
The Lack of Light-Dark and Feeding-Fasting Cycles Alters Temporal Events in the Goldfish (Carassius auratus) Stress Axis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Housing
2.2. Experimental Design
2.3. Locomotor Activity Recordings
2.4. Cortisol Plasmatic Levels
2.5. Gene Expression Analysis
2.6. Data Analysis
3. Results
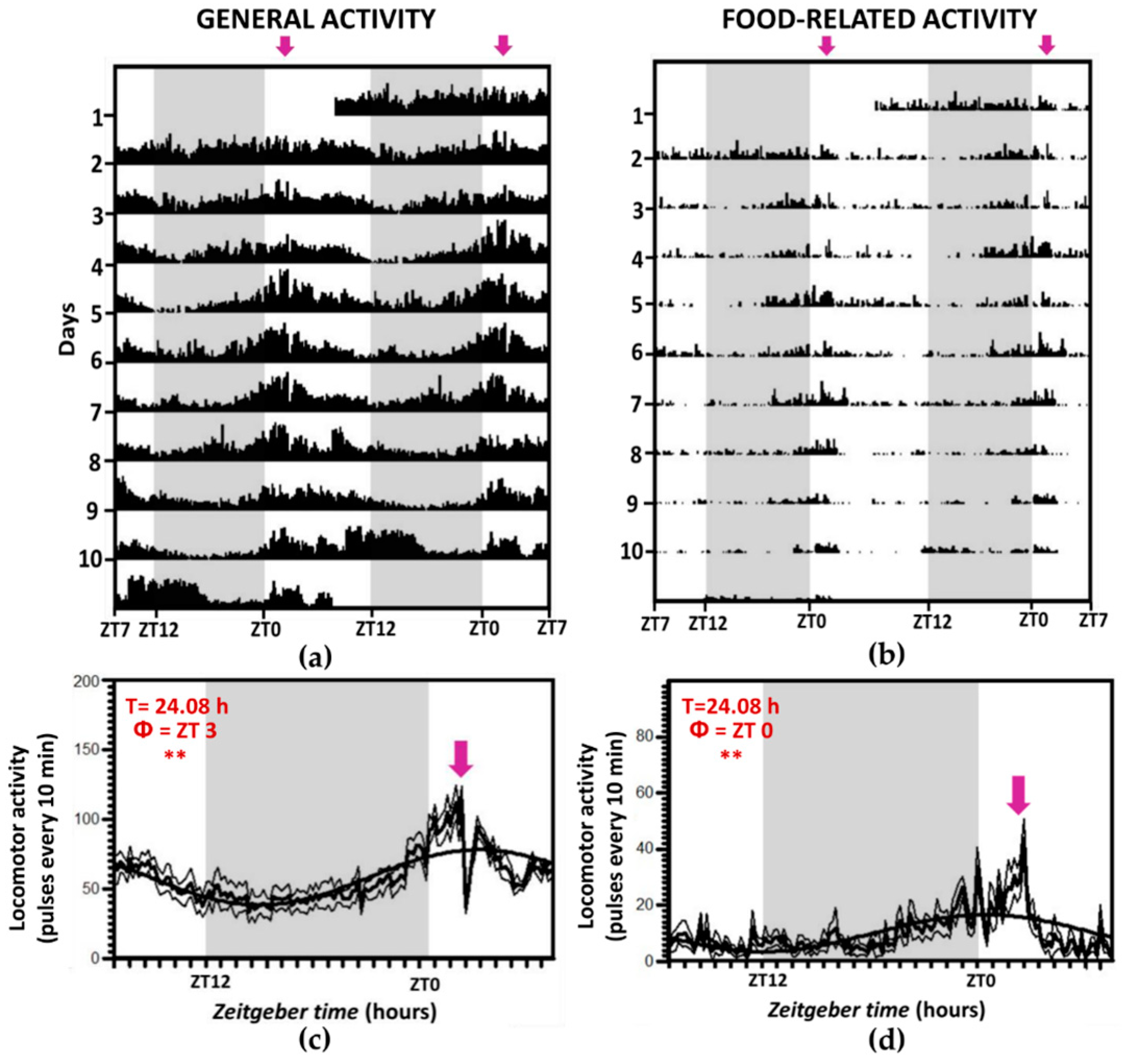
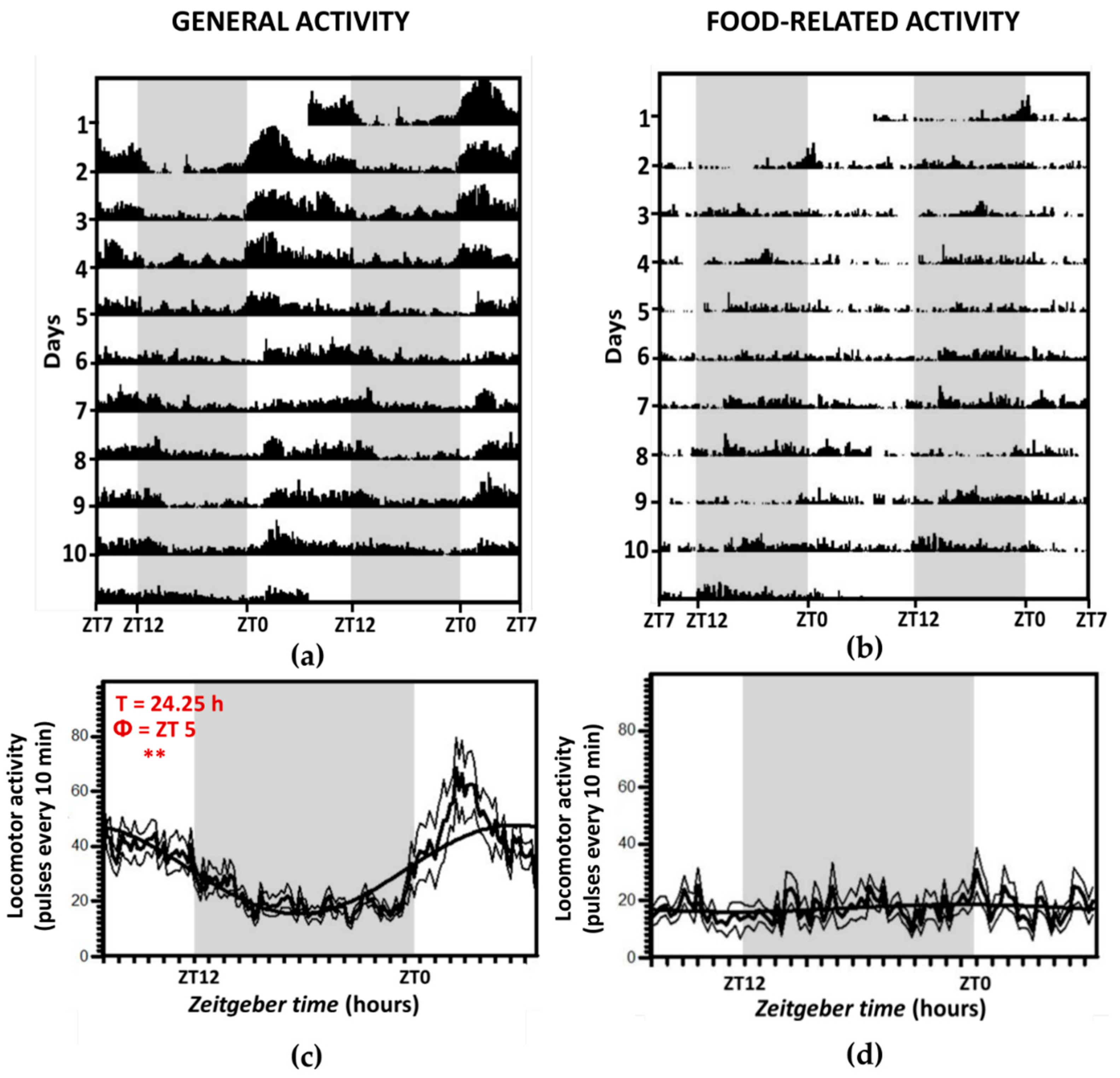
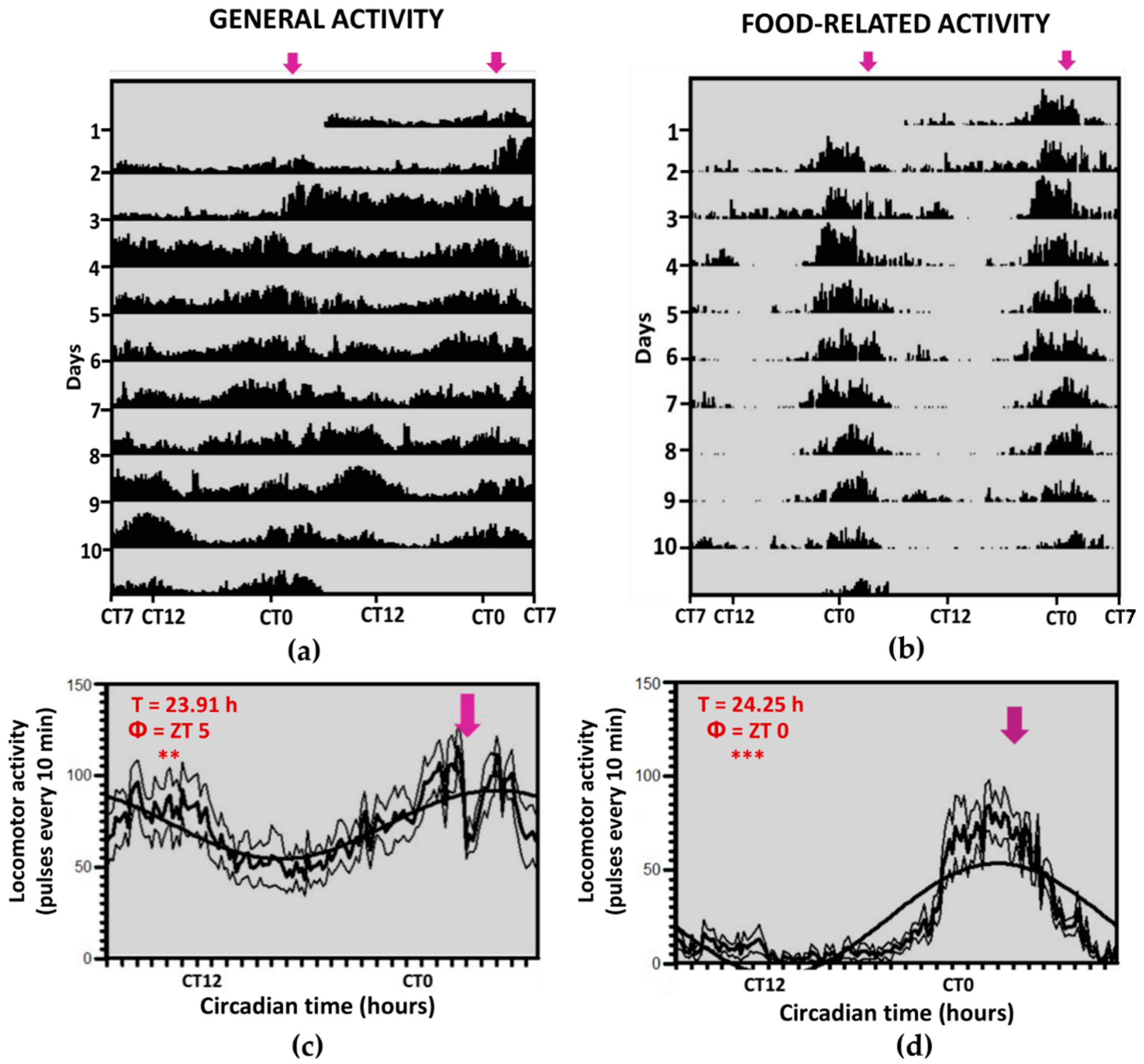
3.1. Locomotor Activity Recordings
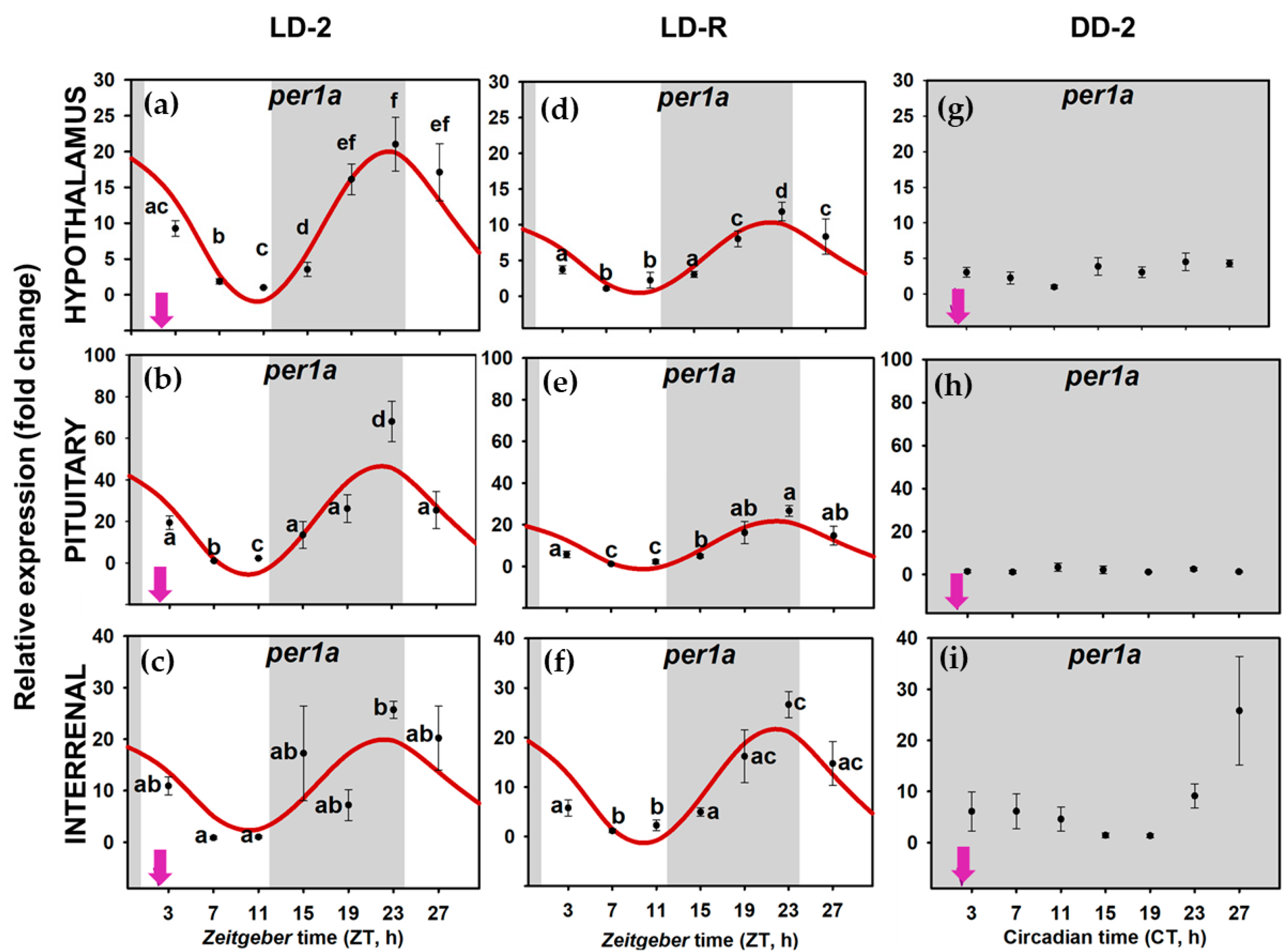
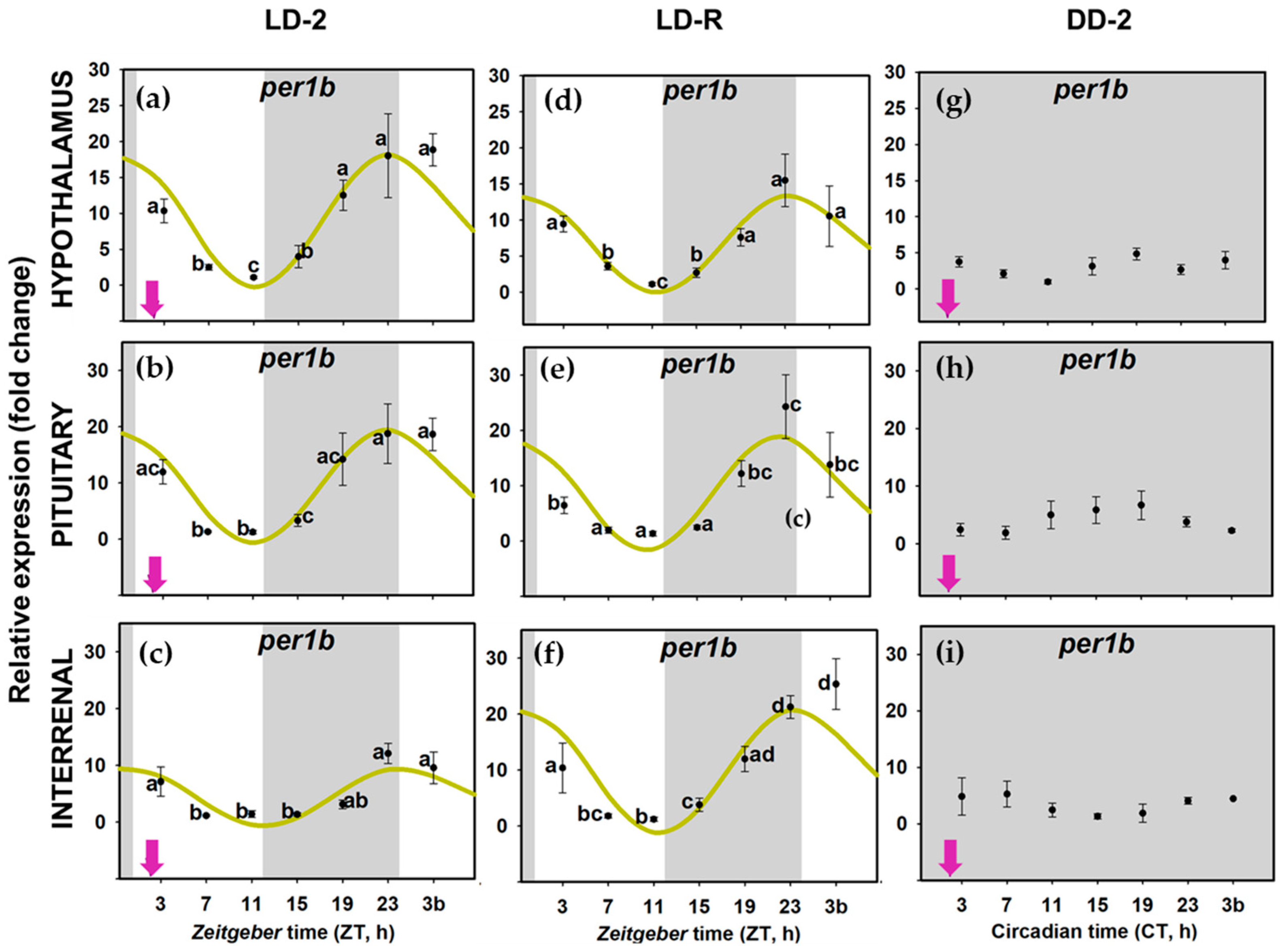
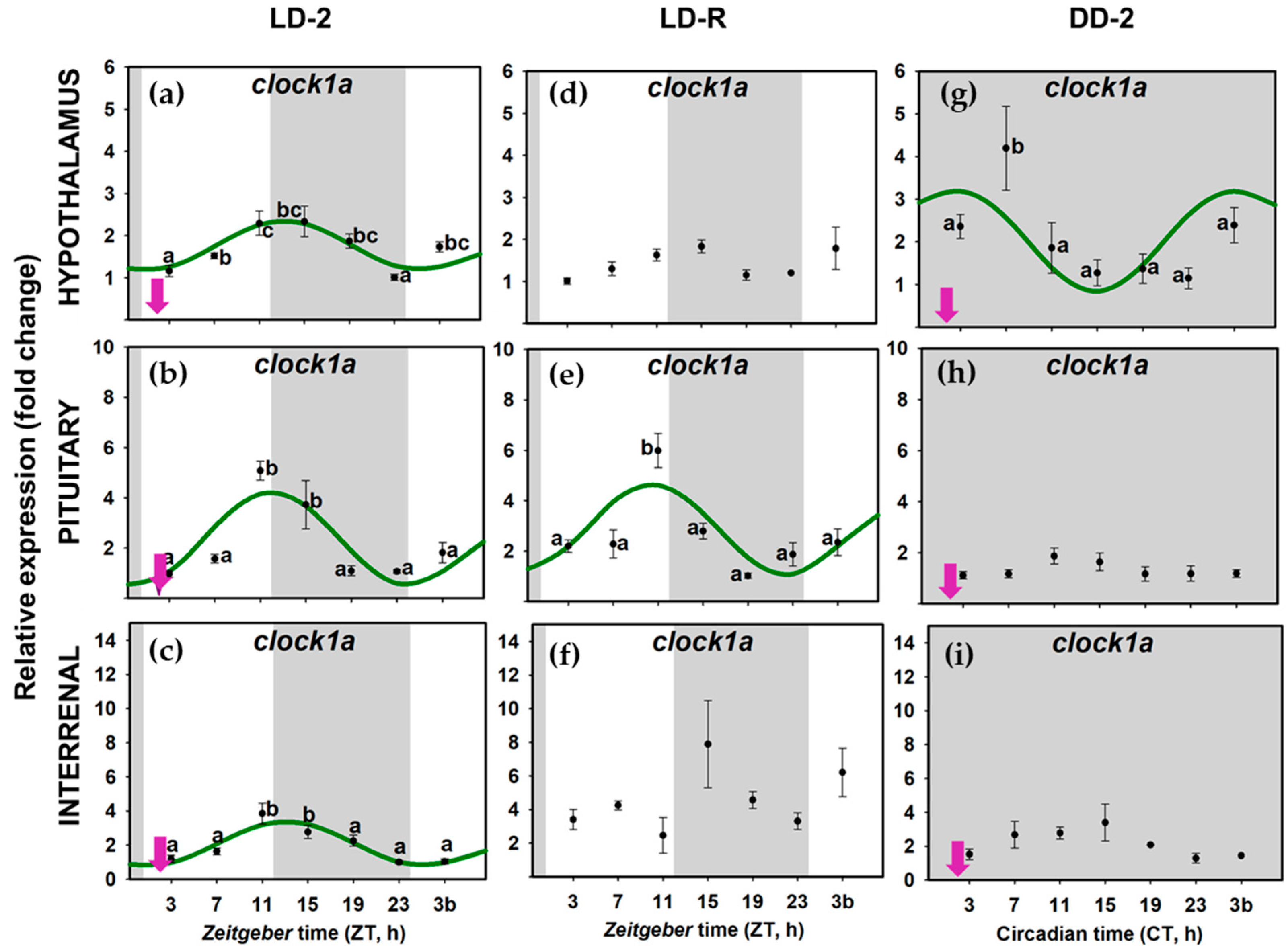
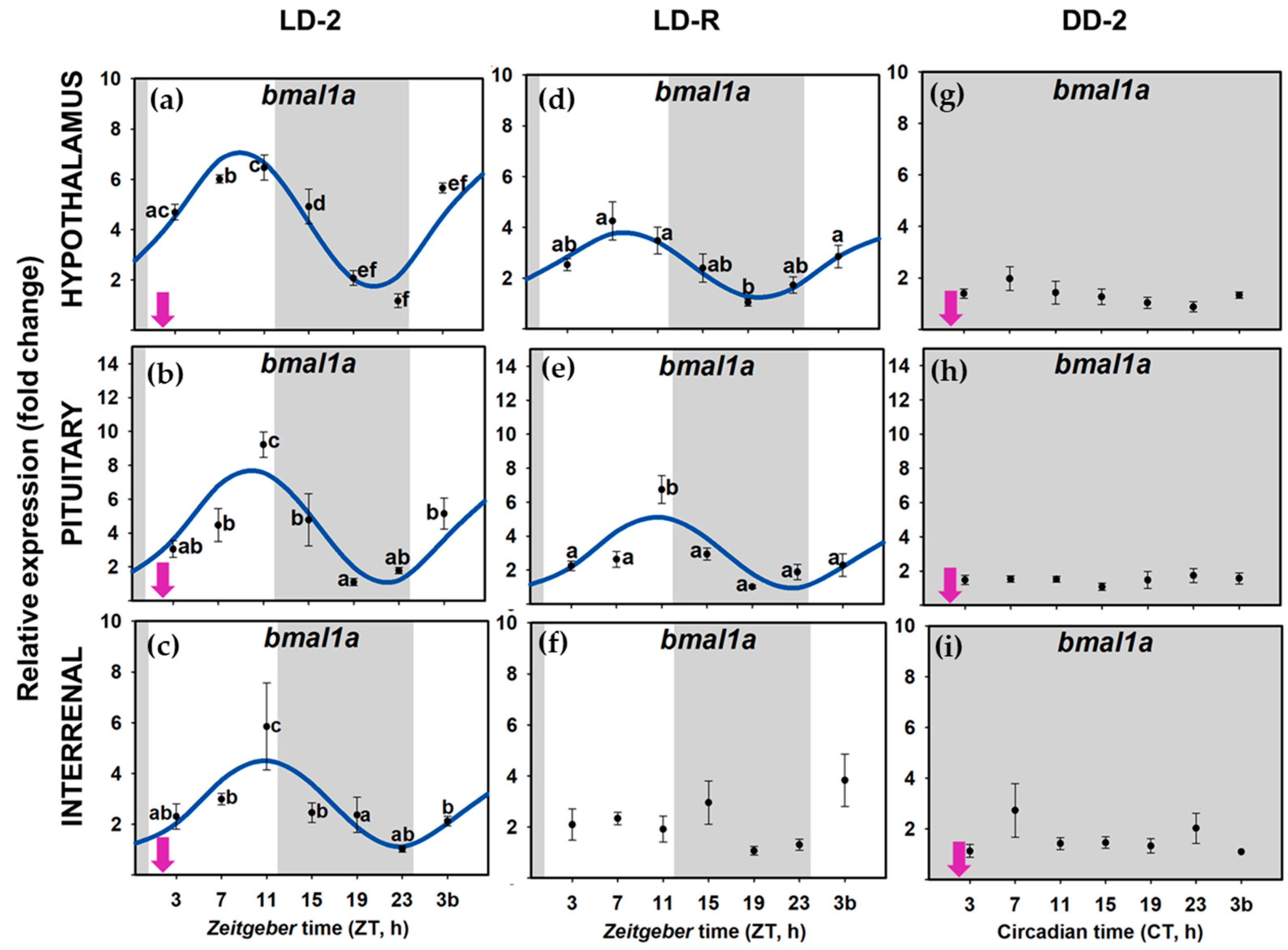
3.2. Daily Variations of Clock Gene Expression
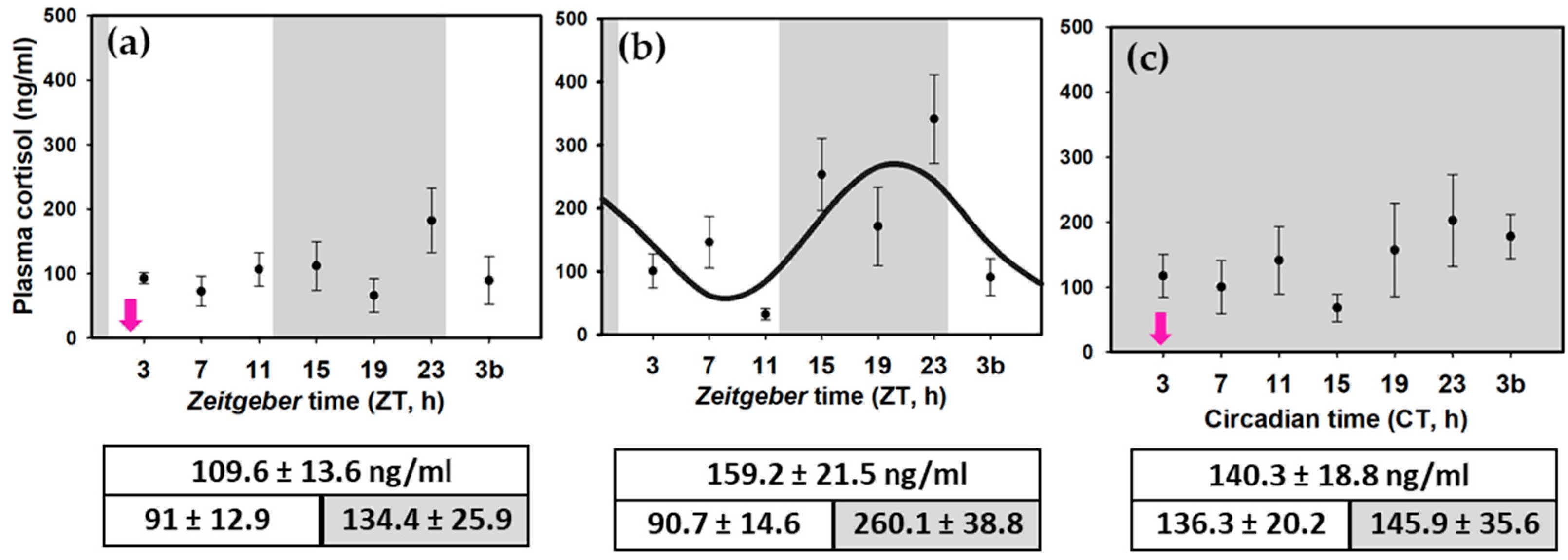
3.3. Daily Cortisol Variations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Buhr, E.D.; Takahashi, J.S. Molecular components of the mammalian circadian clock. Handb. Exp. Pharmacol. 2013, 217, 3–27. [Google Scholar]
- Cahill, G.M. Clock mechanisms in zebrafish. Cell Tissue Res. 2002, 309, 27–34. [Google Scholar] [CrossRef]
- Bell-Pedersen, D.; Cassone, V.M.; Earnest, D.J.; Golden, S.S.; Hardin, P.E.; Thomas, T.L.; Zoran, M.J. Circadian rhythms from multiple oscillators: Lessons from diverse organisms. Nat. Rev. Genet. 2005, 6, 544–556. [Google Scholar] [CrossRef]
- Albrecht, U. Timing to Perfection: The Biology of Central and Peripheral Circadian Clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [Green Version]
- Isorna, E.; de Pedro, N.; Valenciano, A.I.; Alonso-Gómez, Á.L.; Delgado, M.J. Interplay between the endocrine and circadian systems in fishes. J. Endocrinol. 2017, 232, R141–R159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.S.; Serrano, I.; Sánchez-Vázquez, F.J.; López-Olmeda, J.F. Circadian rhythms of clock gene expression in Nile tilapia (Oreochromis niloticus) central and peripheral tissues: Influence of different lighting and feeding conditions. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 775–785. [Google Scholar] [CrossRef]
- Gómez-Boronat, M.; Sáiz, N.; Delgado, M.J.; de Pedro, N.; Isorna, E. Time-Lag in Feeding Schedule Acts as a Stressor That Alters Circadian Oscillators in Goldfish. Front. Physiol. 2018, 9, 1749. [Google Scholar] [CrossRef]
- Damiola, F.; Le Minli, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Vázquez, F.J.; Aranda, A.; Madrid, J.A. Differential effects of meal size and food energy density on feeding entrainment in goldfish. J. Biol. Rhythms 2001, 16, 58–65. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Mendoza, J.; Challet, E. Brain clocks: From the suprachiasmatic nuclei to a cerebral network. Neuroscientist 2009, 15, 477–488. [Google Scholar] [CrossRef]
- Stephan, F.K. The “other” circadian system: Food as a Zeitgeber. J. Biol. Rhythms 2002, 17, 284–292. [Google Scholar] [CrossRef]
- Davidson, A.J. Search for the feeding-entrainable circadian oscillator: A complex proposition. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2006, 290. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, B.T.S.; Araujo, J.F. Food entrainment: Major and recent findings. Front. Behav. Neurosci. 2012, 6, 83. [Google Scholar] [CrossRef] [Green Version]
- Clarke, J.D.; Coleman, G.J. Persistent meal-associated rhythms in SCN-lesioned rats. Physiol. Behav. 1986, 36, 105–113. [Google Scholar] [CrossRef]
- López-Olmeda, J.F.; Tartaglione, E.V.; De La Iglesia, H.O.; Sánchez-Vázquez, F.J. Feeding entrainment of food-anticipatory activity and per1 expression in the brain and liver of zebrafish under different lighting and feeding conditions. Chronobiol. Int. 2010, 27, 1380–1400. [Google Scholar] [CrossRef]
- Aranda, A.; Madrid, J.A.; Sánchez-Vázquez, F.J. Influence of light on feeding anticipatory activity in goldfish. J. Biol. Rhythms 2001, 16, 50–57. [Google Scholar] [CrossRef]
- Feliciano, A.; Vivas, Y.; de Pedro, N.; Delgado, M.J.; Velarde, E.; Isorna, E. Feeding time synchronizes clock gene rhythmic expression in brain and liver of goldfish (Carassius auratus). J. Biol. Rhythms 2011, 26, 24–33. [Google Scholar] [CrossRef]
- Sánchez-Vázquez, F.J.; Madrid, J.A.; Zamora, S.; Tabata, M. Feeding entrainment of locomotor activity rhythms in the goldfish is mediated by a feeding-entrainable circadian oscillator. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1997, 181, 121–132. [Google Scholar]
- Schibler, U.; Gotic, I.; Saini, C.; Gos, P.; Curie, T.; Emmenegger, Y.; Sinturel, F.; Gosselin, P.; Gerber, A.; Fleury-Olela, F.; et al. Clock-talk: Interactions between central and peripheral circadian oscillators in mammals. Cold Spring Harb. Symp. Quant. Biol. 2016, 80, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dibner, C.; Schibler, U.; Albrecht, U. The Mammalian Circadian Timing System: Organization and Coordination of Central and Peripheral Clocks. Annu. Rev. Physiol. 2010, 72, 517–549. [Google Scholar] [CrossRef] [Green Version]
- Roenneberg, T.; Merrow, M. The network of time: Understanding the molecular circadian system. Curr. Biol. 2003, 13, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Vatine, G.; Vallone, D.; Gothilf, Y.; Foulkes, N.S. It’s time to swim! Zebrafish and the circadian clock. FEBS Lett. 2011, 585, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bretaño, A.; Callejo, M.; Montero, M.; Alonso-Gómez, Á.L.; Delgado, M.J.; Isorna, E. Performing a hepatic timing signal: Glucocorticoids induce gper1a and gper1b expression and repress gclock1a and gbmal1a in the liver of goldfish. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2016, 186, 73–82. [Google Scholar] [CrossRef]
- Whitmore, D.; Foulkes, N.S.; Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature 2000, 404, 87–91. [Google Scholar] [CrossRef]
- Blanco, A.M.; Sundarrajan, L.; Bertucci, J.I.; Unniappan, S. Why goldfish? Merits and challenges in employing goldfish as a model organism in comparative endocrinology research. Gen. Comp. Endocrinol. 2018, 257, 13–28. [Google Scholar] [CrossRef]
- Azpeleta, C.; Sánchez-Bretaño, A.; Isorna, E.; Nisembaum, L.G.; Velarde, E.; De Pedro, N.; Alonso-Gómez, A.L.; Delgado, M.J. Understanding the circadian system as a net of clocks: Daily expression of clock genes in the hypothalamus-pituitary-interrenal axis in Carassius auratus. Adv. Comp. Endocrinol. 2011, 9, 37–40. [Google Scholar]
- Nisembaum, L.G.; Velarde, E.; Tinoco, A.B.; Azpeleta, C.; De Pedro, N.; Alonso-Gómez, A.L.; Delgado, M.J.; Isorna, E. Light-dark cycle and feeding time differentially entrains the gut molecular clock of the goldfish (Carassius auratus). Chronobiol. Int. 2012, 29, 665–673. [Google Scholar] [CrossRef] [PubMed]
- Velarde, E.; Haque, R.; Iuvone, P.M.; Azpeleta, C.; Alonso-Gomez, A.L.; Delgado, M.J. Circadian clock genes of goldfish, carassius auratus: CDNA cloning and rhythmic expression of period and cryptochrome transcripts in retina, liver, and gut. J. Biol. Rhythms 2009, 24, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef] [PubMed]
- Bernier, N.J.; Peter, R.E. The hypothalamic-pituitary-interrenal axis and the control of food intake in teleost fish. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 639–644. [Google Scholar] [CrossRef]
- Grassi Milano, E.; Basari, F.; Chimenti, C. Adrenocortical and adrenomedullary homologs in eight species of adult and developing teleosts: Morphology, histology, and immunohistochemistry. Gen. Comp. Endocrinol. 1997, 108, 483–496. [Google Scholar] [CrossRef]
- To, T.T.; Hahner, S.; Nica, G.; Rohr, K.B.; Hammerschmidt, M.; Winkler, C.; Allolio, B. Pituitary-Interrenal Interaction in Zebrafish Interrenal Organ Development. Mol. Endocrinol. 2007, 21, 472–485. [Google Scholar] [CrossRef] [Green Version]
- Mommsen, T.P.; Vijayan, M.M.; Moon, T.W. Cortisol in teleosts: Dynamics, mechanisms of action, and metabolic regulation. Rev. Fish Biol. Fish. 1999, 9, 211–268. [Google Scholar] [CrossRef]
- Spiga, F.; Walker, J.J.; Terry, J.R.; Lightman, S.L. HPA axis-rhythms. Compr. Physiol. 2014, 4, 1273–1298. [Google Scholar]
- West, A.C.; Iversen, M.; Jørgensen, E.H.; Sandve, S.R.; Hazlerigg, D.G.; Wood, S.H. Diversified regulation of circadian clock gene expression following whole genome duplication. PLoS Genet. 2020, 16, e1009097. [Google Scholar] [CrossRef]
- Razzoli, M.; Karsten, C.; Yoder, J.M.; Bartolomucci, A.; Engeland, W.C. Chronic subordination stress phase advances adrenal and anterior pituitary clock gene rhythms. Am. J. Physiol. Integr. Comp. Physiol. 2014, 307, R198–R205. [Google Scholar] [CrossRef] [Green Version]
- Ishida, A.; Mutoh, T.; Ueyama, T.; Bando, H.; Masubuchi, S.; Nakahara, D.; Tsujimoto, G.; Okamura, H. Light activates the adrenal gland: Timing of gene expression and glucocorticoid release. Cell Metab. 2005, 2, 297–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girotti, M.; Weinberg, M.S.; Spencer, R.L. Diurnal expression of functional and clock-related genes throughout the rat HPA axis: System-wide shifts in response to a restricted feeding schedule. Am. J. Physiol. Metab. 2009, 296, E888–E897. [Google Scholar] [CrossRef] [PubMed]
- Kalsbeek, A.; van der Spek, R.; Lei, J.; Endert, E.; Buijs, R.M.; Fliers, E. Circadian rhythms in the hypothalamo-pituitary-adrenal (HPA) axis. Mol. Cell. Endocrinol. 2012, 349, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Azpeleta, C.; Martínez-Álvarez, R.M.; Delgado, M.J.; Isorna, E.; De Pedro, N. Melatonin reduces locomotor activity and circulating cortisol in goldfish. Horm. Behav. 2010, 323–329. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Duggleby, R.G. A nonlinear regression program for small computers. Anal. Biochem. 1981, 110, 9–18. [Google Scholar] [CrossRef]
- Delgado, M.J.; Aeonso-Gómez, A.L.; Gancedo, B.; de Pedro, N.; Valenciano, A.I.; Alonso-Bedate, M. Serotonin N-Acetyltransferase (NAT) Activity and Melatonin Levels in the Frog Retina Are Not Correlated during the Seasonal Cycle. Gen. Comp. Endocrinol. 1993, 92, 143–150. [Google Scholar] [CrossRef]
- Challet, E.; Mendoza, J.; Dardente, H.; Pévet, P. Neurogenetics of food anticipation. Eur. J. Neurosci. 2009, 30, 1676–1687. [Google Scholar] [CrossRef]
- Sánchez-Vázquez, F.J.; Madrid, J.A.; Zamora, S.; Iigo, M.; Tabata, M. Demand feeding and locomotor circadian rhythms in the goldfish, Carassius auratus: Dual and independent phasing. Physiol. Behav. 1996, 60, 665–674. [Google Scholar] [CrossRef]
- Vera, L.M.; De Pedro, N.; Gómez-Milán, E.; Delgado, M.J.; Sánchez-Muros, M.J.; Madrid, J.A.; Sánchez-Vázquez, F.J. Feeding entrainment of locomotor activity rhythms, digestive enzymes and neuroendocrine factors in goldfish. Physiol. Behav. 2007, 90, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Iigo, M.; Tabata, M. Circadian rhythms of locomotor activity in the goldfish Carassius auratus. Physiol. Behav. 1996, 60, 775–781. [Google Scholar] [CrossRef]
- López-Olmeda, J.F.; Madrid, J.A.; Sánchez-Vázquez, F.J. Melatonin effects on food intake and activity rhythms in two fish species with different activity patterns: Diurnal (goldfish) and nocturnal (tench). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2006, 144, 180–187. [Google Scholar] [CrossRef]
- Vera, L.M.; Negrini, P.; Zagatti, C.; Frigato, E.; Sánchez-Vázquez, F.J.; Bertolucci, C. Light and feeding entrainment of the molecular circadian clock in a marine teleost (Sparus aurata). Chronobiol. Int. 2013, 30, 649–661. [Google Scholar] [CrossRef]
- Sánchez, J.A.; López-Olmeda, J.F.; Blanco-Vives, B.; Sánchez-Vázquez, F.J. Effects of feeding schedule on locomotor activity rhythms and stress response in sea bream. Physiol. Behav. 2009, 98, 125–129. [Google Scholar] [CrossRef]
- Ceinos, R.M.; Chivite, M.; López-Patiño, M.A.; Naderi, F.; Soengas, J.L.; Foulkes, N.S.; Míguez, J.M. Differential circadian and light-driven rhythmicity of clock gene expression and behaviour in the turbot, Scophthalmus maximus. PLoS ONE 2019, 14, e0219153. [Google Scholar] [CrossRef]
- Cavallari, N.; Frigato, E.; Vallone, D.; Fröhlich, N.; Lopez-Olmeda, J.F.; Foà, A.; Berti, R.; Sánchez-Vázquez, F.J.; Bertolucci, C.; Foulkes, N.S. A Blind Circadian Clock in Cavefish Reveals that Opsins Mediate Peripheral Clock Photoreception. PLoS Biol. 2011, 9, e1001142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oike, H.; Oishi, K.; Kobori, M. Nutrients, Clock Genes, and Chrononutrition. Curr. Nutr. Rep. 2014, 3, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Oster, H.; Challet, E.; Ott, V.; Arvat, E.; de Kloet, E.R.; Dijk, D.J.; Lightman, S.; Vgontzas, A.; Van Cauter, E. The functional and clinical significance of the 24-h rhythm of circulating glucocorticoids. Endocr. Rev. 2017, 38, 3–45. [Google Scholar] [CrossRef]
- Tahara, Y.; Aoyama, S.; Shibata, S. The mammalian circadian clock and its entrainment by stress and exercise. J. Physiol. Sci. 2017, 67, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Helfrich-Förster, C.; Albrecht, U.; Pilorz, V.; Zhao, J.; Chen, L.; Xie, Y.; Tang, Q.; Chen, G.; Xie, M.; Yu, S. New Insights Into the Circadian Rhythm and Its Related Diseases. Front. Physiol. 2019, 10, 682. [Google Scholar]
- Montoya, A.; López-Olmeda, J.F.; Garayzar, A.B.S.; Sánchez-Vázquez, F.J. Synchronization of daily rhythms of locomotor activity and plasma glucose, cortisol and thyroid hormones to feeding in Gilthead seabream (Sparus aurata) under a light-dark cycle. Physiol. Behav. 2010, 101, 101–107. [Google Scholar] [CrossRef]
- Minnetti, M.; Hasenmajer, V.; Pofi, R.; Venneri, M.A.; Alexandraki, K.I.; Isidori, A.M. Fixing the broken clock in adrenal disorders: Focus on glucocorticoids and chronotherapy. J. Endocrinol. 2020, R13–R31. [Google Scholar] [CrossRef]
- Lazado, C.C.; Pedersen, P.B.; Nguyen, H.Q.; Lund, I. Rhythmicity and plasticity of digestive physiology in a euryhaline teleost fish, permit (Trachinotus falcatus). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2017, 212, 107–116. [Google Scholar] [CrossRef]
- Patiño, M.A.L.; Rodríguez-Illamola, A.; Conde-Sieira, M.; Soengas, J.L.; Míguez, J.M. Daily rhythmic expression patterns of clock1a, bmal1, and per1 genes in retina and hypothalamus of the rainbow trout, oncorhynchus mykiss. Chronobiol. Int. 2011, 28, 381–389. [Google Scholar] [CrossRef]
- Karolczak, M.; Burbach, G.J.; Sties, G.; Korf, H.-W.; Stehle, J.H. Clock gene mRNA and protein rhythms in the pineal gland of mice. Eur. J. Neurosci. 2004, 19, 3382–3388. [Google Scholar] [CrossRef] [PubMed]
- Asher, G.; Gatfield, D.; Stratmann, M.; Reinke, H.; Dibner, C.; Kreppel, F.; Mostoslavsky, R.; Alt, F.W.; Schibler, U. SIRT1 Regulates circadian clock gene expression through PER2 deacetylation. Cell 2008, 134, 317–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooley, J.J.; Schomer, A.; Saper, C.B. The dorsomedial hypothalamic nucleus is critical for the expression of food-entrainable circadian rhythms. Nat. Neurosci. 2006, 9, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Mieda, M.; Williams, S.C.; Richardson, J.A.; Tanaka, K.; Yanagisawa, M. The dorsomedial hypothalamic nucleus as a putative food-entrainable circadian pacemaker. Proc. Natl. Acad. Sci. USA 2006, 103, 12150–12155. [Google Scholar] [CrossRef] [Green Version]
- Fuller, P.M.; Lu, J.; Saper, C.B. Differential rescue of light- and food-entrainable circadian rhythms. Science 2008, 320, 1074–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, G.H.; Chung, S.; Choe, H.K.; Kim, H.-D.; Baik, S.-M.; Lee, H.; Lee, H.-W.; Choi, S.; Sun, W.; Kim, H.; et al. Adrenal peripheral clock controls the autonomous circadian rhythm of glucocorticoid by causing rhythmic steroid production. Proc. Natl. Acad. Sci. USA 2008, 105, 20970–20975. [Google Scholar] [CrossRef] [Green Version]
- Oster, H.; Damerow, S.; Kiessling, S.; Jakubcakova, V.; Abraham, D.; Tian, J.; Hoffmann, M.W.; Eichele, G. The circadian rhythm of glucocorticoids is regulated by a gating mechanism residing in the adrenal cortical clock. Cell Metab. 2006, 4, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Fahrenkrug, J.; Hannibal, J.; Georg, B. Diurnal rhythmicity of the canonical clock genes Per1, Per2 and Bmal1 in the rat adrenal gland is unaltered after hypophysectomy. J. Neuroendocrinol. 2008, 20, 323–329. [Google Scholar] [CrossRef]
- Meier, A.H. Daily variation in concentration of plasma corticosteroid in hypophysectomized rats. Endocrinology 1976, 98, 1475–1479. [Google Scholar] [CrossRef]
- Torres-Farfan, C.; Abarzua-Catalan, L.; Valenzuela, F.J.; Mendez, N.; Richter, H.G.; Valenzuela, G.J.; Serón-Ferré, M. Cryptochrome 2 expression level is critical for adrenocorticotropin stimulation of cortisol production in the capuchin monkey adrenal. Endocrinology 2009, 150, 2717–2722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richter, H.G.; Torres-Farfan, C.; Garcia-Sesnich, J.; Abarzua-Catalan, L.; Henriquez, M.G.; Alvarez-Felmer, M.; Gaete, F.; Rehren, G.E.; Seron-Ferre, M. Rhythmic expression of functional MT1 melatonin receptors in the rat adrenal gland. Endocrinology 2008, 149, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Storch, K.F.; Weitz, C.J. Daily rhythms of food-anticipatory behavioral activity do not require the known circadian clock. Proc. Natl. Acad. Sci. USA 2009, 106, 6808–6813. [Google Scholar] [CrossRef] [PubMed] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saiz, N.; Gómez-Boronat, M.; De Pedro, N.; Delgado, M.J.; Isorna, E. The Lack of Light-Dark and Feeding-Fasting Cycles Alters Temporal Events in the Goldfish (Carassius auratus) Stress Axis. Animals 2021, 11, 669. https://doi.org/10.3390/ani11030669
Saiz N, Gómez-Boronat M, De Pedro N, Delgado MJ, Isorna E. The Lack of Light-Dark and Feeding-Fasting Cycles Alters Temporal Events in the Goldfish (Carassius auratus) Stress Axis. Animals. 2021; 11(3):669. https://doi.org/10.3390/ani11030669
Chicago/Turabian StyleSaiz, Nuria, Miguel Gómez-Boronat, Nuria De Pedro, María Jesús Delgado, and Esther Isorna. 2021. "The Lack of Light-Dark and Feeding-Fasting Cycles Alters Temporal Events in the Goldfish (Carassius auratus) Stress Axis" Animals 11, no. 3: 669. https://doi.org/10.3390/ani11030669