Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Sampling, DNA Preparation and Sequence Analysis
2.3. PCR Amplification and Sequencing
2.4. Statistical Analysis
3. Results
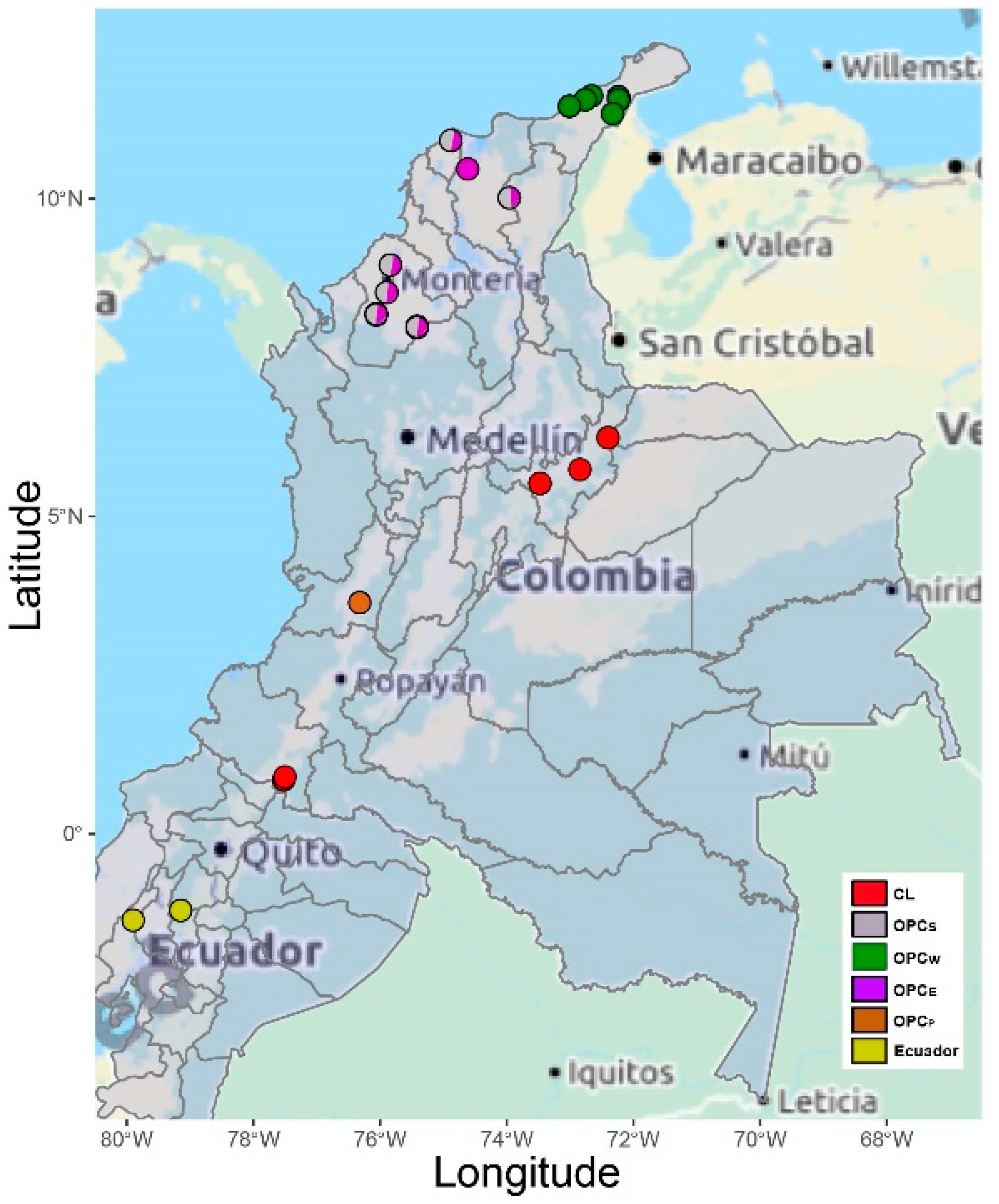
3.1. Genetic Diversity within Colombian Creole Sheep
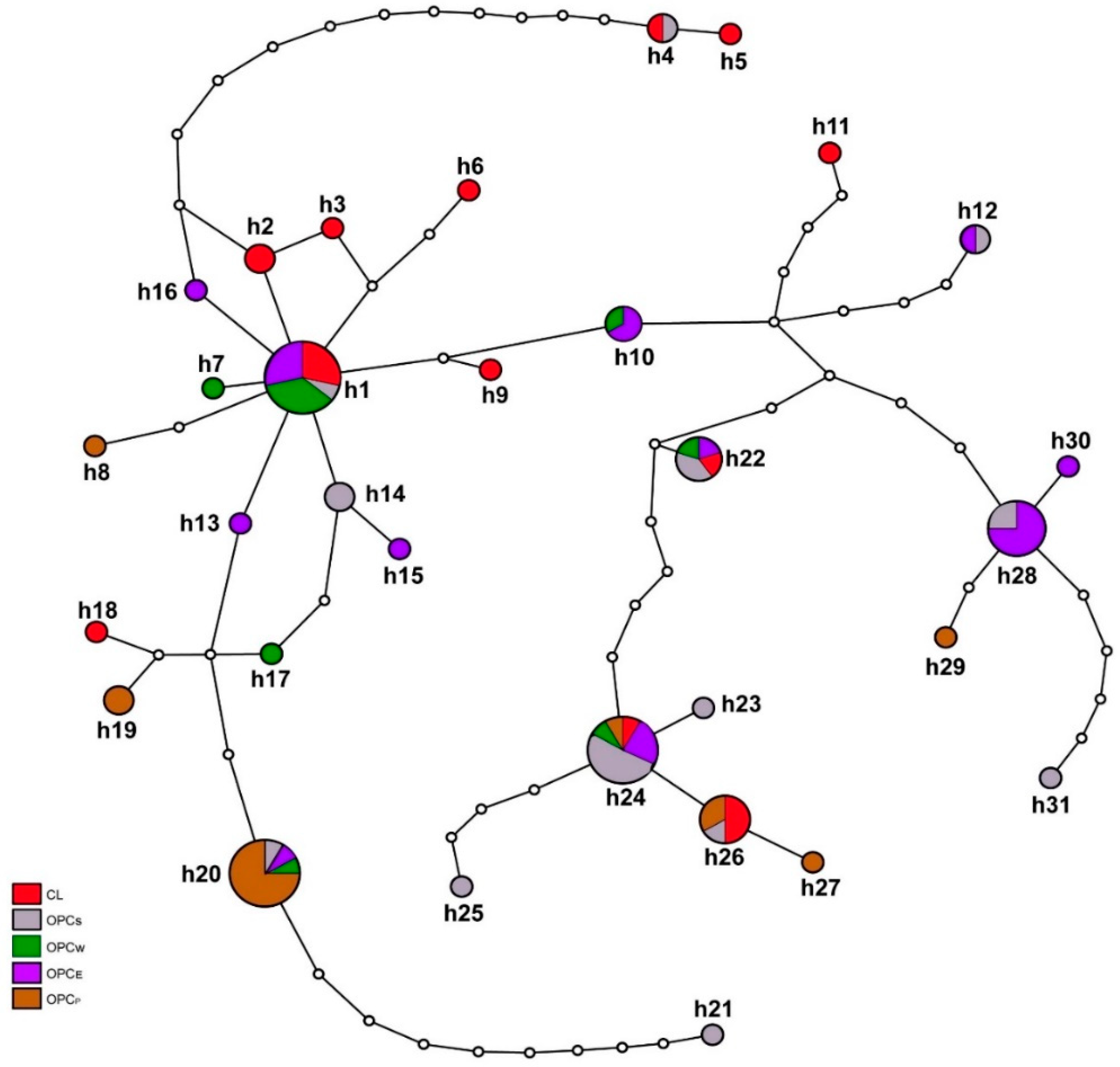
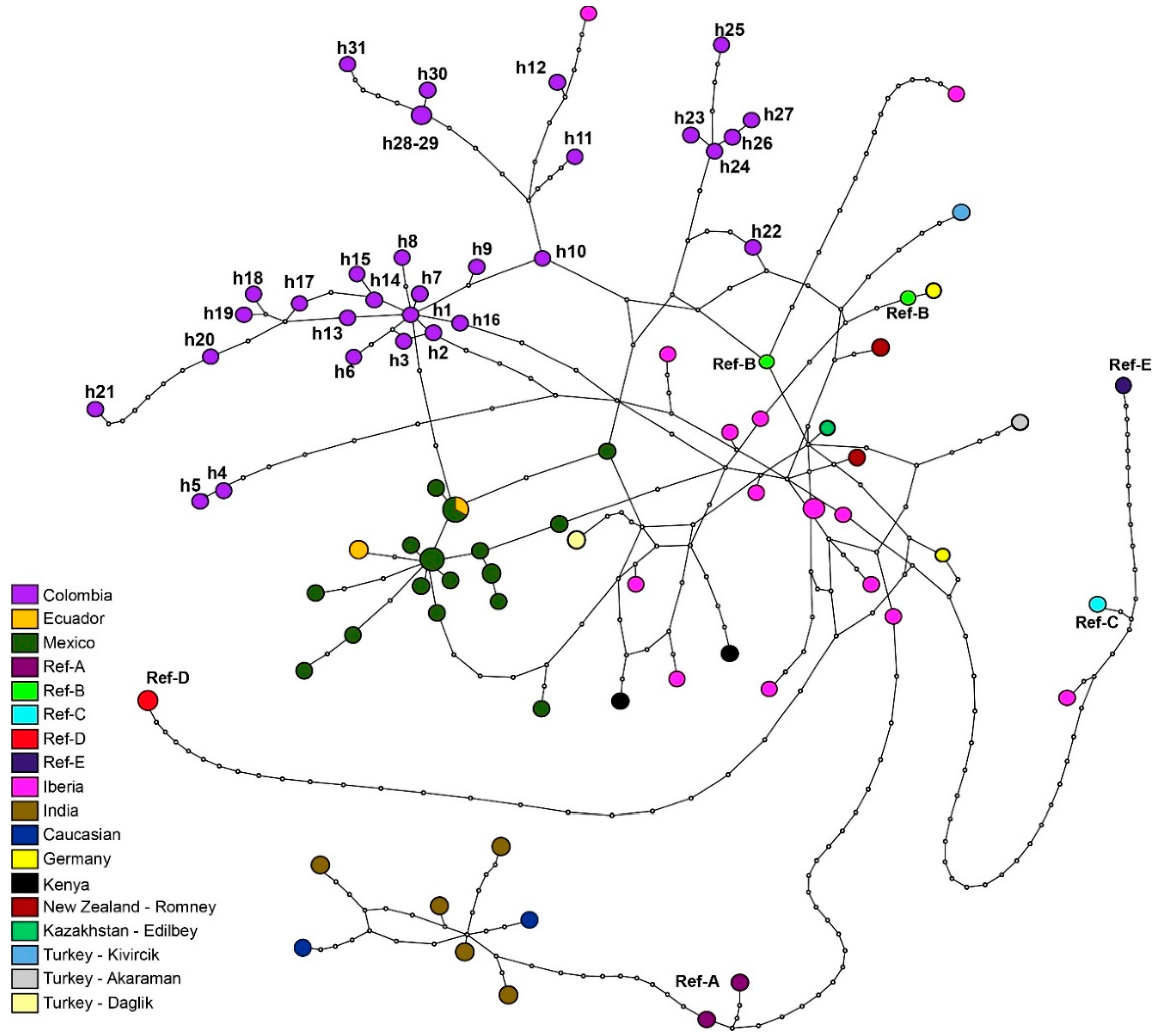
3.2. Genetic Structure of Colombian Creole Sheep
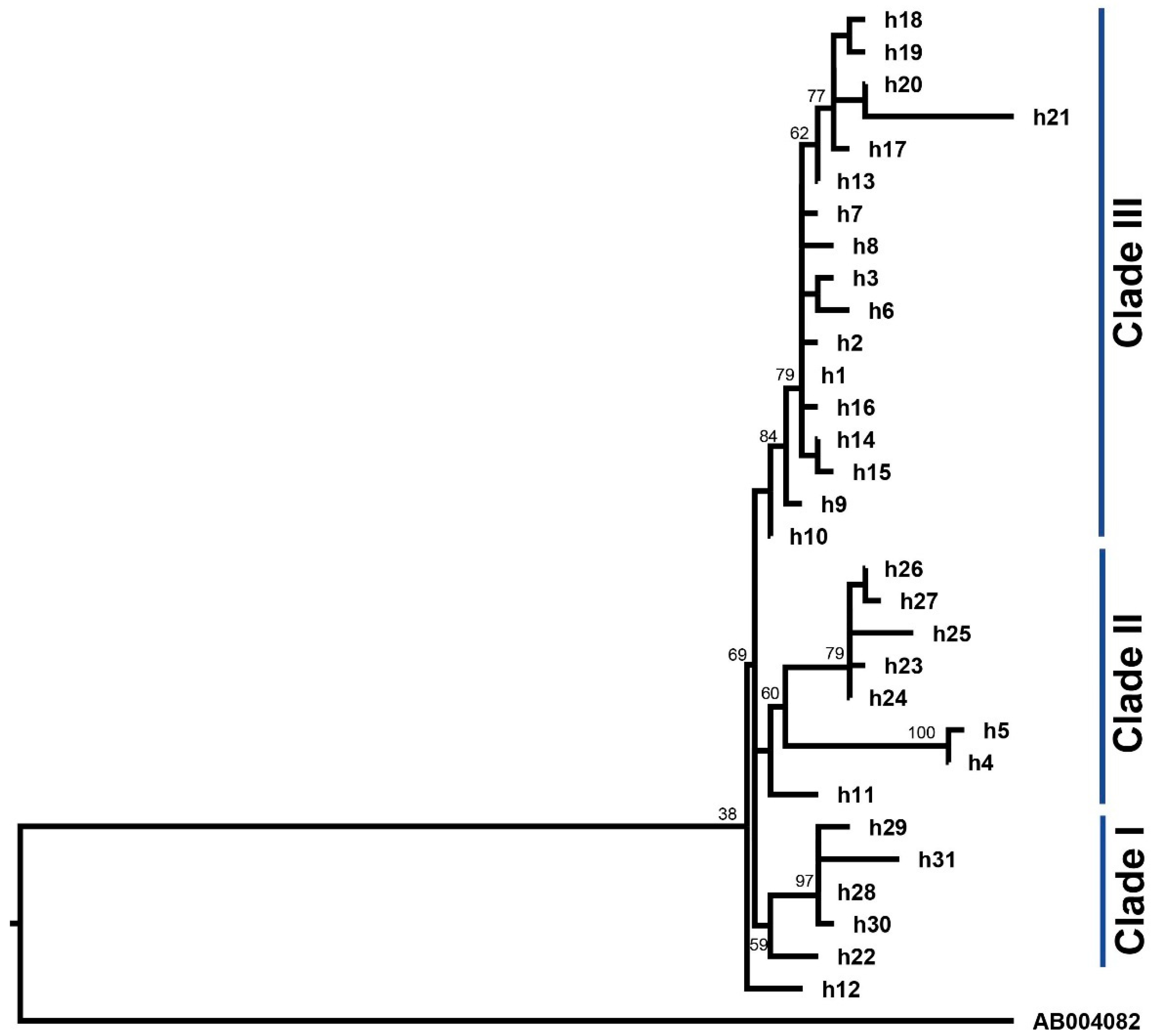
3.3. Phylogenetic Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Delgado, J.V.; León, J.M.; Gomez, M.; Nogales, S.; Camacho, M. Las razas ovinas ibéricas y su participación en la colonización de Iberoamérica. In Biodiversidad Ovina Iberoamericana—Caracterización y Uso Sustentable; Delgado, J.V., Nogales, S., Eds.; Universidad de Córdoba: Córdoba, Andalucía, Spain, 2010; pp. 17–30. [Google Scholar]
- Peña, S.; Martínez, A.; Villegas Castagnasso, E.; Aulicino, M.; Género, E.; Giovambattista, G.; Martínez, R.D. Caracterización genética de cuatro poblaciones de ovinos criollos de Argentina. J. Basic Appl. Genet. 2017, 28, 43–55. [Google Scholar]
- Alvarez, I.; Capote, J.; Traore, A.; Fonseca, N.; Perez, K.; Cuervo, M.; Fernandez, I.; Goyache, F. Mitochondrial analysis sheds light on the origin of hair sheep. Anim. Genet. 2013, 44, 344–347. [Google Scholar] [CrossRef] [PubMed]
- Alonso, R.A.; Ulloa-Arvizu, R.; Gayosso-Vazquez, A. Mitochondrial DNA sequence analysis of the Mexican Creole sheep (Ovis aries) reveals a narrow Iberian maternal origin. Mitochondrial DNA Part A DNA 2017, 28, 793–800. [Google Scholar] [CrossRef]
- Perezgrovas, R.; Parés, P.-M.; Hummel, J.; Zaragoza, L.; Delgado, J.V. Características de la lana en 4 razas de ovejas autóctonas de España y México. Actas Iberoam. Conserv. Anim. 2011, 1, 380–383. [Google Scholar]
- Perezgrovas Garza, R. Los Carneros de San Juan: Ovinocultura Indigena en Los Altos de Chiapas; Instituto de Estudios Indígenas, Universidad Autónoma de Chiapas: San Cristóbal de las Casas, Mexico, 2004. [Google Scholar]
- Pons, A.; Landi, V.; Martinez, A.; Delgado, J. The biodiversity and genetic structure of Balearic sheep breeds. J. Anim. Breed. Genet. 2015, 132, 268–276. [Google Scholar] [CrossRef]
- Spangler, G.L.; Rosen, B.D.; Ilori, M.B.; Hanotte, O.; Kim, E.-S.; Sonstegard, T.S.; Burke, J.M.; Morgan, J.L.M.; Notter, D.R.; Van Tassell, C.P. Whole genome structural analysis of Caribbean hair sheep reveals quantitative link to West African ancestry. PLoS ONE 2017, 12, e0179021. [Google Scholar] [CrossRef] [Green Version]
- Lancioni, H.; Di Lorenzo, P.; Ceccobelli, S.; Perego, U.A.; Miglio, A.; Landi, V.; Antognoni, M.T.; Sarti, F.M.; Lasagna, E.; Achilli, A. Phylogenetic Relationships of Three Italian Merino-Derived Sheep Breeds Evaluated through a Complete Mitogenome Analysis. PLoS ONE 2013, 8, e73712. [Google Scholar] [CrossRef] [Green Version]
- Miretti, M.; Dunner, S.; Naves, M.; Contel, E.P.; Ferro, J.A. Predominant African-Derived mtDNA in Caribbean and Brazilian Creole Cattle is also Found in Spanish Cattle (Bos taurus). J. Hered. 2004, 95, 450–453. [Google Scholar] [CrossRef] [Green Version]
- Ginja, C.; Penedo, M.C.T.; Melucci, L.; Quiroz, J.; López, O.R.M.; Revidatti, M.A.; Delgado, J.V.; Da Gama, L.T.; Martínez-Martínez, A. Origins and genetic diversity of New World Creole cattle: Inferences from mitochondrial and Y chromosome polymorphisms. Anim. Genet. 2010, 41, 128–141. [Google Scholar] [CrossRef]
- Pedrosa, S.; Arranz, J.J.; Brito, N.V.; Molina, A.; Primitivo, F.S.; Bayón, Y. Mitochondrial diversity and the origin of Iberian sheep. Genet. Sel. Evol. 2007, 39, 91–103. [Google Scholar] [CrossRef]
- Pereira, F.; Davis, S.J.M.; Pereira, L.; McEvoy, B.; Bradley, D.G.; Amorim, A. Genetic Signatures of a Mediterranean Influence in Iberian Peninsula Sheep Husbandry. Mol. Boil. Evol. 2006, 23, 1420–1426. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, R.; Cardona, H.; Martínez, R. Genetic diversity of Colombian sheep by microsatellite markers. Chil. J. Agric. Res. 2016, 76, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Del Arco Aguilar, M.M.; Hernández, C.G.; Adrián, M.C.R.; Armas, E.A.; Aguilar, M.J.; Aguilar, M.d.C. El menceyato de Icod en el poblamiento de Tenerife: D. Gaspar, Las Palomas y Los Guanches. Sobre el poblamiento y las estrategias de alimentación vegetal entre los guanches. Eres. Arqueol./Bioantropol. 2000, 9, 67–129. [Google Scholar]
- Arcos Dorado, J.C.; Romero Huertas, H.; Vanegas Rivera, M.A.; Riveros Escobar, E. Ovinos Colombianos de Pelo: Alternativa Productiva para el Sur del Departamento del Tolima.; Arcos Dorado, J.C., Romero Huertas, H., Vanegas Rivera, M.A., Riveros Escobar, E., Eds.; Corporación Colombiana de Investigación Agropecuaria: Tolima, Colombia, 2002; Volume 1. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Stecher, G.; Nei, M.; Kumar, S.; Peterson, N. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Boil. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Hiendleder, S.; Kaupe, B.; Wassmuth, R.; Janke, A. Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies. Proc. R Soc. B Boil. Sci. 2002, 269, 893–904. [Google Scholar] [CrossRef] [Green Version]
- Meadows, J.R.S.; Cemal, I.; Karaca, O.; Gootwine, E.; Kijas, J. Five Ovine Mitochondrial Lineages Identified from Sheep Breeds of the Near East. Genetics 2006, 175, 1371–1379. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Lischer, H.E.L.; Excoffier, L. PGDSpider: An automated data conversion tool for connecting population genetics and genomics programs. Bioinformatics 2011, 28, 298–299. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Boil. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinform. 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Trifinopoulos, J.; Nguyen, L.-T.; Von Haeseler, A.; Minh, B. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Boil. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Felsenstein, J. Phylogenies and the Comparative Method. Am. Nat. 1985, 125, 1–15. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, A.M.; Cabezas, M.P.; Tavares, A.I.; Xavier, R.; Branco, M. tcsBU: A tool to extend TCS network layout and visualization. Bioinformatics 2015, 32, 627–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Bentley, P. Immune Memory and Gene Library Evolution in the Dynamic Clonal Selection Algorithm. Genet. Program. Evolvable Mach. 2004, 5, 361–391. [Google Scholar] [CrossRef]
- Horsburgh, K.A.; Rhines, A. Genetic characterization of an archaeological sheep assemblage from South Africa’s Western Cape. J. Archaeol. Sci. 2010, 37, 2906–2910. [Google Scholar] [CrossRef]
- Tapio, M.; Marzanov, N.; Ozerov, M.; Ćinkulov, M.; Gonzarenko, G.; Kiselyova, T.; Murawski, M.; Viinalass, H.; Kantanen, J. Sheep Mitochondrial DNA Variation in European, Caucasian, and Central Asian Areas. Mol. Boil. Evol. 2006, 23, 1776–1783. [Google Scholar] [CrossRef]
- Resende, A.; Gonçalves, J.; Muigai, A.W.T.; Pereira, F. Mitochondrial DNA variation of domestic sheep (Ovis aries) in Kenya. Anim. Genet. 2016, 47, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, S.K., Jr.; Kolte, A.P.; Kumar, S. Extensive Variation and Sub-Structuring in Lineage A mtDNA in Indian Sheep: Genetic Evidence for Domestication of Sheep in India. PLoS ONE 2013, 8, e77858. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J.; Phua, S.H. Variation in the control region sequence of the sheep mitochondrial genome. Anim. Genet. 2009, 27, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Delgado, J.V.; Fresno, M.R.; Camacho, M.E.; Rodero, E.; Barba, C. Origin and influences of Canarian Sheep. Arch. Zootec. 1998, 47, 511–516. [Google Scholar]
- Primo, A.T. El ganado bovino ibérico en las Américas: 500 años después. Arch. Zootec. 1992, 41, 13. [Google Scholar]
- Mogren, E.W. New Mexico’s Spanish Livestock Heritage: Four Centuries of Animals, Land, and People. By William W. Dunmire. Environ. Hist. 2014, 19, 784–786. [Google Scholar] [CrossRef]
- Mason, I.L. Razas indígenas de ovinos y caprinos en América Latina. In Animal Production and Health; Gelman, B.M.-H.a.J., Ed.; FAO: Roma, Italy, 1981; pp. 132–140. [Google Scholar]
- Muñoz, C.E.; Vazquez, F.B. Importancia del merino en el desarrollo del ovino en América. Feagas 2000, 18, 88–96. [Google Scholar]
- Aguilar-Martínez, C.U.; Berruecos-Villalobos, J.M.; Espinoza-Gutiérrez, B.; Segura-Correa, J.C.; Valencia-Méndez, J.; Roldán-Roldán, A. Origen, historia y situación actual de la oveja pelibuey en México. Trop. Subtrop. Agroecosyst. 2017, 20, 429–439. [Google Scholar]
- Del Arco Aguilar, M.D.C.; Del Arco Aguilar, M.M.; Armas, E.A.; Hopf, M. Estudio de los restos vegetales de la cueva de Don Gaspar y algunas anotaciones sobre la agricultura prehistórica de Tenerife. Investig. Arqueol. 1990, 2, 13–30. [Google Scholar]
- Vivas Ascue, N.J. Diversidad genética de ovinos criollos colombianos. Master’s Thesis, Universidad Nacional de Colombia—Sede Palmira, Valle del Cauca, Colombia, 2013. [Google Scholar]
- Jiménez-Robayo, L.M.; Triana-Arévalo, L.F.; Cortes Gardyn, O.; Leal-Gutiérrez, J.D.; Sánchez-Isaza, C.A. Estructura genética y caracterización molecular de ovinos criollos de pelo Colombiano utilizando 14 microsatélites autosómicos. Actas Iberoam. Conserv. Anim. 2017, 10, 128–134. [Google Scholar]
- Ferrando, A.; Casas, M.; Marmi, J.; Parés, P.M.; Jordana, J. Variabilidad del ADN mitocondrial en cinco razas ovinas pirenaicas. In Proceedings of the AIDA (2009), XIII Jornadas sobre Producción Animal, Tomo I, Zaragoza, Spain, 12–13 May 2009; pp. 48–50. [Google Scholar]
- Campos, E.; Cuéllar, J.; Salvador, O.; García-Trejo, E.A.; Pereira, F. The genetic diversity and phylogeography of Mexican domestic sheep. Small Rumin. Res. 2020, 187, 106109. [Google Scholar] [CrossRef]
- Adhena, M.H. Genetic Diversity of North Ethiopian Indigenous Sheep Populations Using Mitochondrial DNA; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2018. [Google Scholar]
- Gornas, N.; Weimann, C.; El Hussien, A.; Erhardt, G. Genetic characterization of local Sudanese sheep breeds using DNA markers. Small Rumin. Res. 2011, 95, 27–33. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breed | Acronym | Type | N (f) | Department (region) |
|---|---|---|---|---|
| Ethiopian | OPCE | Hair sheep | 23 (6) | Córdoba, Cesar, Atlántico, Magdalena (Caribbean coast) |
| Sudan | OPCS | Hair sheep | 23 (5) | Córdoba, Cesar, Atlántico (Caribbean coast) |
| Pelibuey | OPCP | Hair sheep | 14 (1) | Valle del Cauca (Pacific Coast, Colombian southwest) |
| Wool Creole | CL | Wool | 18 (8) | Boyacá; Andean region and Nariño (Colombian southwest) |
| Wayúu | OPCW | Hair sheep | 11 (7) | Guajira Peninsula (Caribbean coast) |
| Breed | n | S | NS | Pi | K | h-SD(h) | π-SD(π) |
|---|---|---|---|---|---|---|---|
| CL | 18 | 28 | 9 | 19 | 7.85 | 0.93 ± 0.04 | 0.011 ± 0.002 |
| OPCE | 23 | 23 | 8 | 15 | 6 | 0.87 ± 0.04 | 0.009 ± 0.001 |
| OPCS | 23 | 42 | 17 | 25 | 4.36 | 0.92 ± 0.04 | 0.0014 ± 0.002 |
| OPCP | 14 | 16 | 1 | 15 | 4.60 | 0.59 ± 0.14 | 0.0070 ± 0.002 |
| OPCW | 11 | 16 | 9 | 7 | 4.25 | 0.81 ± 0.1 | 0.006 ± 0.001 |
| TOTAL | 89 | 53 | 17 | 36 | 7.86 | 0.92 ± 0.01 | 0.01 ± 0.001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Revelo, H.A.; López-Alvarez, D.; Landi, V.; Rizzo, L.; Alvarez, L.A. Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes. Animals 2020, 10, 1594. https://doi.org/10.3390/ani10091594
Revelo HA, López-Alvarez D, Landi V, Rizzo L, Alvarez LA. Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes. Animals. 2020; 10(9):1594. https://doi.org/10.3390/ani10091594
Chicago/Turabian StyleRevelo, Herman Alberto, Diana López-Alvarez, Vincenzo Landi, Lauden Rizzo, and Luz Angela Alvarez. 2020. "Mitochondrial DNA Variations in Colombian Creole Sheep Confirm an Iberian Origin and Shed Light on the Dynamics of Introduction Events of African Genotypes" Animals 10, no. 9: 1594. https://doi.org/10.3390/ani10091594