
Massive Infection of Lungs with Exo-Erythrocytic Meronts in European Robin Erithacus rubecula during Natural Haemoproteus attenuatus Haemoproteosis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Sample Collection
2.2. Blood and Histological Samples
2.3. DNA Extraction, PCR and Sequencing
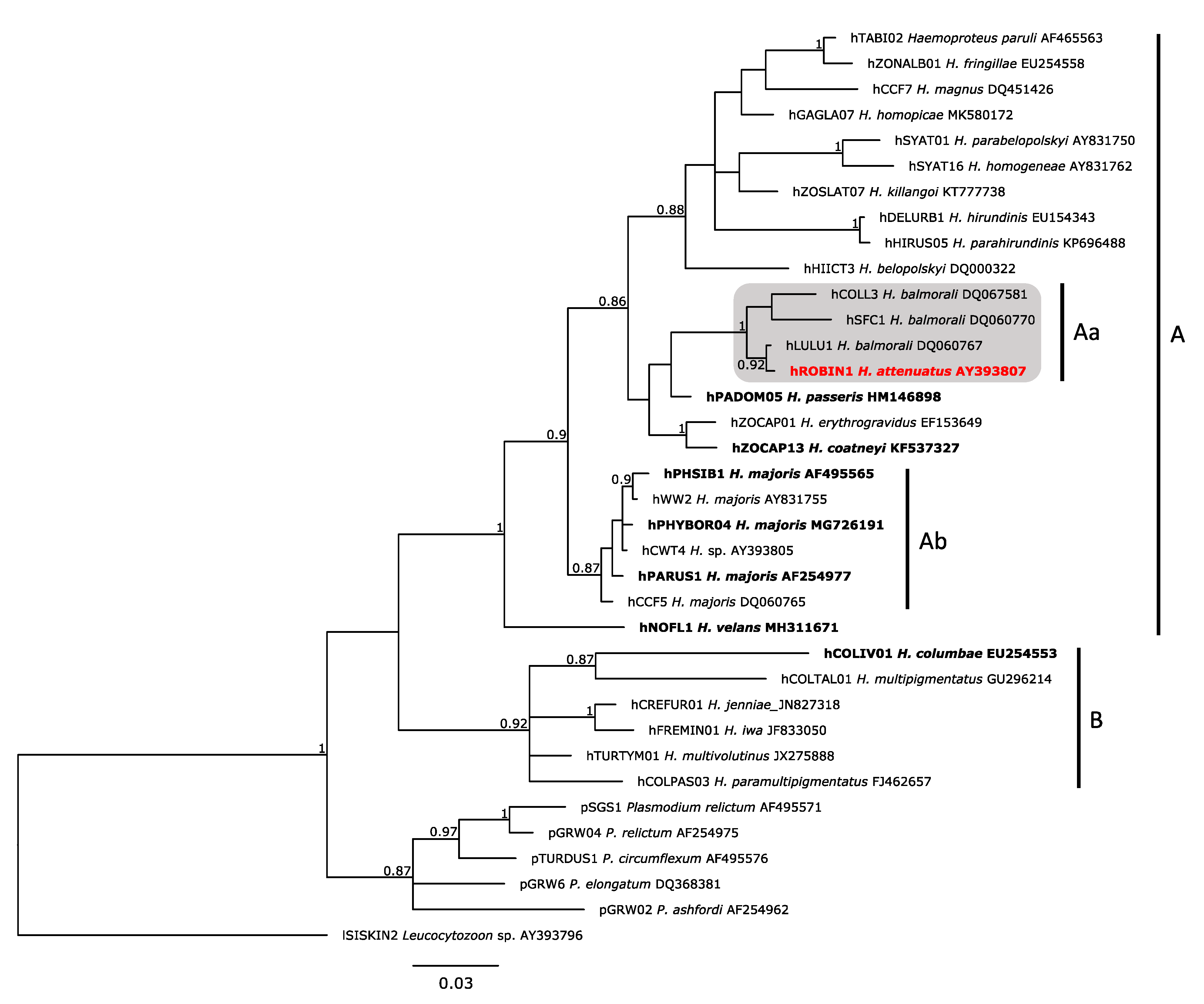
2.4. Phylogenetic Analysis
3. Results
3.1. Molecular Analysis
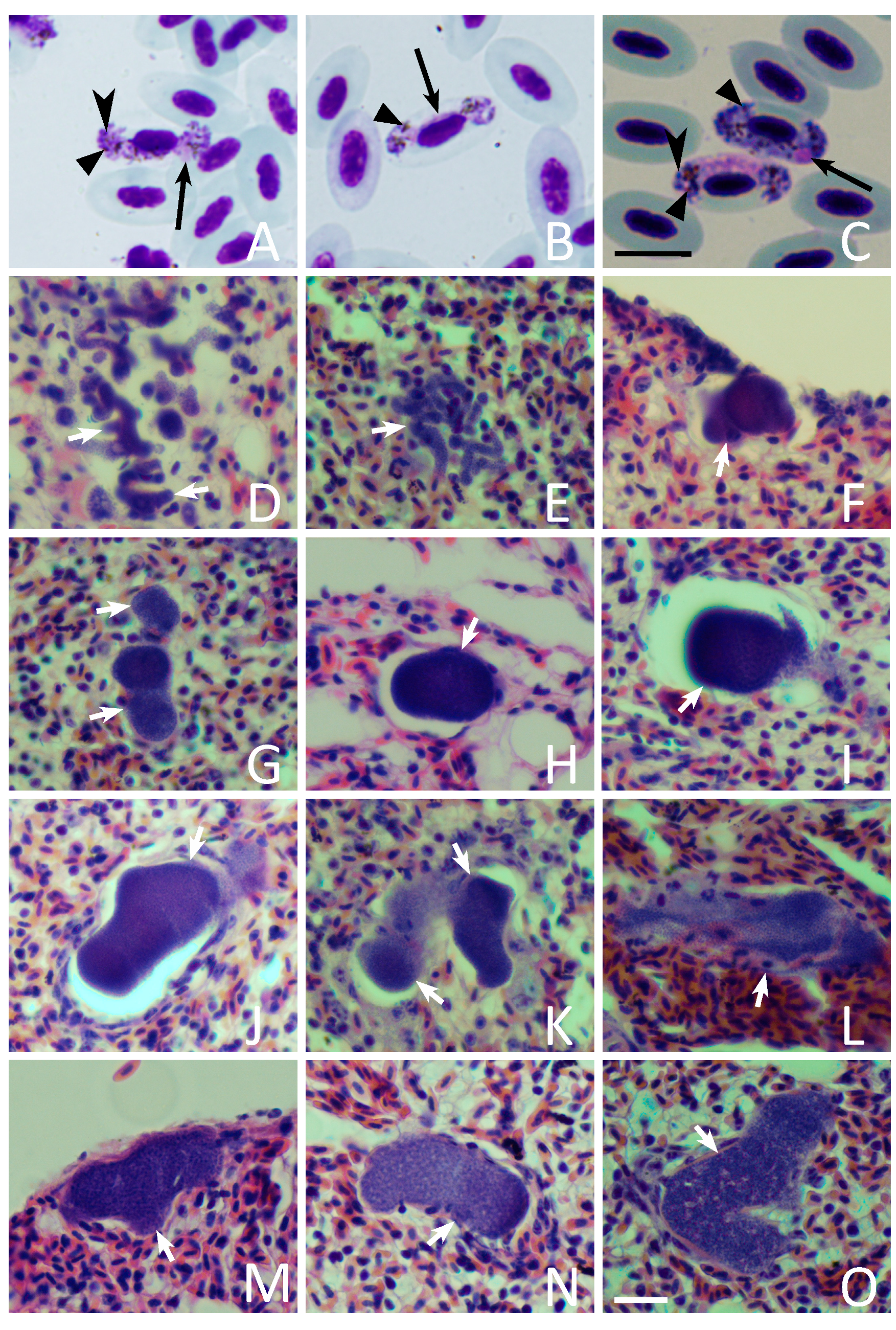
3.2. Blood Stages
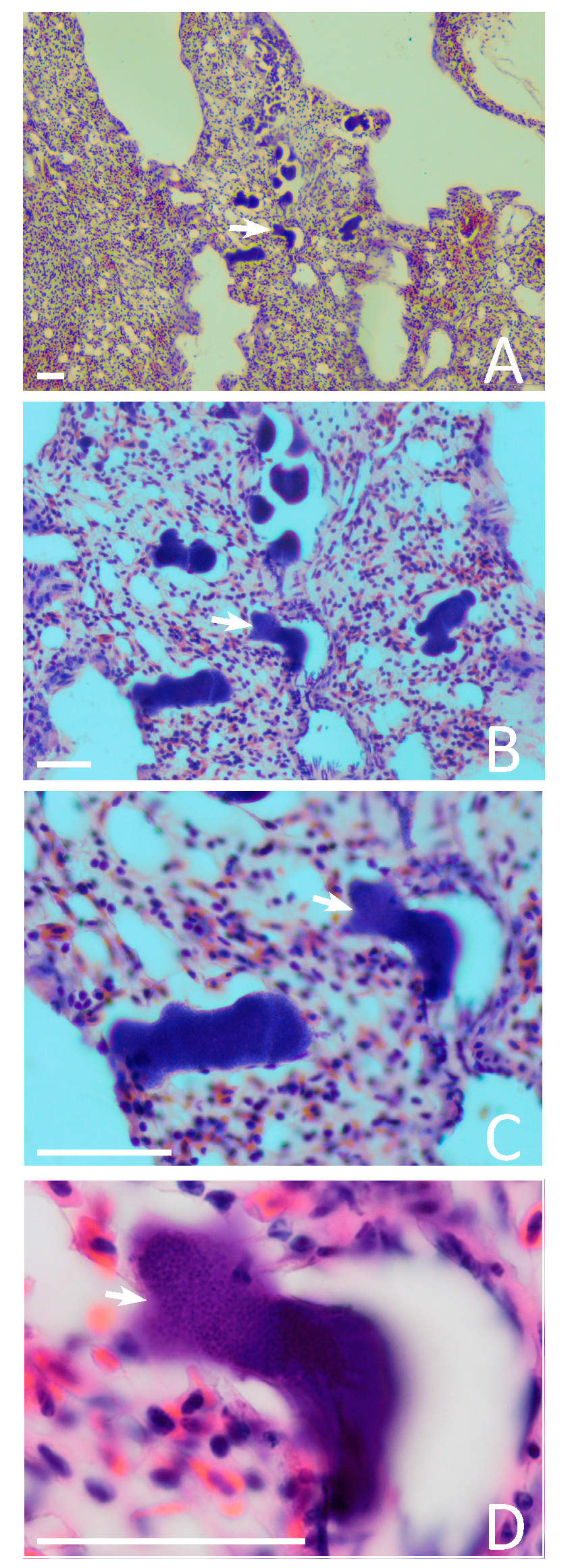
3.3. Exo-Erythrocytic Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005; ISBN 978-0-415-30097-1. [Google Scholar]
- Quillfeldt, P.; Arriero, E.; Martínez, J.; Masello, J.F.; Merino, S. Prevalence of blood parasites in seabirds—A review. Front. Zool. 2011, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Valkiūnas, G.; Iezhova, T.A. Exo-erythrocytic development of avian malaria and related haemosporidian parasites. Malar. J. 2017, 16, 101. [Google Scholar] [CrossRef] [Green Version]
- Cardona, C.J.; Ihejirika, A.; McClellan, L. Haemoproteus lophortyx infection in bobwhite quail. Avian Dis. 2002, 46, 249–255. [Google Scholar] [CrossRef]
- Ferrell, S.T.; Snowden, K.; Marlar, A.B.; Garner, M.; Lung, N.P. Fatal hemoprotozoal infections in multiple avian species in a zoological park. J. Zoo Wildl. Med. 2007, 38, 309–316. [Google Scholar] [CrossRef]
- Donovan, T.A.; Schrenzel, M.; Tucker, T.A.; Pessier, A.P.; Stalis, I.H. Hepatic hemorrhage, hemocoelom, and sudden death due to Haemoproteus infection in passerine birds: Eleven cases. J. Vet. Diagn. Investig. 2008, 20, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Pacheco, M.A.; Escalante, A.A.; Garner, M.M.; Bradley, G.A.; Aguilar, R.F. Haemosporidian infection in captive masked bobwhite quail (Colinus virginianus ridgwayi), an endangered subspecies of the northern bobwhite quail. Vet. Parasitol. 2011, 182, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannell, B.L.; Krasnec, K.V.; Campbell, K.; Jones, H.I.; Miller, R.D.; Stephens, N. The pathology and pathogenicity of a novel Haemoproteus spp. infection in wild little penguins (Eudyptula minor). Vet. Parasitol. 2013, 197, 74–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palinauskas, V.; Iezhova, T.A.; Križanauskienė, A.; Markovets, M.Y.; Bensch, S.; Valkiūnas, G. Molecular characterization and distribution of Haemoproteus minutus (Haemosporida, Haemoproteidae): A pathogenic avian parasite. Parasitol. Int. 2013, 62, 358–363. [Google Scholar] [CrossRef] [PubMed]
- Duc, M.; Ilgūnas, M.; Valkiūnas, G. Patterns of Haemoproteus majoris (Haemosporida, Haemoproteidae) megalomeront development. Acta Trop. 2020, 212, 105706. [Google Scholar] [CrossRef]
- Duc, M.; Ilgūnas, M.; Kubiliūnaitė, M.; Valkiūnas, G. First report of Haemoproteus (Haemosporida, Haemoproteidae) megalomeronts in the brain of an avian host, with description of megalomerogony of Haemoproteus pastoris, the blood parasite of the Common starling. Animals 2021, 11, 2824. [Google Scholar] [CrossRef] [PubMed]
- Shokrani, H.; Norouzian, H.; Dezfoulian, O. Exo-Erythrocytic stages of Haemoproteus sp. in Common buzzard (Buteo buteo): A histopathological and molecular study. Int. J. Parasitol. Parasites Wildl. 2021, 16, 64–69. [Google Scholar] [CrossRef]
- Iezhova, T. Comparative Morphology of Haemoproteids (Haemosporidia, Haemoproteidae) of the Palearctic birds. Ph.D. Thesis, Institute of Ecology, Vilnius, Lithuania, 1994. (In Russian). [Google Scholar]
- Lillie, R.D. Histopathologic Technic and Practical Histochemistry, 3rd ed.; McGraw-Hill Book Co.: New York, NY, USA, 1965; ISBN 978-0-07-037861-2. [Google Scholar]
- Godfrey, R.D.; Fedynich, A.M.; Pence, D.B. Quantification of hematozoa in blood smears. J. Wildl. Dis. 1987, 23, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989; ISBN 978-1-936113-41-5. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Örjan, Ö.; Hannson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. R. Soc. Lond. B Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [Green Version]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES science gateway for inference of large phylogenetic trees. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 1–8. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in bayesian phylogenetics using tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree: Tree Figure Drawing Tool; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2006. [Google Scholar]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis; The University of British Columbia, Vancouver, Canada and Oregon State University: Corvallis, OR, USA, 2018. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and high-performance computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Bukauskaitė, D.; Iezhova, T.A.; Ilgūnas, M.; Valkiūnas, G. High susceptibility of the laboratory-reared biting midges Culicoides nubeculosus to Haemoproteus infections, with review on Culicoides species that transmit avian haemoproteids. Parasitology 2019, 146, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Ellis, V.A.; Huang, X.; Westerdahl, H.; Jönsson, J.; Hasselquist, D.; Neto, J.M.; Nilsson, J.; Nilsson, J.; Hegemann, A.; Hellgren, O.; et al. Explaining prevalence, diversity and host specificity in a community of avian haemosporidian parasites. Oikos 2020, 129, 1314–1329. [Google Scholar] [CrossRef]
- Dimitrov, D.; Zehtindjiev, P.; Bensch, S. Genetic diversity of avian blood parasites in SE Europe: Cytochrome b lineages of the genera Plasmodium and Haemoproteus (Haemosporida) from Bulgaria. Acta Parasitol. 2010, 55. [Google Scholar] [CrossRef]
- Drovetski, S.V.; Aghayan, S.A.; Mata, V.A.; Lopes, R.J.; Mode, N.A.; Harvey, J.A.; Voelker, G. Does the niche breadth or trade-off hypothesis explain the abundance-occupancy relationship in avian Haemosporidia? Mol. Ecol. 2014, 23, 3322–3329. [Google Scholar] [CrossRef]
- Garcia-Longoria, L.; Marzal, A.; de Lope, F.; Garamszegi, L. Host-parasite interaction explains variation in the prevalence of avian haemosporidians at the community level. PLoS ONE 2019, 14, e0205624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellgren, O.; Waldenström, J.; Peréz-Tris, J.; Szöll Ösi, E.; Hasselquist, D.; Krizanauskiene, A.; Ottosson, U.; Bensch, S. Detecting shifts of transmission areas in avian blood parasites—A phylogenetic approach. Mol. Ecol. 2007, 16, 1281–1290. [Google Scholar] [CrossRef]
- Mata, V.A.; da Silva, L.P.; Lopes, R.J.; Drovetski, S.V. The Strait of Gibraltar poses an effective barrier to host-specialised but not to host-generalised lineages of avian Haemosporidia. Int. J. Parasitol. 2015, 45, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Santiago-Alarcon, D.; MacGregor-Fors, I.; Kühnert, K.; Segelbacher, G.; Schaefer, H.M. Avian haemosporidian parasites in an urban forest and their relationship to bird size and abundance. Urban Ecosyst. 2016, 19, 331–346. [Google Scholar] [CrossRef]
- Stanković, D.; Jönsson, J.; Raković, M. Diversity of avian blood parasites in wild passerines in Serbia with special reference to two new lineages. J. Ornithol. 2019. [Google Scholar] [CrossRef]
- Ciloglu, A.; Ergen, A.G.; Inci, A.; Dik, B.; Duzlu, O.; Onder, Z.; Yetismis, G.; Bensch, S.; Valkiūnas, G.; Yildirim, A. Prevalence and genetic diversity of avian haemosporidian parasites at an intersection point of bird migration routes: Sultan Marshes National Park, Turkey. Acta Trop. 2020, 210, 105465. [Google Scholar] [CrossRef]
- Hellgren, O. The occurrence of haemosporidian parasites in the Fennoscandian bluethroat (Luscinia svecica) population. J. Ornithol. 2005, 146, 55–60. [Google Scholar] [CrossRef]
- Nilsson, E.; Taubert, H.; Hellgren, O.; Huang, X.; Palinauskas, V.; Markovets, M.Y.; Valkiūnas, G.; Bensch, S. Multiple cryptic species of sympatric generalists within the avian blood parasite Haemoproteus majoris. J. Evol. Biol. 2016, 29, 1812–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valkiūnas, G.; Kazlauskienė, R.; Bernotienė, R.; Palinauskas, V.; Iezhova, T.A. Abortive long-lasting sporogony of two Haemoproteus species (Haemosporida, Haemoproteidae) in the mosquito Ochlerotatus cantans, with perspectives on haemosporidian vector research. Parasitol. Res. 2013, 112, 2159–2169. [Google Scholar] [CrossRef]
- Sibley, L.D.; Werner, J.K. Susceptibility of Pekin and Muscovy ducks to Haemoproteus nettionis. J. Wildl. Dis. 1984, 20, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Anschütz, G. Ueber den entwickelungsgang des “Haemoproteus orizivorae” nov. spec. Zent. Für Bakteriol. Mikrobiol. Hyg. 1 Abt Orig. B 1909, 51, 654–659. [Google Scholar]
- Peirce, M.A. Haemoproteus balearicae sp. nov., from crowned cranes, Balearica pavonina pavonina and B. pavonina gibbericeps. Bull. Epizoot. Dis. Afr. 1973, 21, 467–475. [Google Scholar] [PubMed]
- Khan, R.A.; Fallis, A.M. Endogenous stages of Parahaemoproteus fringillae (Labbé, 1894) and Leucocytozoon fringillinarum Woodcock, 1910. Can. J. Zool. 1969, 47, 37–39. [Google Scholar] [CrossRef]
- Garnham, P.; Duggan, A. Catalogue of the Garnham Collection of Malaria Parasites and Other Haemosporidia; The Wellcome Trust: London, UK, 1986; ISBN 978-0-521-35725-8. [Google Scholar]
- Valkiūnas, G.; Liutkevičius, G.; Iezhova, T.A. Complete development of three species of Haemoproteus (Haemosporida, Haemoproteidae) in the biting midge Culicoides impunctatus (Diptera, Ceratopogonidae). J. Parasitol. 2002, 88, 864–868. [Google Scholar] [CrossRef]
- Žiegytė, R.; Markovets, M.Y.; Bernotienė, R.; Mukhin, A.; Iezhova, T.A.; Valkiūnas, G.; Palinauskas, V. The widespread biting midge Culicoides Impunctatus (Ceratopogonidae) is susceptible to infection with numerous Haemoproteus (Haemoproteidae) species. Parasit. Vectors 2017, 10, 397. [Google Scholar] [CrossRef]
- Garvin, M.C.; Homer, B.L.; Greiner, E.C. Pathogenicity of Haemoproteus danilewskyi, Kruse, 1890, in Blue Jays (Cyanocitta cristata). J. Wildl. Dis. 2003, 39, 161–169. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.R. Haemoproteus palumbis sp. nov. (Sporozoa, Haemosporina) of the English wood-pigeon Columba p. palumbus. J. Protozool. 1966, 13, 515–519. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasit. Vectors 2019, 12, 40. [Google Scholar] [CrossRef]
- Ilgūnas, M.; Romeiro Fernandes Chagas, C.; Bukauskaitė, D.; Bernotienė, R.; Iezhova, T.; Valkiūnas, G. The life-cycle of the avian haemosporidian parasite Haemoproteus majoris, with emphasis on the exoerythrocytic and sporogonic development. Parasit. Vectors 2019, 12, 516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Himmel, T.; Harl, J.; Kübber-Heiss, A.; Konicek, C.; Fernández, N.; Juan-Sallés, C.; Ilgūnas, M.; Valkiūnas, G.; Weissenböck, H. Molecular probes for the identification of avian Haemoproteus and Leucocytozoon parasites in tissue sections by chromogenic in situ hybridization. Parasit. Vectors 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paperna, I.; GilI, H. Schizogonic stages of Haemoproteus from Wenyon’s Baghdad sparrows are also found in Passer domesticus biblicus in Israel. Parasitol. Res. 2003, 91, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Farmer, J.N. Gizzard lesions associated with Haemoproteus sacharovi infections of pigeons. Proc. Iowa Acad. Sci. 1964, 71, 537–542. [Google Scholar]
- Miltgen, F.; Landau, I.; Ratanaworabhan, N.; Yenbutra, S. Parahaemoproteus desseri n. sp.; Gamétogonie et schizogonie chez l’hôte naturel: Psittacula roseata de Thaïlande, et sporogonie expérimentale chez Culicoides nubeculosus. Ann. Parasitol. Hum. Comp. 1981, 56, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.T.; Greiner, E.C.; Forrester, D.J. Pre-erythrocytic development and associated host responses to Haemoproteus meleagridis (Haemosporina: Haemoproteidae) in experimentally infected domestic turkeys. J. Protozool. 1986, 33, 375–381. [Google Scholar] [CrossRef]
- Atkinson, C.T.; Forrester, D.J.; Greiner, E.C. Pathogenicity of Haemoproteus meleagridis (Haemosporina: Haemoproteidae) in experimentally infected domestic turkeys. J. Parasitol. 1988, 74, 228. [Google Scholar] [CrossRef]
- Burtikashvili, L. New species of blood parasite, Haemoproteus zasukhini sp. nov. (Sporozoa, Haemoproteidae) from the House sparrow (Passer domesticus L.). Soobsh Akad Nauk Gruz SSR 1973, 72, 697–700. (In Russian) [Google Scholar]
- Burtikashvili, L. Blood Parasites of Wild Birds in Georgia; Metsniereba: Tbilisi, Georgia, 1978. (In Russian) [Google Scholar]
- Peirce, M.A. Haemoproteid parasites of Passer spp. Parasitology 1976, 73, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Wenyon, C. Protozoology: A Manual for Medical Men, Veterinarians and Zoologists, Etc; Bailliére Tindal & Cox: London, UK, 1926; ISBN 978-0-266-92218-6. [Google Scholar]
- Earle, R.A.; Bastianello, S.S.; Bennett, G.F.; Krecek, R.C. Histopathology and morphology of the tissue stages of Haemoproteus columbae causing mortality in Columbiformes. Avian Pathol. 1993, 22, 67–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammed, A. Systematic and Experimental Studies on Protozoan Blood Parasites of Egyptian Birds; Cairo University Press: Cairo, Egypt, 1958. [Google Scholar]
- Peirce, M.A.; Lederer, R.; Adlard, R.D.; O’Donoghue, P.J. Pathology associated with endogenous development of haematozoa in birds from Southeast Queensland. Avian Pathol. 2004, 33, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.E.; Mohammed, A.-H.H. Schizogony in Haemoproteus columbae Kruse. J. Protozool. 1977, 24, 389–393. [Google Scholar] [CrossRef] [PubMed]
- Acton, H.; Knowles, R. Studies on the halteridium parasite of the pigeon, Haemoproteus columbae Celli and San Felice. Indian J. Med. Res. 1914, 1, 663–690. [Google Scholar]
- Aragão, H. Über den entwicklungsgang and die übertragung von Haemoproteus columbae. Arch. Protistenkd. 1908, 12, 154–167. [Google Scholar]
- Huff, C. Schizogony and gametocyte development in Leucocytozoon simondi, and comparison with Plasmodium and Haemoproteus. J. Infect. Dis. 1942, 71, 18–32. [Google Scholar] [CrossRef]
- Linxian, H.; Yuguang, L.; Hongping, J. On sporogony and schizogony of Haemoproteus columbae of pigeon. Acta Zool. Sin. 1989, 35, 306–312. [Google Scholar]
- Mohammed, A.H.H. Studies on the schizogony of Haemoproteus columbae Kruse, 1890. Proc. Egypt. Acad. Sci. 1967, 19, 37–46. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sample No. | Age | Parasitemia (%) | Meronts in Lung Tissue | Meront Length (µm), Min–Max (n) | Enlarged Spleen | Darkened | Figure | |
|---|---|---|---|---|---|---|---|---|
| Spleen | Liver | |||||||
| 1 | Juvenile | 0.8 | + | 17.1–49.8 (7) | − | + | − | |
| 2 | Juvenile | 0.95 | − | − | + | + | − | |
| 3 | Juvenile | 1.85 | + | 5.8–77.58 (25) | − | − | − | Figure 2D,H,I and Figure 3A–C |
| 4 | Juvenile | 2.9 | + | 40.4–99.8 (5) | + | + | + | Figure 2M–O |
| 5 | Juvenile | 23.6 | + | 20.6–28.5 (2) | − | + | + | |
| 6 | Adult | 10.45 | + | 108.7 (1) | − | + | − | |
| 7 | Unknown | 26.5 | + | 9.46–94.27 (14) | − | − | − | Figure 2E–G,J–L |
| Host Order | Host Family | Host Species | Location a | Reference |
|---|---|---|---|---|
| Diptera | Ceratopogonidae | Culicoides festivipennisb | Lithuania | Bernotienė et al., unpublished d |
| C. obsoletusb | Lithuania | Bernotienė et al., unpublished d | ||
| C. nubeculosus | Lithuania | [29] | ||
| Coraciiformes | Alcedinidae | Alcedo atthisb | Spain | Rojo et al., unpublished e |
| Passeriformes | Certhiidae | Certhia familiarisb | Sweden | [30] |
| Acrocephalidae | Acrocephalus schoenobaenusb | Sweden | [30] | |
| Sylviidae | Sylvia communisb | Sweden | Hellgren et al., unpublished d | |
| Muscicapidae | Erithacus rubeculac | Bulgaria, Germany, Lithuania, Morocco, NWA, NWI, Portugal, Russia, Serbia, Spain, Sweden | [30,31,32,33,34,35,36,37] | |
| Luscinia lusciniac | Lithuania, Russia, Sweden, Turkey, WGC | [30,32,34,38,39,40,41] | ||
| L. megarhynchosc | Bulgaria, Germany, TRC | [31,32,34] | ||
| Saxicola rubetra | Nigeria, Sweden, TRC | [30,32,34] | ||
| Turdidae | Turdus merulab | TRC | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hernández-Lara, C.; Duc, M.; Ilgūnas, M.; Valkiūnas, G. Massive Infection of Lungs with Exo-Erythrocytic Meronts in European Robin Erithacus rubecula during Natural Haemoproteus attenuatus Haemoproteosis. Animals 2021, 11, 3273. https://doi.org/10.3390/ani11113273
Hernández-Lara C, Duc M, Ilgūnas M, Valkiūnas G. Massive Infection of Lungs with Exo-Erythrocytic Meronts in European Robin Erithacus rubecula during Natural Haemoproteus attenuatus Haemoproteosis. Animals. 2021; 11(11):3273. https://doi.org/10.3390/ani11113273
Chicago/Turabian StyleHernández-Lara, Carolina, Mélanie Duc, Mikas Ilgūnas, and Gediminas Valkiūnas. 2021. "Massive Infection of Lungs with Exo-Erythrocytic Meronts in European Robin Erithacus rubecula during Natural Haemoproteus attenuatus Haemoproteosis" Animals 11, no. 11: 3273. https://doi.org/10.3390/ani11113273