Study on the Function and Mechanism of Lin28B in the Formation of Chicken Primordial Germ Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Reagents
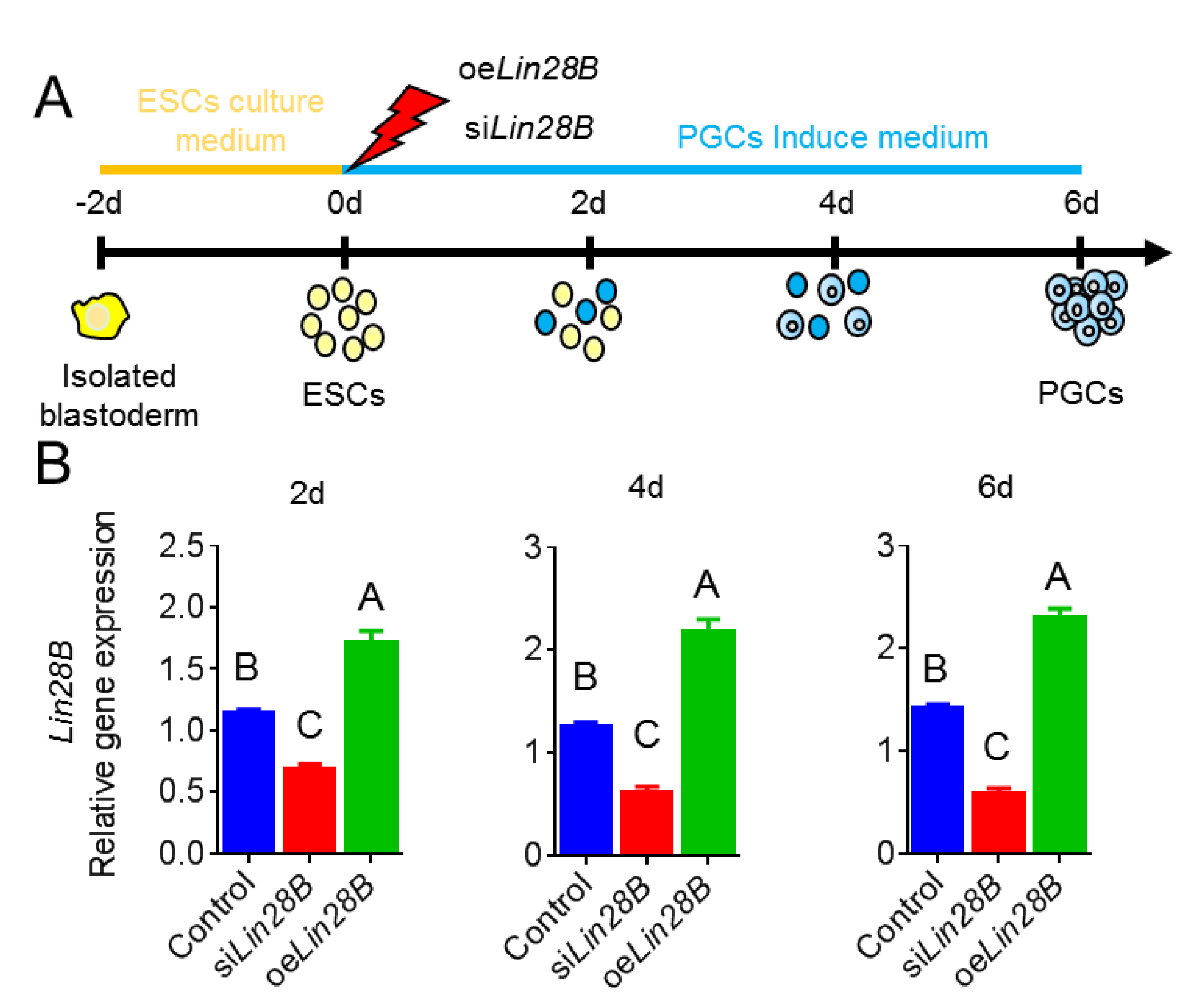
2.3. Cell Treatments and Grouping
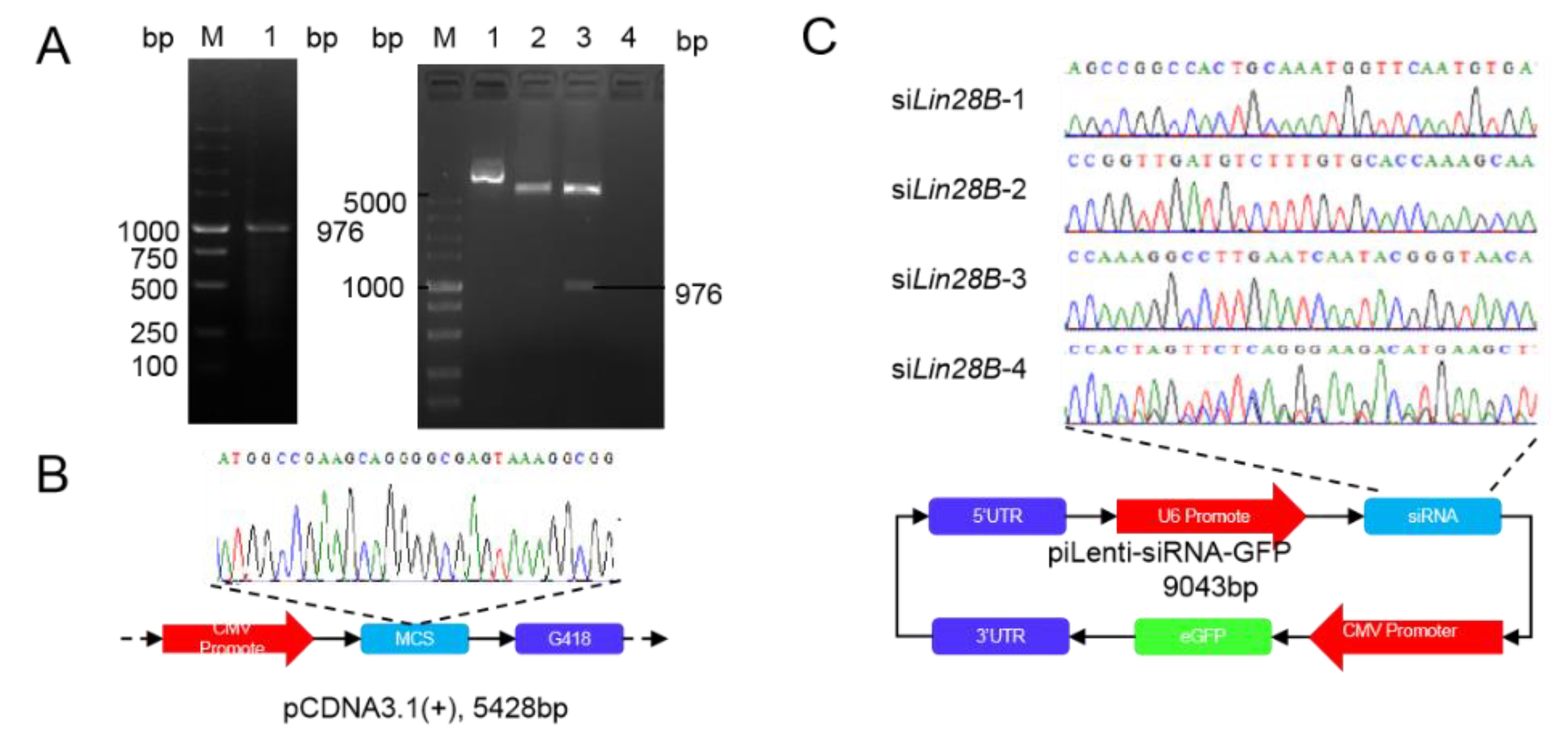
2.4. Construction of Lin28B Overexpression Vector
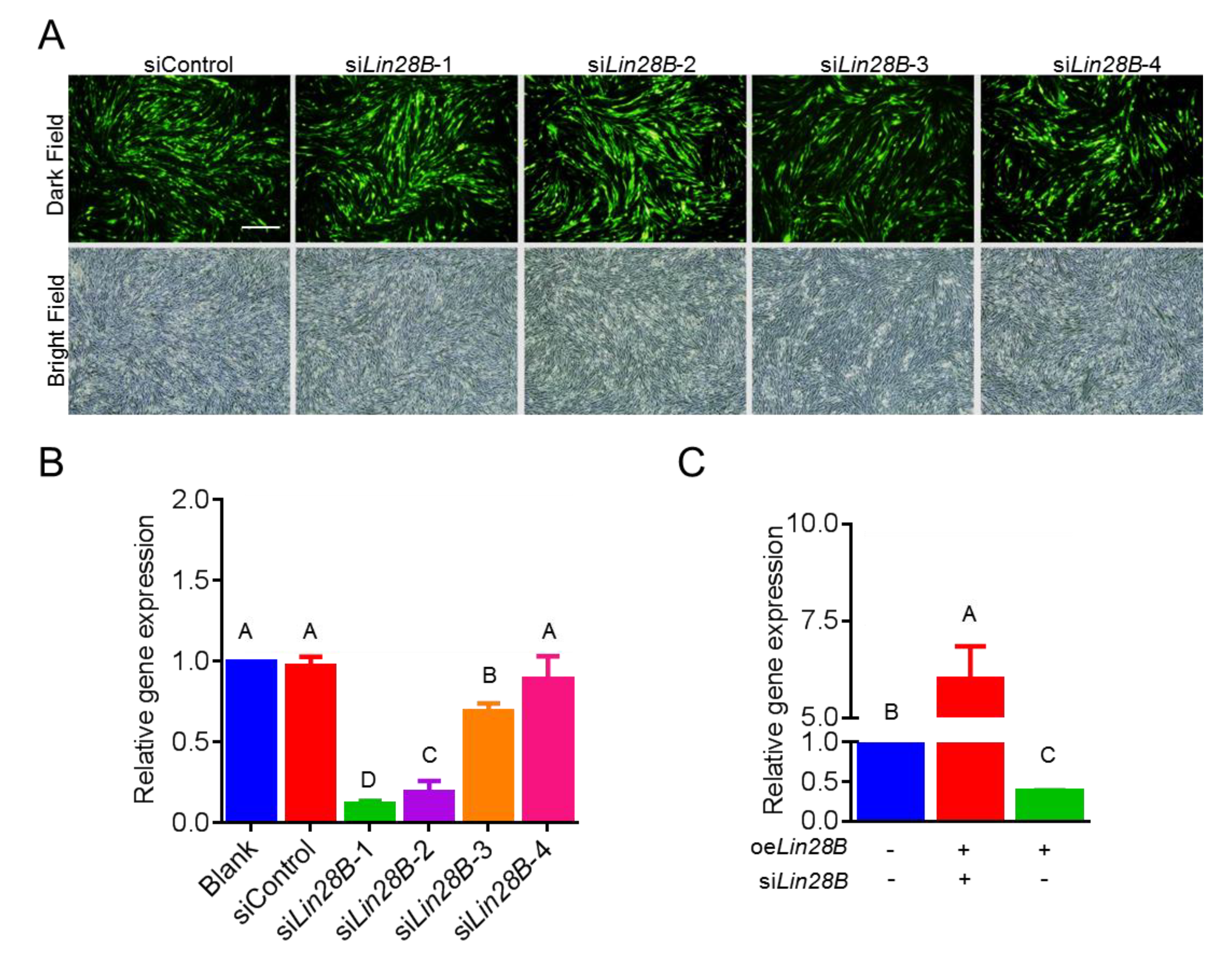
2.5. Construction of the Lin28B Interference Vector
2.6. qRT-PCR
2.7. PAS Staining
2.8. Screening and Testing of Lin28B-Binding micRNA
2.9. Immunocytochemical Detection of Reproductive Marker Protein
2.10. Data Analysis
3. The Results
3.1. Subsection Construction of Overexpression and Interference Vector of Lin28B
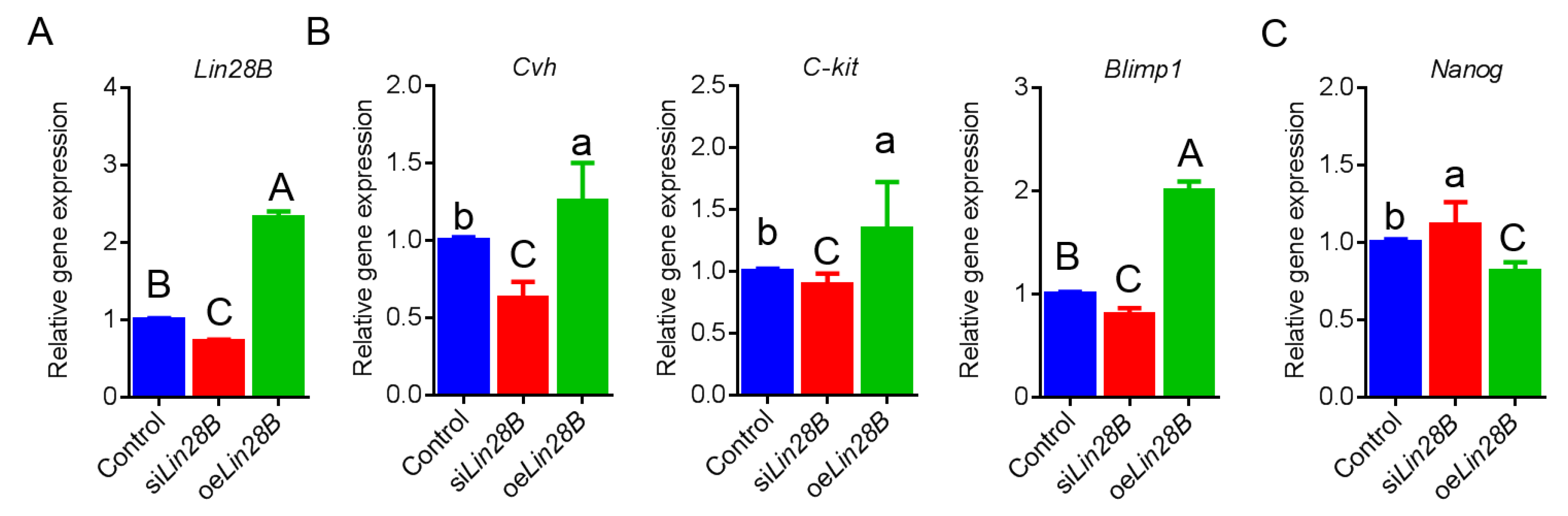
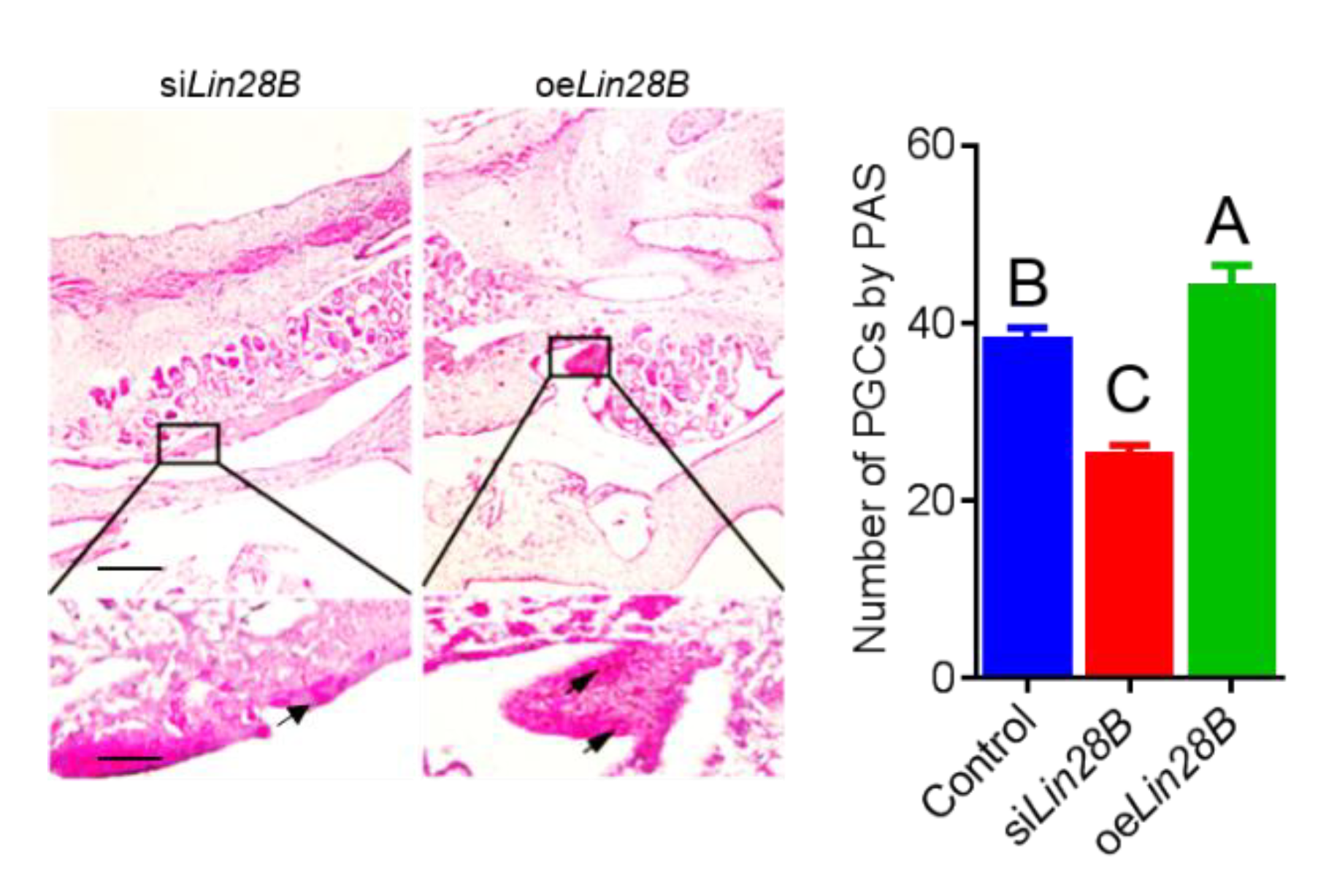
3.2. Lin28B Overexpression/Interference Could Promote/Inhibit PGCs Formation In Vivo
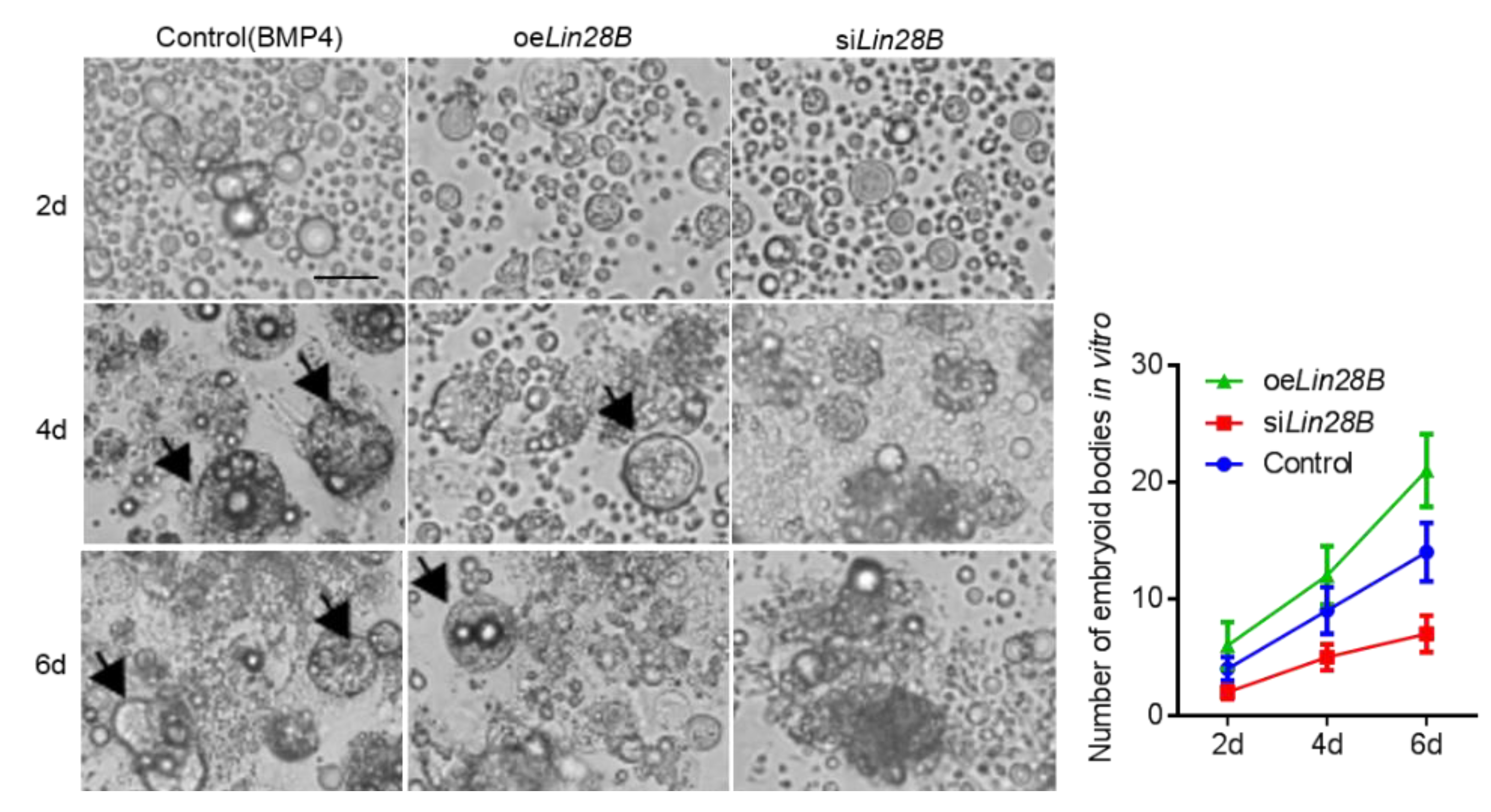
3.3. Lin28B Could Promote Formation of EB in Bmp4-Induced Model In Vitro
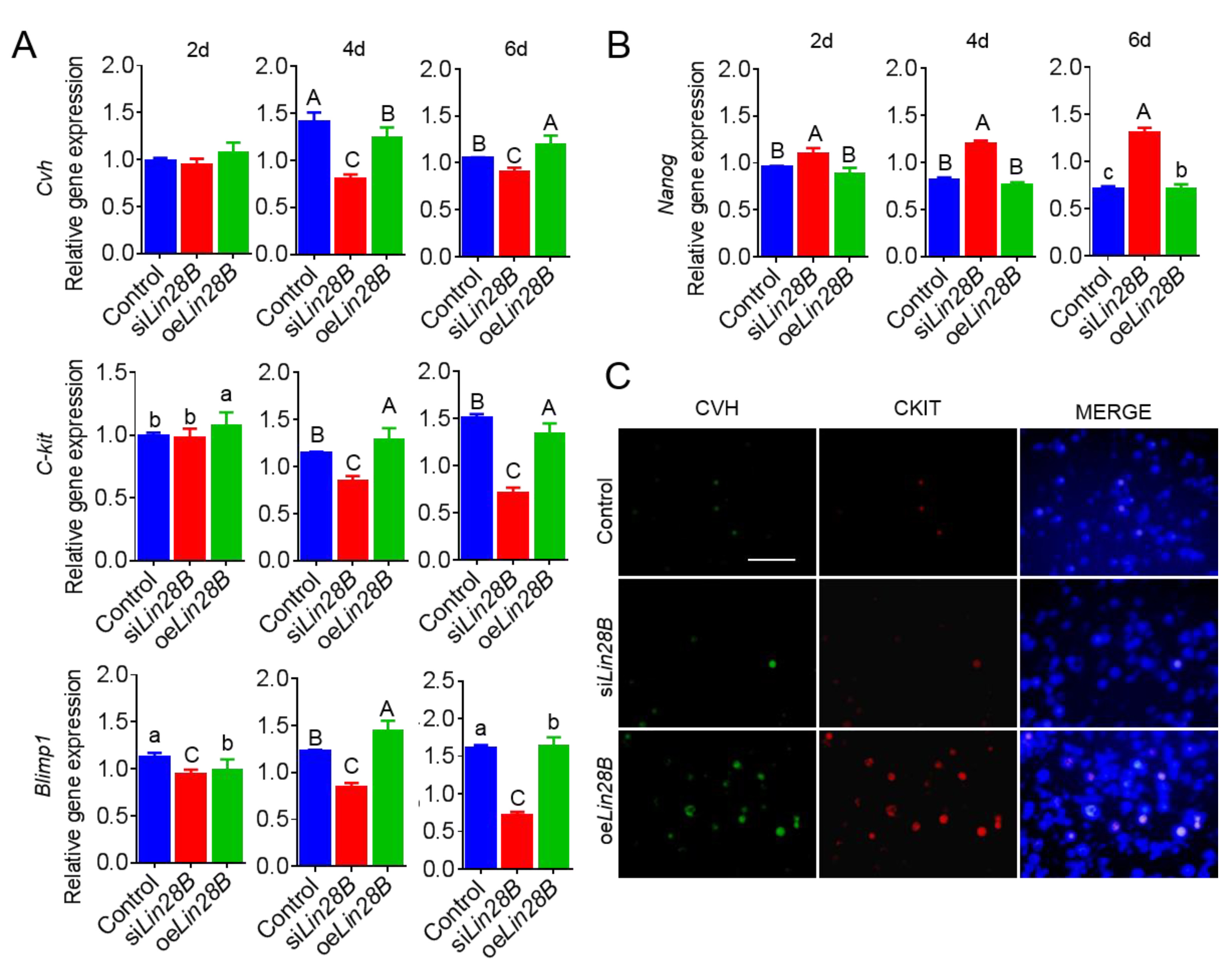
3.4. Lin28B Could Positively Modulate Formation of PGCs in Bmp4-Induced Model In Vitro
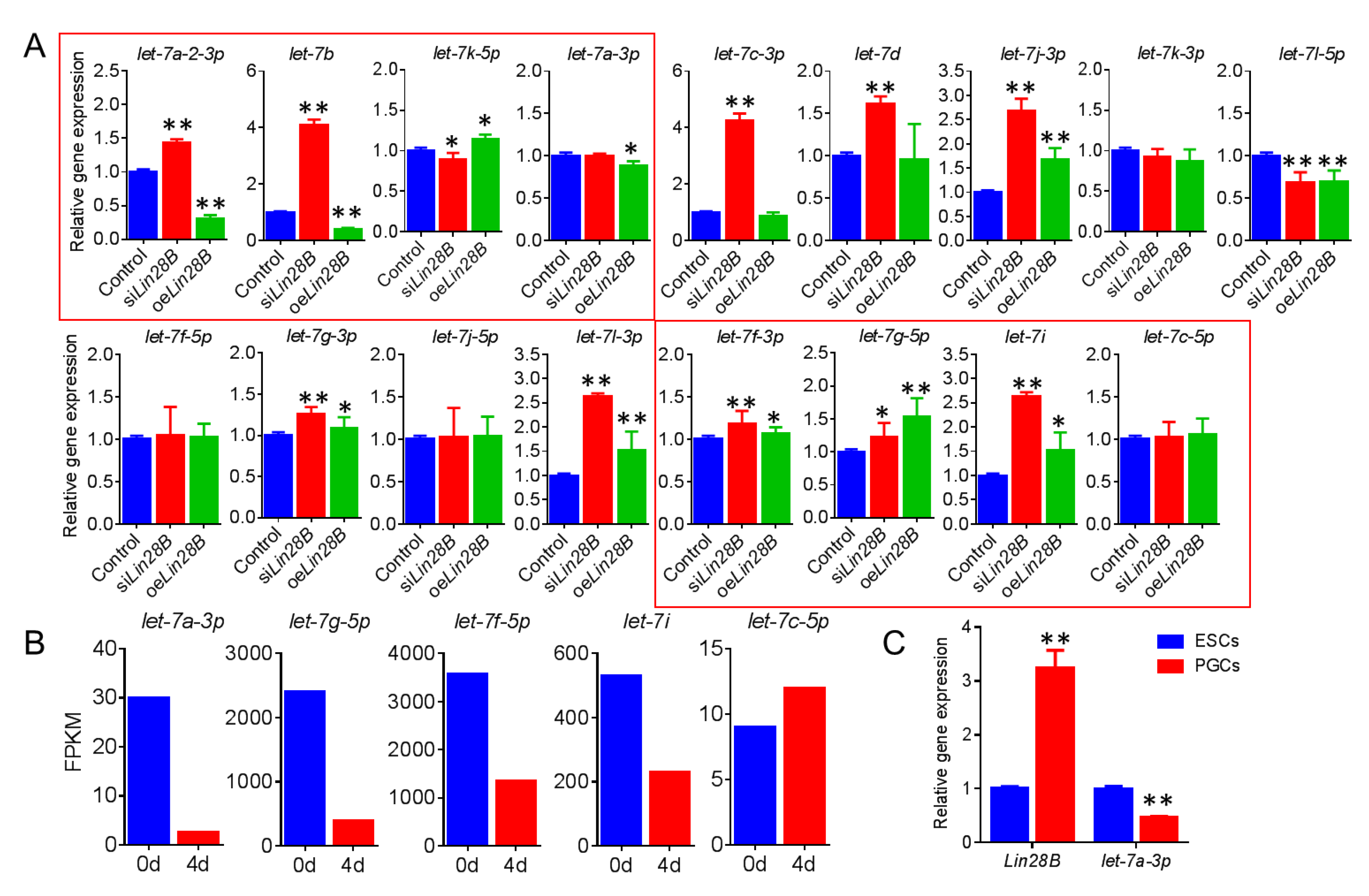
3.5. Lin28B Promotes Formation of PGCs Through Inhibition of gga-Let-7a-3p
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faas, L.; Warrander, F.C.; Maguire, R.; Ramsbottom, S.A.; Quinn, D.; Genever, P.; Isaacs, H.V. Lin28 proteins are required for germ layer specification in Xenopus. Development 2013, 140, 976–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, Y.; Yamamoto, J.; Iwamoto, T. The Heterochronic Genes lin-28a and lin-28b Play an Essential and Evolutionarily Conserved Role in Early Zebrafish Development. PLoS ONE 2014, 9, e88086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, S.; Bohndorf, M.; Schröter, F.; Assar, F.; Wruck, W.; Sleegers, K.; Van Broeckhoven, C.; Adjaye, J. Lymphoblast-derived integration-free ISRM-CON9 iPS cell line from a 75 year old female. Stem Cell Res. 2017, 26, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, M.; Attari, F.; Khoshnam, S.E. Concise Review: LIN28/let-7 Signaling, a Critical Double-Negative Feedback Loop During Pluripotency, Reprogramming, and Tumorigenicity. Cell. Reprogramming 2017, 19, 289–293. [Google Scholar] [CrossRef]
- Moss, E.G.; Lee, R.C.; Ambros, V. The Cold Shock Domain Protein LIN-28 Controls Developmental Timing in C. elegans and Is Regulated by the lin-4 RNA. Cell 1997, 88, 637–646. [Google Scholar] [CrossRef] [Green Version]
- West, J.A.; Viswanathan, S.R.; Yabuuchi, A.; Cunniff, K.; Takeuchi, A.; Park, I.H.; Sero, J.E.; Zhu, H.; Perez-Atayde, A.; Frazier, A.L.; et al. A role for Lin28 in primordial germ-cell development and germ-cell malignancy. Nature 2009, 460, 909–913. [Google Scholar] [CrossRef] [Green Version]
- Tran, N.D.; Laird, D.; Kissner, M.; Conti, M.; Blelloch, R. Lin28-let7-Blimp1 Circuitry regulates human primordial germ cells (hPGC) development from human embryonic stem cells (hESC). Fertil. Steril. 2011, 96, 104–115. [Google Scholar] [CrossRef]
- Matzuk, M.M. Lin28 lets BLIMP1 take the right course. Dev. Cell 2009, 17, 160–161. [Google Scholar] [CrossRef] [Green Version]
- Perry, J.R.B.; Stolk, L.; Franceschini, N.; Lunetta, K.L.; Zhai, G.; McArdle, P.F.; Smith, A.V.; Aspelund, T.; Bandinelli, S.; Boerwinkle, E.; et al. Meta-analysis of genome-wide association data identifies two loci influencing age at menarche. PLoS ONE 2014, 41, 648–650. [Google Scholar] [CrossRef] [Green Version]
- Ong, K.K.; Elks, C.E.; Li, S.; Zhao, J.H.; Andersen, L.B.; Bingham, S.A.; Brage, S.; Smith, G.D.; Ekelund, U.; Gillson, C.J.; et al. Genetic variation in LIN28B is associated with the timing of puberty. Nat. Genet. 2009, 41, 729–733. [Google Scholar] [CrossRef] [Green Version]
- Sulem, P.; Gudbjartsson, D.F.; Rafnar, T.; Holm, H.; Olafsdottir, E.J.; Olafsdottir, G.H.; Jonsson, T.; Alexandersen, P.; Feenstra, B.; Boyd, H.A.; et al. Genome-wide association study identifies sequence variants on 6q21 associated with age at menarche. Nat. Genet. 2009, 41, 734–738. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, G.; Shyh-Chang, N.; Soysa, T.Y.D.; Zhu, H.; Seligson, M.T.; Shah, S.P.; Abo-Sido, N.; Yabuuchi, A.; Hagan, J.P.; Gregory, R.I.; et al. Fetal Deficiency of Lin28 Programs Life-Long Aberrations in Growth and Glucose Metabolism. Stem Cells 2013, 31, 1563–1573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Hu, Z.; Pan, X.; Li, M.; Togun, T.A.; Tuck, D.; Pelizzola, M.; Huang, J.; Ye, X.; Yin, Y.; et al. Germline competency of parthenogenetic embryonic stem cells from immature oocytes of adult mouse ovary. Hum. Mol. Genet. 2011, 20, 1339–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, A.J.; Kinnell, H.L.; He, J.; Anderson, R.A. LIN28 Is Selectively Expressed by Primordial and Pre-Meiotic Germ Cells in the Human Fetal Ovary. Stem Cells Dev. 2012, 21, 2343–2349. [Google Scholar] [CrossRef] [Green Version]
- El-Khairi, R.; Parnaik, R.; Duncan, A.J.; Lin, L.; Gerrelli, D.; Dattani, M.T.; Conway, G.S.; Achermann, J.C. Analysis of LIN28A in early human ovary development and as a candidate gene for primary ovarian insufficiency. Mol. Cell. Endocrinol. 2012, 351, 264–268. [Google Scholar] [CrossRef]
- Shyh-Chang, N.; Daley, G.Q. Lin28: Primal Regulator of Growth and Metabolism in Stem Cells. Cell Stem Cell 2013, 12, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Mayr, F.; Heinemann, U. Mechanisms of Lin28-Mediated miRNA and mRNA Regulation—A Structural and Functional Perspective. Int. J. Mol. Sci. 2013, 14, 16532–16553. [Google Scholar] [CrossRef]
- Nam, Y.; Chen, C.; Gregory, R.I.; Chou, J.J.; Sliz, P. Molecular Basis for Interaction of let-7 MicroRNAs with Lin28. Cell 2011, 147, 1080–1091. [Google Scholar] [CrossRef] [Green Version]
- Thornton, J.E.; Chang, H.M.; Piskounova, E.; Gregory, R.I. Lin28-mediated control of let-7 microRNA expression by alternative TUTases Zcchc11 (TUT4) and Zcchc6 (TUT7). RNA 2012, 18, 1875–1885. [Google Scholar] [CrossRef] [Green Version]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; LaPierre, R.J.; Pothoulakis, C.; Hagan, J.P.; Iliopoulos, D.; Gregory, R.I. Lin28A and Lin28B Inhibit let-7 MicroRNA Biogenesis by Distinct Mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Y.; Zuo, Q.; Li, D.; Zhang, W.; Lian, C.; Tang, B.; Xiao, T.; Wang, M.; Wang, K.; et al. Effects of the Transforming Growth Factor Beta Signaling Pathway on the Differentiation of Chicken Embryonic Stem Cells into Male Germ Cells. Cell Reprogramming 2016, 18, 401–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Q.Q.; Sun, M.; Zhang, Z.T.; Zhang, Y.N.; Elsayed, A.K.; Zhang, L.; Huang, X.M.; Li, B.C. A screen of suitable inducers for germline differentiation of chicken embryonic stem cells. Anim. Reprod. Sci. 2014, 147, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Elks, C.E.; Perry, J.R.; Sulem, P.; Chasman, D.I.; Franceschini, N.; He, C.; Lunetta, K.L.; Visser, J.A.; Byrne, E.M.; Cousminer, D.L.; et al. Thirty new loci for age at menarche identified by a meta-analysis of genome-wide association studies. Nat. Genet. 2010, 42, 1077–1085. [Google Scholar] [CrossRef] [Green Version]
- Chatterji, P.; Rustgi, A.K. RNA Binding Proteins in Intestinal Epithelial Biology and Colorectal Cancer. Trends Mol. Med. 2018, 24, 490–506. [Google Scholar] [CrossRef] [PubMed]
- Thomson, J.M.; Newman, M.; Parker, J.S.; Morin-Kensicki, E.M.; Wright, T.; Hammond, S.M. Extensive post-transcriptional regulation of microRNAs and its implications for cancer. Genes Dev. 2006, 20, 2202–2207. [Google Scholar] [CrossRef] [Green Version]
- Büssing, I.; Slack, F.J.; Großhans, H. let-7 microRNAs in development, stem cells and cancer. Trends Mol. Med. 2008, 14, 400–409. [Google Scholar] [CrossRef]
- Nie, K.; Gomez, M.; Landgraf, P.; Garcia, J.F.; Liu, Y.; Tan, L.H.; Chadburn, A.; Tuschl, T.; Knowles, D.M.; Tam, W. MicroRNA-Mediated Down-Regulation of PRDM1/Blimp-1 in Hodgkin/Reed-Sternberg Cells: A Potential Pathogenetic Lesion in Hodgkin Lymphomas. Am. J. Pathol. 2008, 173, 242–252. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, Q.; Zhou, J.; Wang, M.; Zhang, Y.; Chen, G.; Li, B. Study on the Function and Mechanism of Lin28B in the Formation of Chicken Primordial Germ Cells. Animals 2021, 11, 43. https://doi.org/10.3390/ani11010043
Zuo Q, Zhou J, Wang M, Zhang Y, Chen G, Li B. Study on the Function and Mechanism of Lin28B in the Formation of Chicken Primordial Germ Cells. Animals. 2021; 11(1):43. https://doi.org/10.3390/ani11010043
Chicago/Turabian StyleZuo, Qisheng, Jing Zhou, Man Wang, Yani Zhang, Guohong Chen, and Bichun Li. 2021. "Study on the Function and Mechanism of Lin28B in the Formation of Chicken Primordial Germ Cells" Animals 11, no. 1: 43. https://doi.org/10.3390/ani11010043