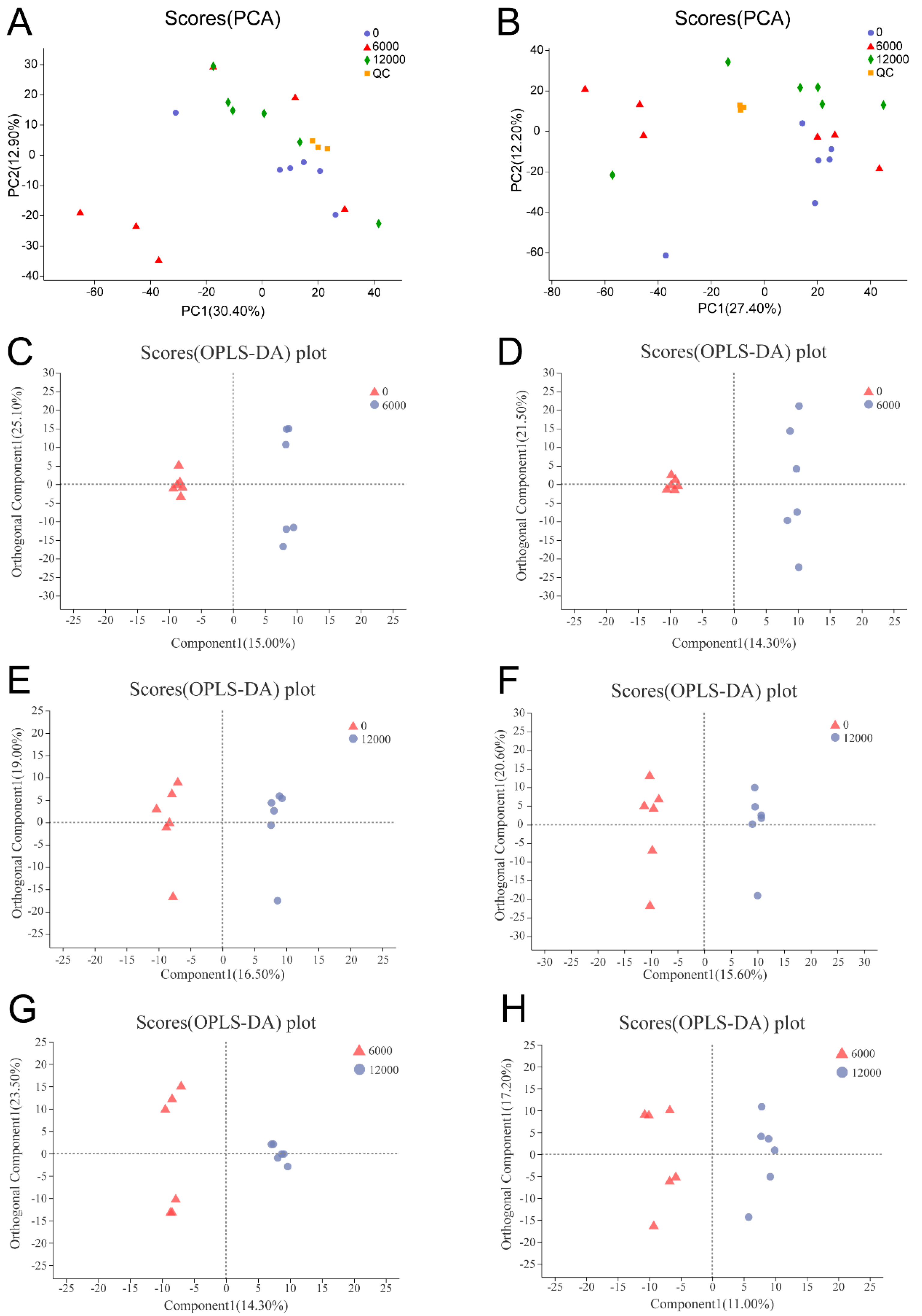
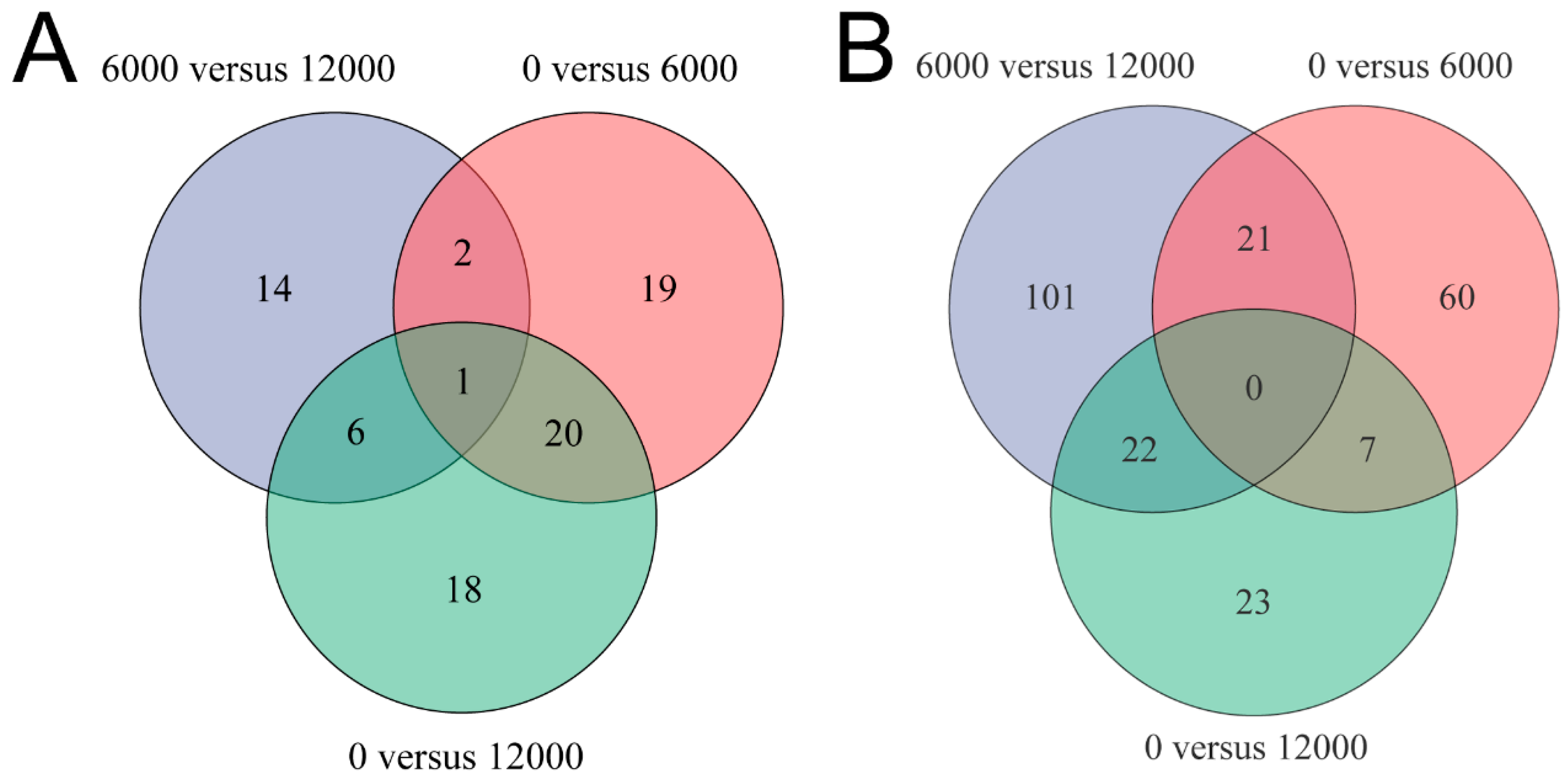
2.5.3. Metabolomic Data Analysis
The raw data obtained from the LC-MS analysis of all samples were processed initially using Progenesis QI software (Waters Corporation, Milford, MA, USA). comment HMDB database (
http://www.hmdb.ca/), METLIN database (
https://metlin.scripps.edu/), KEGG database (
https://www.genome.jp/kegg/), and a self-built database were selected for retrieval. Multivariate analyses including principal component analysis (PCA) and orthogonal partial least squares discrimination analysis (OPLS-DA) were performed by using ROPLS software (v1.6.2). Also, the OPLS-DA models were validated using a permutation test with 200 as the permutation number. To select differential metabolites and potential biomarkers, the variable importance in the projection (VIP, VIP > 1) values of metabolites in the OPLS-DA model and
p values (
p < 0.05) acquired from the
t-test analysis were regarded as the screening condition.
2.6. Transcriptomics Analysis
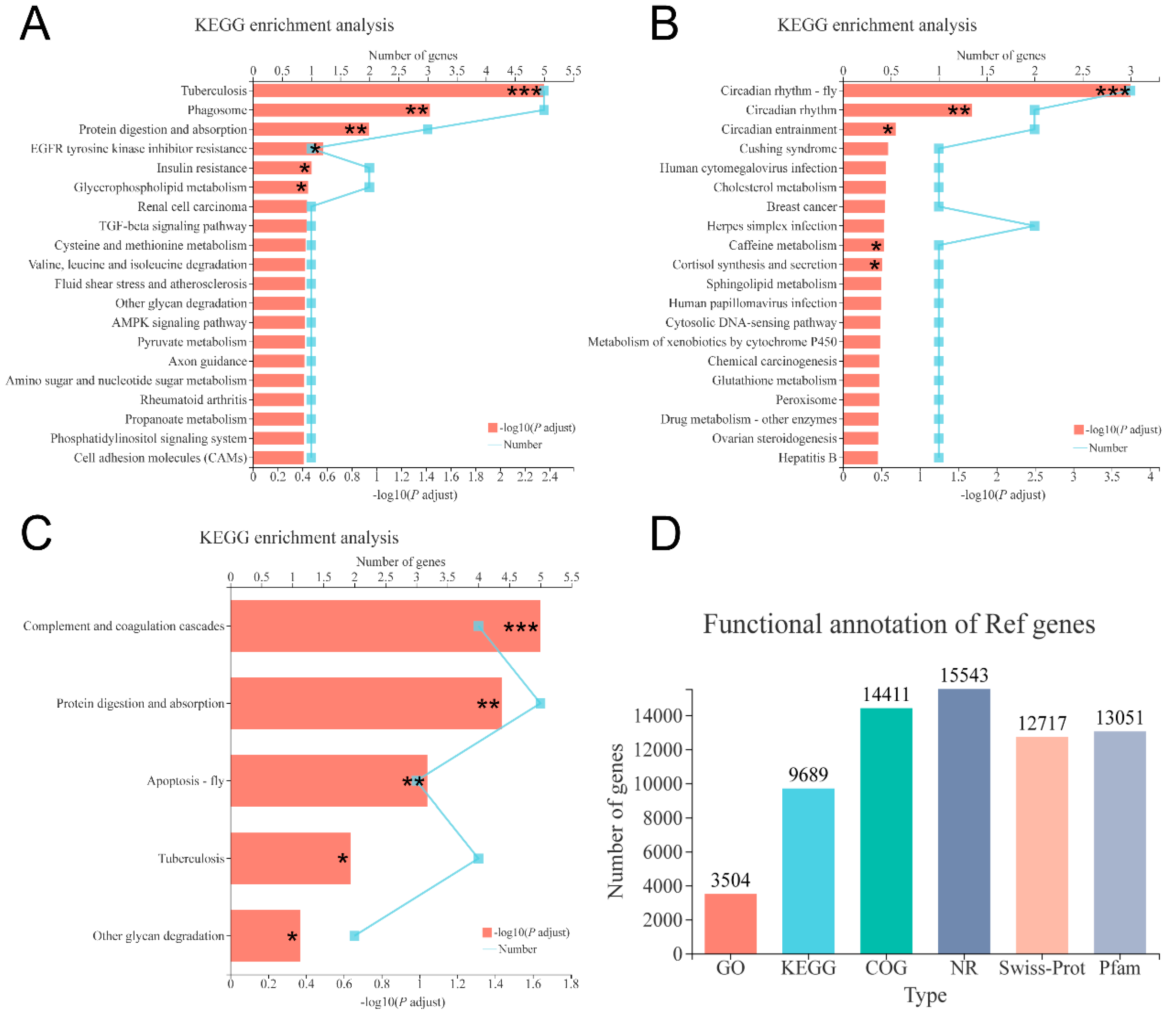
The total RNA of hepatopancreas samples dissected from individual shrimp fed 0, 6000, and 12,000 mg/kg choline chloride (3 shrimps per diet) was extracted using the TRIzol kit (Invitrogen, Carlsbad, CA, USA). The integrity, quality, and quantity of RNA were examined with an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) and a NanoDrop 2000 spectrophotometer (Thermo Scientific, Delaware, USA). Nine cDNA libraries were constructed using a TruSeqTM RNA sample prep kit (Illumina, San Diego, CA, USA) according to the manufacturer’s procedure. Nine libraries were sequenced using Illumina Novaseq 6000 sequencing platform (2 × 150 bp read length). The raw data were filtered with SeqPrep and Sickle software to obtain clean data. Then clean reads were mapped to the L. vannamei reference genome by using HISAT2 software and were assembled using Stringtie (v1.3.3b). The assembled transcriptome was annotated using the NR, Pfam, Swiss-Prot, COG, GO, and KEGG databases. Gene expression levels were calculated using RSEM software (v1.3.1) with the transcripts per million (TPM) method. Differentially expressed genes (DEGs) were analyzed using the DESeq2 software (v1.24.0), and the p-values were adjusted using the Benjamini-Hochberg method (P adjust). The selection criteria were p adjust < 0.05 and |log2 (fold change, FC)| ≥ 1.
2.7. Statistical Analysis
Statistical analysis was performed using SPSS 17.0 for Windows (SPSS Inc., New York, NY, USA). The outliers in each treatment were removed using box plot analysis. All the filtered data were first tested to confirm normal distribution and homogeneity of variance. One-way analysis of variance (ANOVA) was used to test the main effect of dietary manipulation using Tukey’s honestly significant difference (HSD) test as post hoc test [
20]. If both the raw data and log-transformed data showed a lot of heteroscedasticity, the analysis was performed using Welch’s ANOVA and Games-Howell post hoc test [
20]. All data were expressed as mean ± standard error (SE), and statistical significance was set at
p < 0.05.
4. Discussion
Choline has been considered as an essential vitamin for some fish species and other animals [
2,
5]; in this study, no significant differences in weight gain, survival, hepatosomatic index, and condition factor were found between different groups. It indicated that the total choline level of 1082 mg/kg in the basal diet was sufficient for the normal growth of the
L. vannamei. A previous study showed that the dietary choline requirement of
L. vannamei (0.72 ± 0.20 g) based on the instantaneous growth rate is 871 mg/kg, but choline requirement is not evident when lecithin is more than 1.5% of the diet [
15]. However, 3254.1 and 6488.3 mg/kg dietary choline were estimated to be the optimal choline requirements for juvenile
L. vannamei (initial weight of 0.30 ± 0.00 g) based on percent weight gain [
21]. The discrepancy of optimal choline requirements may be related to animal growth stage, diet ingredient, culture condition, and evaluation criteria [
2,
4]. Furthermore, it is generally believed that there may be interactions between dietary choline, betaine, methionine, and phosphatidylcholine, and these ingredients may influence the requirement for choline [
22]. Thus, more studies are needed in the future to determine the optimal choline requirement of
L. vannamei.
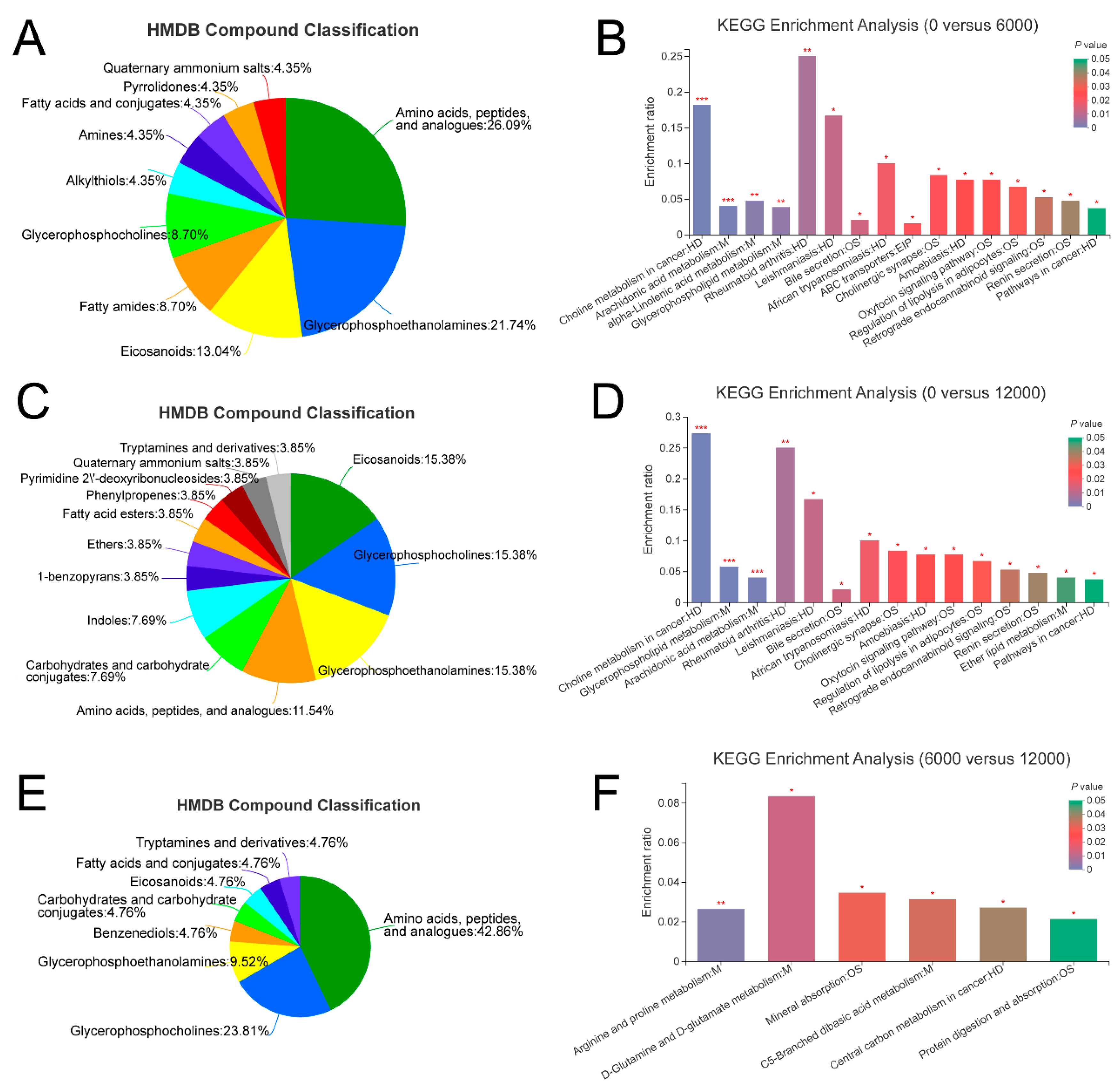
Trypsin as an important digestive protease mainly participates in food digestion, hydrolysis, and activation of zymogens [
23]. Compared with shrimp given 6000 mg/kg dietary choline chloride, the expression of trypsin-1-like was downregulated in the protein digestion and absorption pathway of shrimp given 12,000 mg/kg dietary choline. Thus, 12,000 mg/kg dietary choline might inhibit trypsin secretion, damages the protein utilization and deposition of
L. vannamei, thereby reducing the whole-body crude protein content. Paradoxically, the expressions of carboxypeptidase A1-like were upregulated in shrimp given 0 and 12,000 mg/kg dietary choline chloride. Carboxypeptidase A is a digestive carboxypeptidase and serves in the degradation of proteins in the digestive tract [
23]. Thus, the mechanisms of how dietary choline chloride regulated the whole-body crude protein of
L. vannamei need further study.
Overproduced reactive oxygen species (ROS) would lead to cell and tissue oxidative damage [
24]. Generally, MDA is a key metabolite production and a good biomarker of lipid peroxidation [
25]. This study showed that the MDA contents in shrimp hepatopancreas were decreased by dietary choline chloride, suggesting that choline supplement can decrease oxidative damage in the hepatopancreas of
L. vannamei. Similarly, higher MDA contents were found in the kidney, hepatopancreas and intestine of Jian carp fed the choline-deficient diet [
5,
26]. Choline deficiency induced rats fatty liver and lower levels of antioxidants, resulting in lipid peroxidation [
27]. In this study, the higher SOD and GSH-Px activities in choline-deficient shrimp may be related to the high ROS levels. ROS are rapidly eliminated by the antioxidant defense system [
28]. The SOD converts superoxide anion into hydrogen peroxide that passes freely through membranes and GSH-Px plays a vital role in removing hydrogen peroxide from cells [
29]. Furthermore, compared with the 12,000 mg/kg dietary choline chloride group, transcriptome results showed that glutathione S-transferase 1-like gene expression was significantly upregulated in the shrimp given 0 mg/kg dietary choline chloride. Glutathione S-transferases protect aquatic organisms from oxidative damage [
30]. However, scanty evidence is available concerning the relationship between choline and antioxidant response in
L. vannamei. As reported, choline may give a rise to antioxidant activity as decomposers of hydroperoxides [
31].
In
S. ocellatus, dietary choline induced higher concentrations of cholesterol, cholesterol esters, triglycerides, and phosphatidylcholine in plasma [
1]. Blood triglyceride, cholesterol, and phospholipid concentrations were higher in hybrid tilapia fed the diets supplemented with choline [
3]. Choline acts as a lipotropic factor preventing excessive lipid accumulation [
32]. In fish, choline deficiency can impair hepatic lipoprotein secretion, resulting in excessive accumulation of lipids in the liver [
12]. The hepatic secretion of lipoprotein requires active synthesis of phosphatidylcholine [
33]. Choline is a major component of phosphatidylcholine and sufficient phosphatidylcholine promotes lipid transport [
9,
12]. As a result, the contents of lipid metabolites in the blood increase when fish are supplied with dietary choline. Similarly,
P. monodon fed choline-deficient diets have higher hepatopancreatic lipid concentration than that of shrimp fed with supplemented choline chloride [
9]. On the contrary, higher concentrations of lipid metabolites in hemolymph, including high-density lipoprotein and low-density lipoprotein, were found in
L. vannamei fed the basal diet [
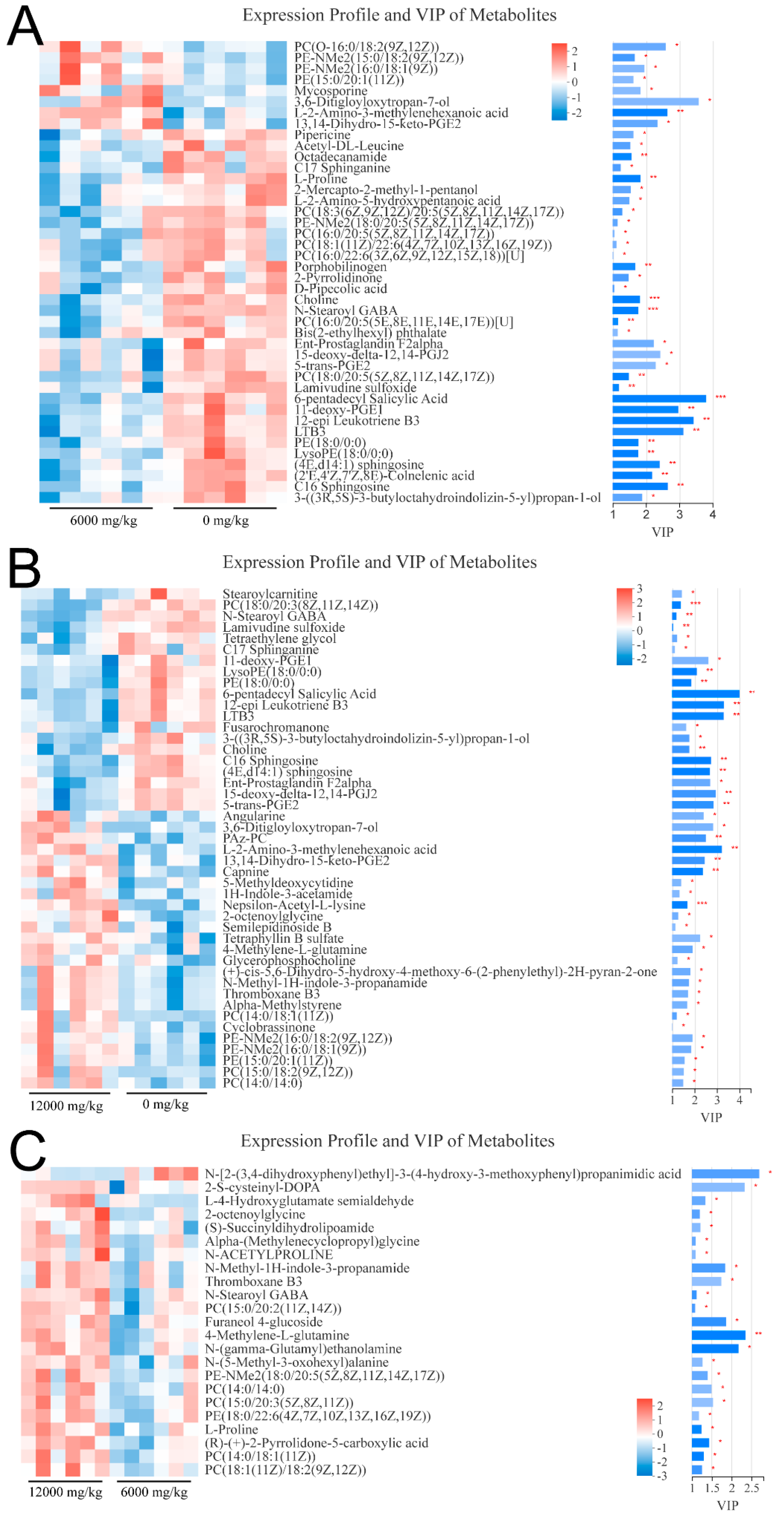
21]. In this study, compared with shrimp fed 0 mg/kg choline, phosphatidylcholine (lecithin) was downregulated in shrimp fed 6000 mg/kg choline (
Supplementary Figure S1B). Therefore, choline might reduce phosphatidylcholine content in
L. vannamei, resulting in a decrease in hemolymph lipid content. Moreover, dietary choline had no statistical effect on hemolymph cholesterol and triglyceride contents of
L. vannamei [
21]. In this study, no significant difference was found in serum triglyceride among all treatment groups, while the serum total cholesterol in shrimp fed 6000 mg/kg choline was significantly higher than that in shrimp fed 4000 mg/kg choline. Interestingly, no significant difference was found though the serum total cholesterol content of shrimp given 12,000 mg/kg choline was about 1.8 times higher than that of shrimp given 0 mg/kg choline. In hamsters and rabbits, plasma cholesterol levels are low in part by stimulating the production of mRNA for the low-density lipoprotein receptor in the liver [
34]. In this study, the gene expression of low-density lipoprotein receptor 1-like was upregulated in shrimp fed the basal diet. Thus, we postulate that dietary choline can increase serum cholesterol by impairing gene expression of low-density lipoprotein receptor.
Compared with shrimp given the basal diet, significantly lower serum choline was gained in shrimp fed 6000 mg/kg choline chloride (
Supplementary Table S1 and Figure S1B). Moreover, the results of the transcriptome showed that the expression of bile salt-activated lipase-like (EC: 3.1.1.7) was down-regulated, so the synthesis of choline from O-Acetylcholine was inhibited in shrimp fed 6000 mg/kg choline (
Supplementary Figure S1A). A possible explanation is that the decreased serum choline could help maintain the homeostasis of hemolymph osmotic pressure when cholesterol content increased, because choline could be metabolized to organic osmolytes such as phosphatidylcholine and betaine [
35].
Apolipoprotein D plays various physiological roles in an organism [
36,
37]. The overexpression of apolipoprotein D can protect
Drosophila against acute oxidative stress [
37]. The loss of mouse apolipoprotein D function can increase the sensitivity to oxidative stress and aggravate the level of brain lipid peroxidation [
38]. Particularly, arachidonic acid, a precursor for prostaglandin synthesis, is known to be mobilized from membranes upon oxidative stress, and apolipoprotein D can bind and mobilize arachidonic acid specifically for the synthesis of prostaglandin [
36]. Moreover, this binding may quench deleterious molecules such as lipid peroxides or scavenge other free radicals [
37]. In this study, the 5-trans-PGE
2 and 15-deoxy-delta-12,14-PGJ
2 in the arachidonic acid metabolism pathway increased in shrimp fed 0 mg/kg dietary choline. A previous study has shown that the 15-deoxy-delta-12,14-PGJ
2 reduced the cytotoxicity caused by hydrogen peroxide [
39]. Another vital oxidative stress-preventing role of 15-deoxy-delta-12,14-PGJ
2 is its ability to upregulate the expression of glutamylcysteine synthetase, a rate-limiting enzyme in glutathione synthesis, and induces the synthesis of glutathione [
40]. Therefore, the higher expression of apolipoprotein D and 15-deoxy-delta-12,14-PGJ
2 are the possible physiological adaptation for choline-deficient shrimp to lipid peroxidation. With the data available so far, it is difficult to clarify the physiological role of other prostaglandins such as 11-deoxy-PGE
1, 5-trans-PGE
2, 13,14-Dihydro-15-keto-PGE
2, and Ent-Prostaglandin F
2α in aquatic animals. Prostaglandins play a vital role in regulating many normal cellular functions [
41]. The synthesis of phosphatidylcholine also depends on prostaglandins, which modulates biosynthetic enzymes [
42]. The PGE
2 and PGF
2α can stimulate the formation of choline in a dose-dependent manner [
43,
44].

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}