Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Culture and Sampling
2.2. Experimental Food Making
2.3. Plasma Analysis
2.4. Tissue Analysis
2.5. Statistical Analysis
3. Results
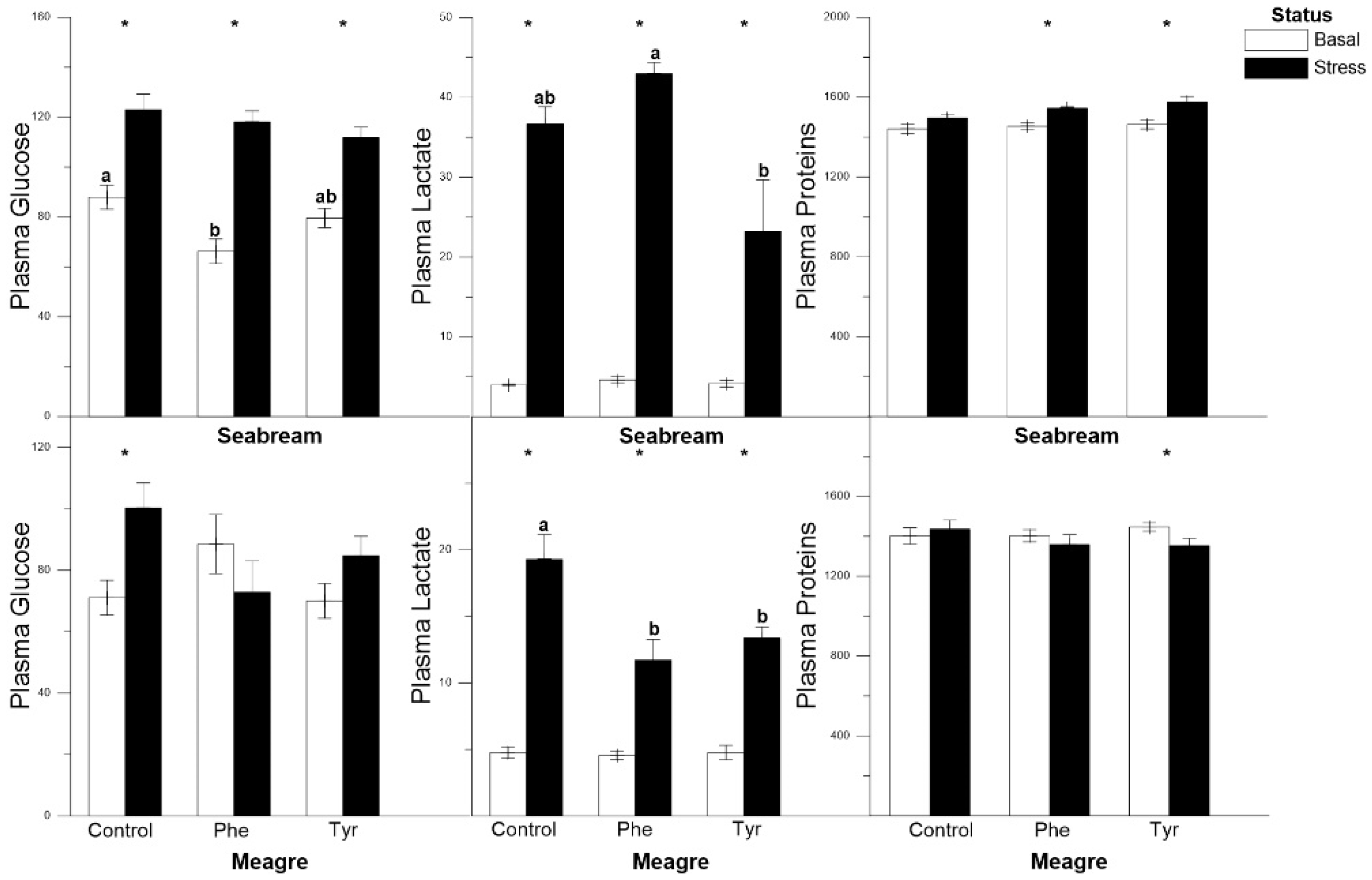
3.1. Metabolite Analysis
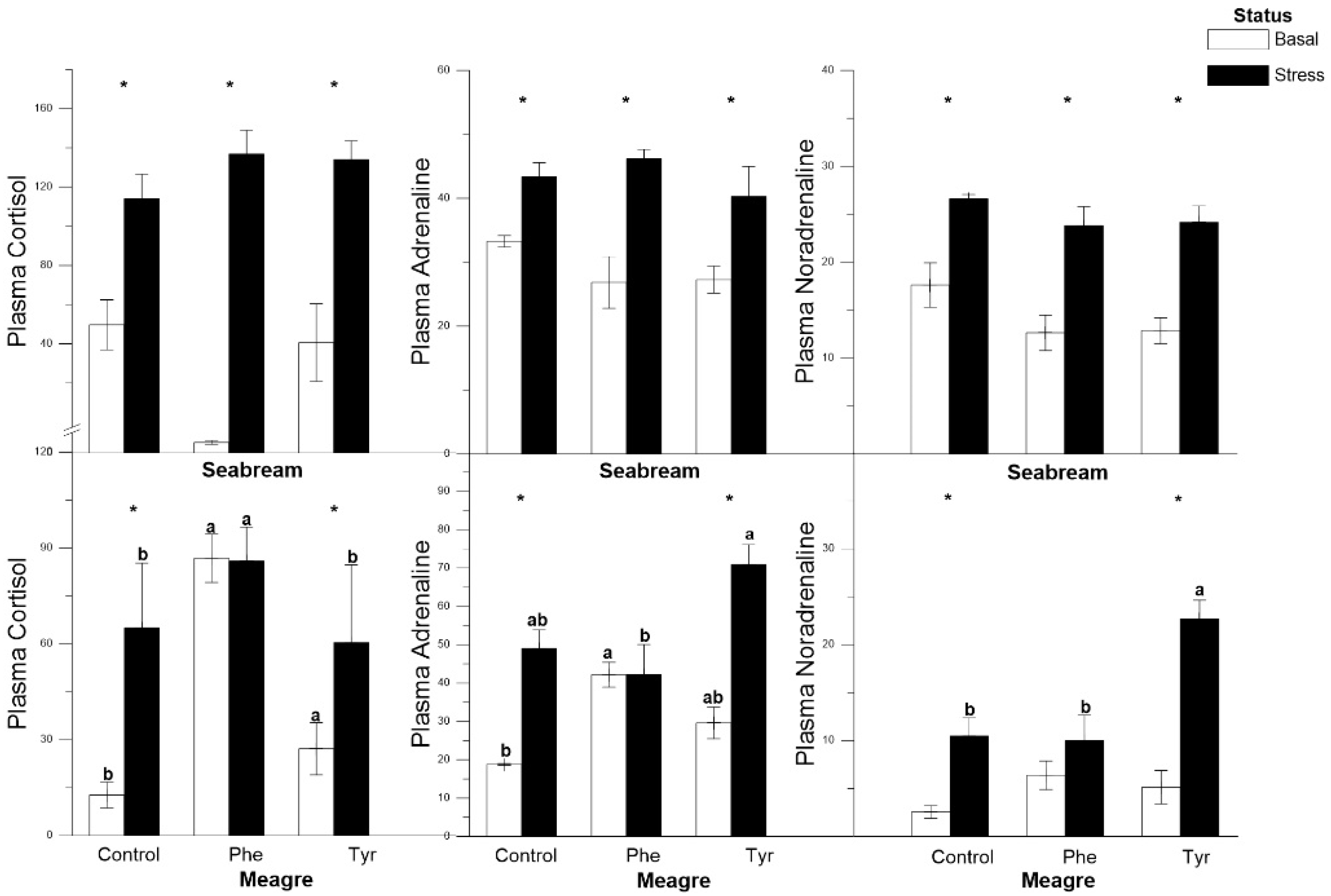
3.2. Plasma Cortisol and Catecholamines
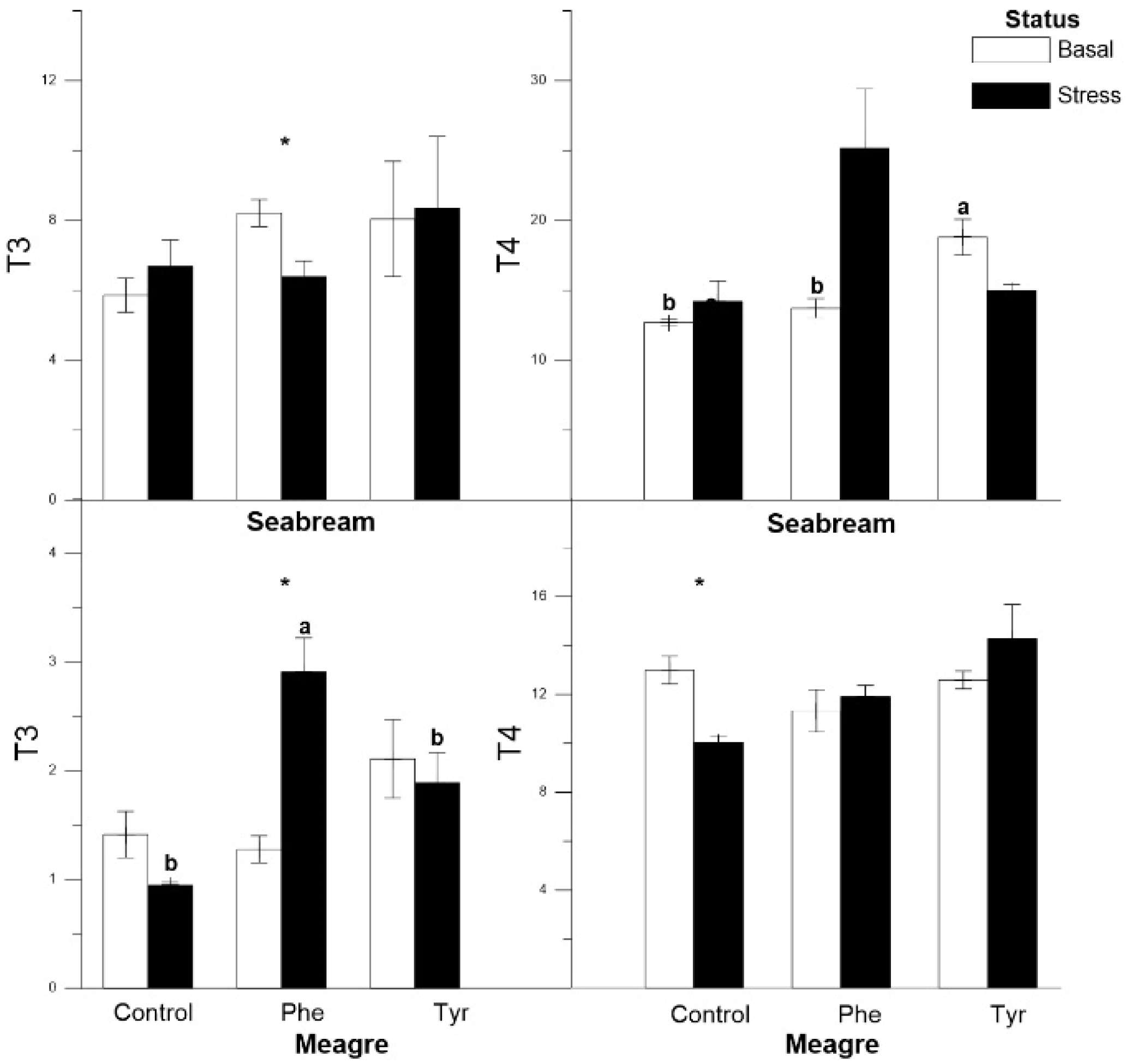
3.3. Thyroid Hormones
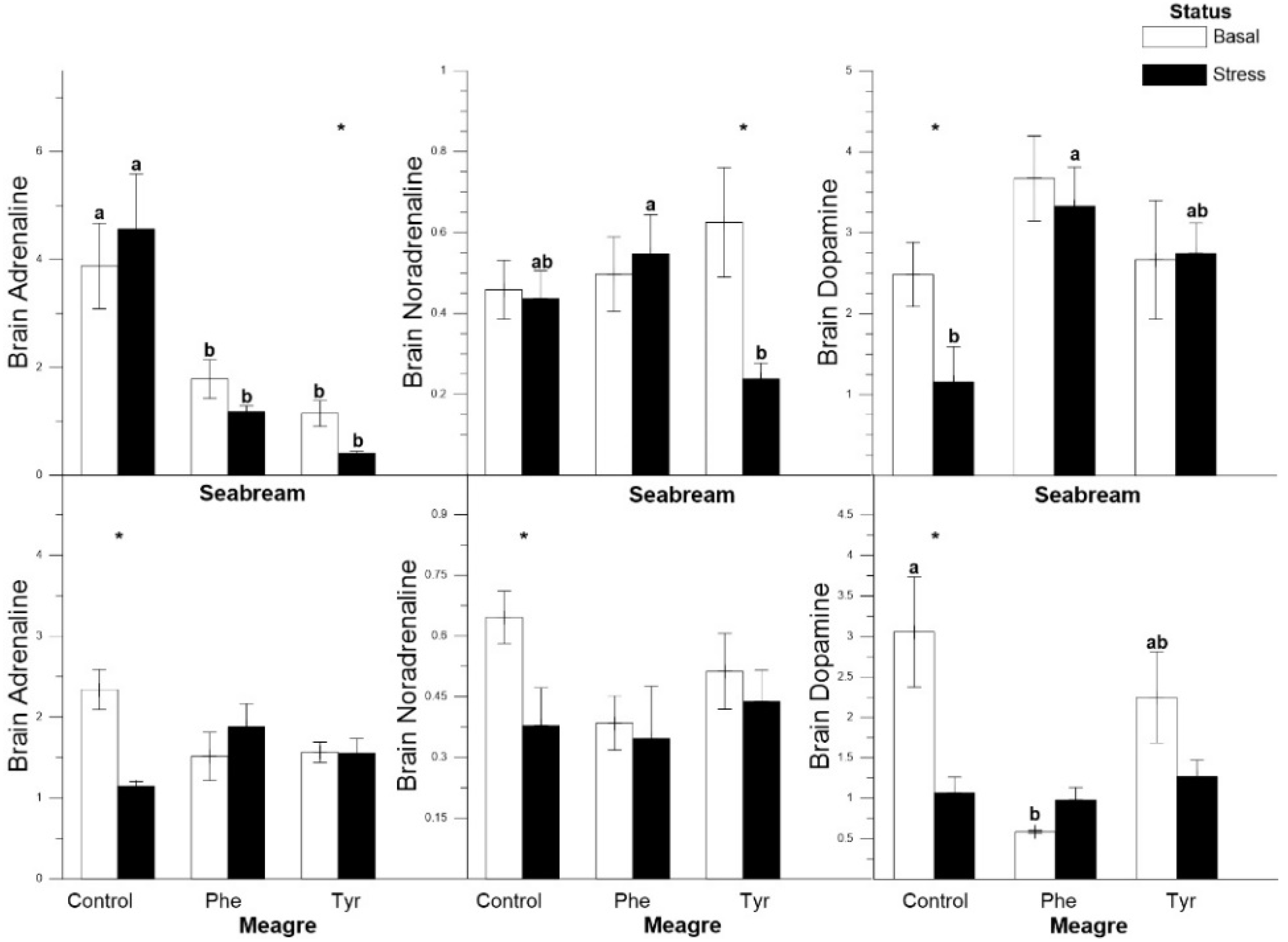
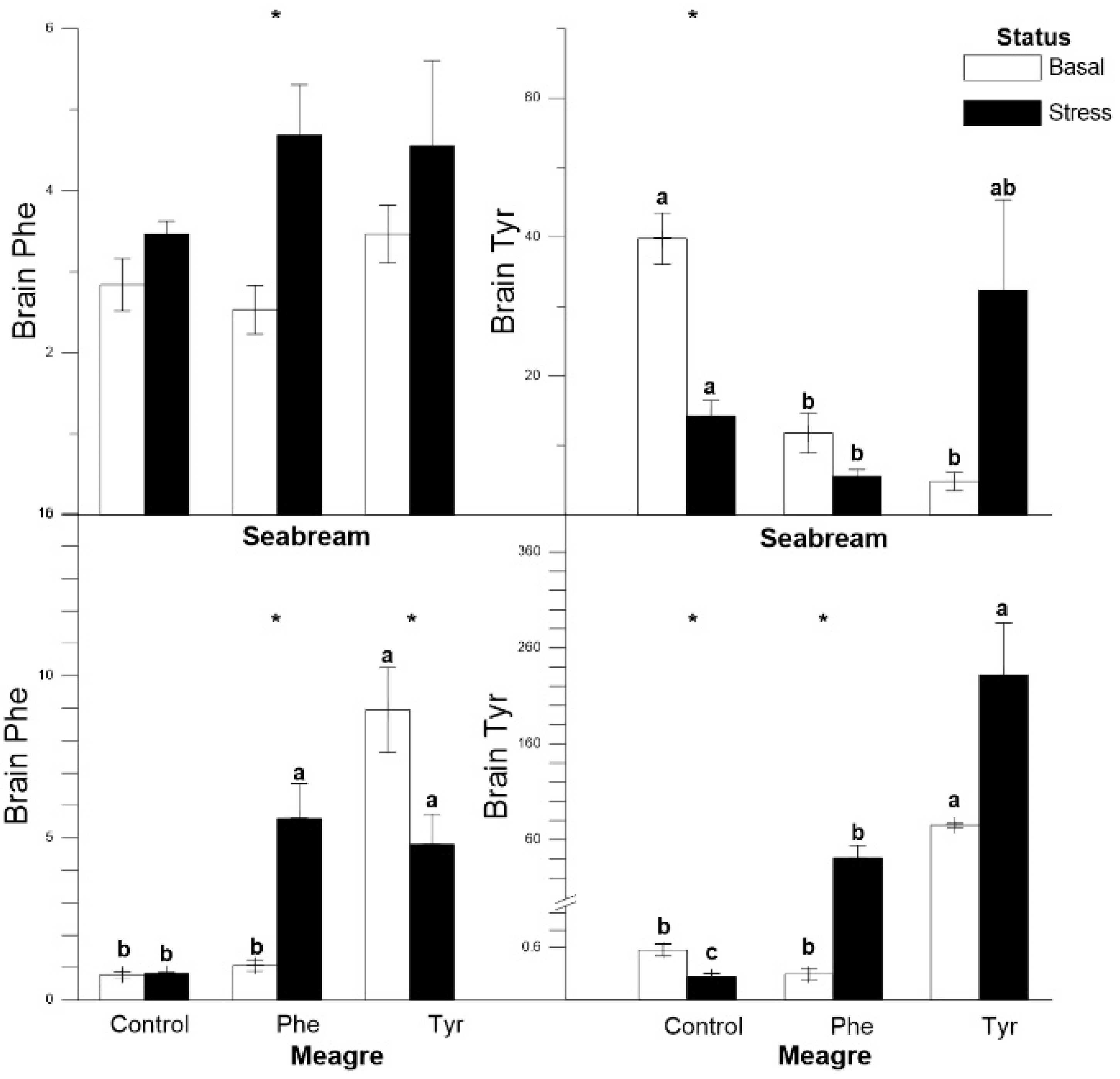
3.4. Tissue Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- AGAPA. La Acuicultura Marina en Andalucía 2018; Agencia de Gestión Agraria y Pesquera de Andalucía; Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible: Sevilla, España, 2019; p. 56.
- Arends, R.J.; Mancera, J.M.; Munoz, J.L.; Bonga, S.W.; Flik, G. The stress response of the gilthead seabream (Sparus aurata L.) to air exposure and confinement. J. Endocrinol. 1999, 163, 149. [Google Scholar] [CrossRef] [PubMed]
- Feidantsis, K.; Pörtner, H.O.; Lazou, A.; Kostoglou, B.; Michaelidis, B. Metabolic and molecular stress responses of the gilthead seabream Sparus aurata during long-term exposure to increasing temperatures. Mar. Biol. 2009, 156, 797–809. [Google Scholar] [CrossRef]
- Jerez-Cepa, I.; Fernández-Castro, M.; Del Santo, T.J.; Martos-Sitcha, J.A.; Martínez-Rodríguez, G.; Mancera, J.M.; Ruiz-Jarabo, I. Transport and recovery of gilthead seabream (Sparus aurata L.) sedated with clove oil and MS-222: Effects on stress axis regulation and intermediary metabolism. Front. Physiol. 2019, 10, 612. [Google Scholar] [CrossRef] [PubMed]
- Martos-Sitcha, J.A.; Wunderink, Y.S.; Straatjes, J.; Skrzynska, A.K.; Mancera, J.M.; Martínez-Rodríguez, G. Different stressors induce differential responses of the CRH-stress system in the gilthead seabream (Sparus aurata). Com. Biochem. Phys. A 2014, 177, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero, D.; Izquierdo, M.; Tort, L.; Robaina, L.; Vergara, J. High stocking density produces crowding stress altering some physiological and biochemical parameters in gilthead seabream, Sparus aurata, juveniles. Fish Physiol. Biochem. 1999, 20, 53–60. [Google Scholar] [CrossRef]
- Herrera, M.; Fernández-Alacid, L.; Sanahuja, I.; Ibarz, A.; Salamanca, N.; Morales, E.; Giráldez, I. Physiological and metabolic effects of a tryptophan-enriched diet to face up chronic stress in meagre (Argyrosomus regius). Aquaculture 2020, 522, 735102. [Google Scholar] [CrossRef]
- Hoseini, S.M.; Mirghaed, A.T.; Yousefi, M. Application of herbal anaesthetics in aquaculture. Rev. Aquac. 2019, 11, 550–564. [Google Scholar] [CrossRef]
- Ross, L.G.; Ross, B. Anaesthetic and Sedative Techniques for Aquatic Animals; Ross, L.G., Ross, B., Eds.; Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2008. [Google Scholar]
- Salamanca, N.; Morales, E.; Ruiz-Azcona, P.; Herrera, M. Endocrine and metabolic effects of Trp-enriched diets for attenuation of chronic stress in the Senegal soles (Solea senegalensis). Aquaculture 2020, 523, 735173. [Google Scholar] [CrossRef]
- Zahl, I.H.; Samuelsen, O.; Kiessling, A. Anaesthesia of farmed fish: Implications for welfare. Fish Physiol. Biochem. 2011, 38, 201–218. [Google Scholar] [CrossRef]
- Vanderzwalmen, M.; Eaton, L.; Mullen, C.; Henriquez, F.; Carey, P.; Snellgrove, D.; Sloman, K.A. The use of feed and water additives for live fish transport. Rev. Aquac. 2018, 11, 263–278. [Google Scholar] [CrossRef] [Green Version]
- Andersen, S.M.; Waagbø, R.; Espe, M. Functional amino acids in fish health and welfare. Front. Biol. 2006, 8, 143–169. [Google Scholar]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Herrera, M.; Ibarz, A. Skin mucus metabolites and cortisol in meagre fed acute stress-attenuating diets: Correlations between plasma and mucus. Aquaculture 2019, 499, 185–194. [Google Scholar] [CrossRef]
- Gonzalez-Silvera, D.; Herrera, M.; Giráldez, I.; Esteban, M. Ángeles effects of the dietary tryptophan and aspartate on the immune response of meagre (Argyrosomus regius) after stress. Fishes 2018, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Shafik, M.; Ibrahime, H.; Elyazeid, I.A.; Abass, O.; Saad, H.M. The stress of phenylalanine on rats to study the phenylketonuria at biochemical and molecular level. J. Appl. Pharm. Sci. 2014, 4, 24–29. [Google Scholar]
- Hoseini, S.M.; Hedayati, A.; Ghelichpour, M. Plasma metabolites, ions and thyroid hormones levels, and hepatic enzymes׳ activity in Caspian roach (Rutilus rutilus caspicus) exposed to waterborne manganese. Ecotoxicol. Environ. Saf. 2014, 107, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Banderet, L.E.; Lieberman, H.R. Treatment with tyrosine, a neurotransmitter precursor, reduces environmental stress in humans. Brain Res. Bull. 1989, 22, 759–762. [Google Scholar] [CrossRef]
- Brady, K.; Brown, J.W.; Thurmond, J.B. Behavioral and neurochemical effects of dietary tyrosine in young and aged mice following cold-swim stress. Pharmacol. Biochem. Behav. 1980, 12, 667–674. [Google Scholar] [CrossRef]
- Lehnert, H.; Reinstein, D.K.; Strowbridge, B.W.; Wurtman, R.J. Neurochemical and behavioral consequences of acute, uncontrollable stress: Effects of dietary tyrosine. Brain Res. 1984, 303, 215–223. [Google Scholar] [CrossRef]
- Cotoia, A.; Scrima, R.; Gefter, J.V.; Piccoli, C.; Cinnella, G.; Dambrosio, M.; Fink, M.P.; Capitanio, N. p-Hydroxyphenylpyruvate, an intermediate of the Phe/Tyr catabolism, improves mitochondrial oxidative metabolism under stressing conditions and prolongs survival in rats subjected to profound hemorrhagic shock. PLoS ONE 2014, 9, e90917. [Google Scholar] [CrossRef] [Green Version]
- Saavedra, M.; Conceição, L.E.; Barr, Y.; Helland, S.; Ferreira, P.P.; Yúfera, M.; Dinis, M.T. Tyrosine and phenylalanine supplementation onDiplodus sarguslarvae: Effect on growth and quality. Aquac. Res. 2010, 41, 1523–1532. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.; Herves, M.A.; Giráldez, I.; Skar, K.; Mogren, H.; Mortensen, A.; Puvanendran, V. Effects of amino acid supplementations on metabolic and physiological parameters in Atlantic cod (Gadus morhua) under stress. Fish Physiol. Biochem. 2016, 43, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.; Aragão, C.; Hachero, I.; Ruiz-Jarabo, I.; Vargas-Chacoff, L.; Mancera, J.M.; Conceição, L.E.C. Physiological short-term response to sudden salinity change in the Senegalese sole (Solea senegalensis). Fish Physiol. Biochem. 2012, 38, 1741–1751. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.; Rodiles, A.; Sánchez, B.; López, J.M.; De la Roca, E. Physiological stress responses to captivity in early developmental stages of the wedge sole Dicologoglossa cuneata (Moreau). Aquac. Res. 2014, 47, 732–740. [Google Scholar] [CrossRef]
- Qiu, Y.; Su, M.; Chen, M.; Liu, Y.; Gu, J.; Zhang, J.; Jia, W. Application of ethyl chloroformate derivatization for gaschromatography mass spectrometry based metabonomic profiling. Anal. Chim. Acta 2007, 583, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.-H.; Wang, J.; Gage, D.A.; Watson, J.T.; Sweeley, C.C.; Hušek, P. Characterization of N-ethoxycarbonyl ethyl esters of amino acids by mass spectrometry. J. Chromatogr. A 1993, 635, 271–281. [Google Scholar] [CrossRef]
- Ren, M.; Liu, B.; Habte-Tsion, H.-M.; Ge, X.; Xie, J.; Zhou, Q.; Liang, H.; Zhao, Z.; Pan, L. Dietary phenylalanine requirement and tyrosine replacement value for phenylalanine of juvenile blunt snout bream, Megalobrama amblycephala. Aquaculture 2015, 442, 51–57. [Google Scholar] [CrossRef]
- Zehra, S.; Khan, M.A. Dietary phenylalanine requirement and tyrosine replacement value for phenylalanine for fingerling Catla catla (Hamilton). Aquaculture 2014, 433, 256–265. [Google Scholar] [CrossRef]
- Fernández-Alacid, L.; Sanahuja, I.; Ordóñez-Grande, B.; Sánchez-Nuño, S.; Viscor, G.; Gisbert, E.; Herrera, M.; Ibarz, A. Skin mucus metabolites in response to physiological challenges: A valuable non-invasive method to study teleost marine species. Sci. Total Environ. 2018, 644, 1323–1335. [Google Scholar] [CrossRef]
- Li, P.; Mai, K.; Trushenski, J.; Wu, G. New developments in fish amino acid nutrition: Towards functional and environmentally oriented aquafeeds. Amino Acids 2008, 37, 43–53. [Google Scholar] [CrossRef]
- Höglund, E.; Balm, P.H.; Winberg, S. Skin darkening, a potential social signal in subordinate Arctic charr (Salvelinus alpinus): The regulatory role of brain monoamines and pro-opiomelanocortin-derived peptides. J. Exp. Biol. 2000, 203, 1711–1721. [Google Scholar]
- Øverli, Ø.; Harris, C.A.; Winberg, S. Short-term effects of fights for social dominance and the establishment of dominant-subordinate relationships on brain monoamines and cortisol in rainbow trout. Brain Behav. Evol. 1999, 54, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Höglund, E.; Kolm, N.; Winberg, S. Stress-induced effects on brain serotonergic activity, plasma cortisol and aggressive behaviour in Arctic charr (Salvelinus alpinus) is counteracted by L-dopa. Physiol. Behav. 2001, 74, 381–389. [Google Scholar] [CrossRef]
- Güttler, F.; Lou, H. Dietary problems of phenylketonuria: Effect on CNS transmitters and their possible role in behaviour and neuropsychological function. J. Inherit. Metab. Dis. 1986, 9, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Costa, C.A.E.; Alves, A.; Pinto, P.C.R.; Sousa, R.A.; Da Silva, E.B.; Reis, R.L.; Rodrigues, A.E. Characterization of ulvan extracts to assess the effect of different steps in the extraction procedure. Carbohydr. Polym. 2012, 88, 537–546. [Google Scholar] [CrossRef]
- Peter, M.S. The role of thyroid hormones in stress response of fish. Gen. Comp. Endocrinol. 2011, 172, 198–210. [Google Scholar] [CrossRef] [PubMed]
- Winberg, S.; Nilsson, G.E. Induction of social dominance by L-dopa treatment in Arctic charr. NeuroReport 1992, 3, 243–246. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Gilthead Seabream | Meagre | ||||
|---|---|---|---|---|---|---|
| Ctrl | Phe | Tyr | Ctrl | Phe | Tyr | |
| Plasma glucose | R↑ | R↑ | R↑ | R↑ | C | C |
| Plasma lactate | R↑ | R↑ | R↑ | C | C | C |
| Plasma proteins | C | C | C | C | C | C |
| Plasma cortisol | R↑ | R↑ | R↑ | C | C | R↑ |
| Plasma adrenaline | C | C | C | C | C | C |
| Plasma noradrenaline | C | C | C | C | C | C |
| Plasma T3 | C | R↓ | C | C | R↑ | C |
| Plasma T4 | C | C | C | R↓ | C | C |
| Brain adrenaline | C | C | R↓ | R↓ | C | C |
| Brain noradrenaline | C | C | R↓ | R↓ | C | C |
| Brain dopamine | R↓ | C | C | R↓ | C | C |
| Brain Phe | C | C | C | C | C | C |
| Brain Tyr | C | C | C | C | C | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salamanca, N.; Giráldez, I.; Morales, E.; de La Rosa, I.; Herrera, M. Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre. Animals 2021, 11, 45. https://doi.org/10.3390/ani11010045
Salamanca N, Giráldez I, Morales E, de La Rosa I, Herrera M. Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre. Animals. 2021; 11(1):45. https://doi.org/10.3390/ani11010045
Chicago/Turabian StyleSalamanca, Natalia, Inmaculada Giráldez, Emilio Morales, Ignacio de La Rosa, and Marcelino Herrera. 2021. "Phenylalanine and Tyrosine as Feed Additives for Reducing Stress and Enhancing Welfare in Gilthead Seabream and Meagre" Animals 11, no. 1: 45. https://doi.org/10.3390/ani11010045