
Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Donor Birds and Examination of Blood Films
2.2. Circadian Rhythm Investigation
2.3. Collection of Filarioid Worms
2.4. DNA Extraction, PCR and Molecular Identification of Parasites
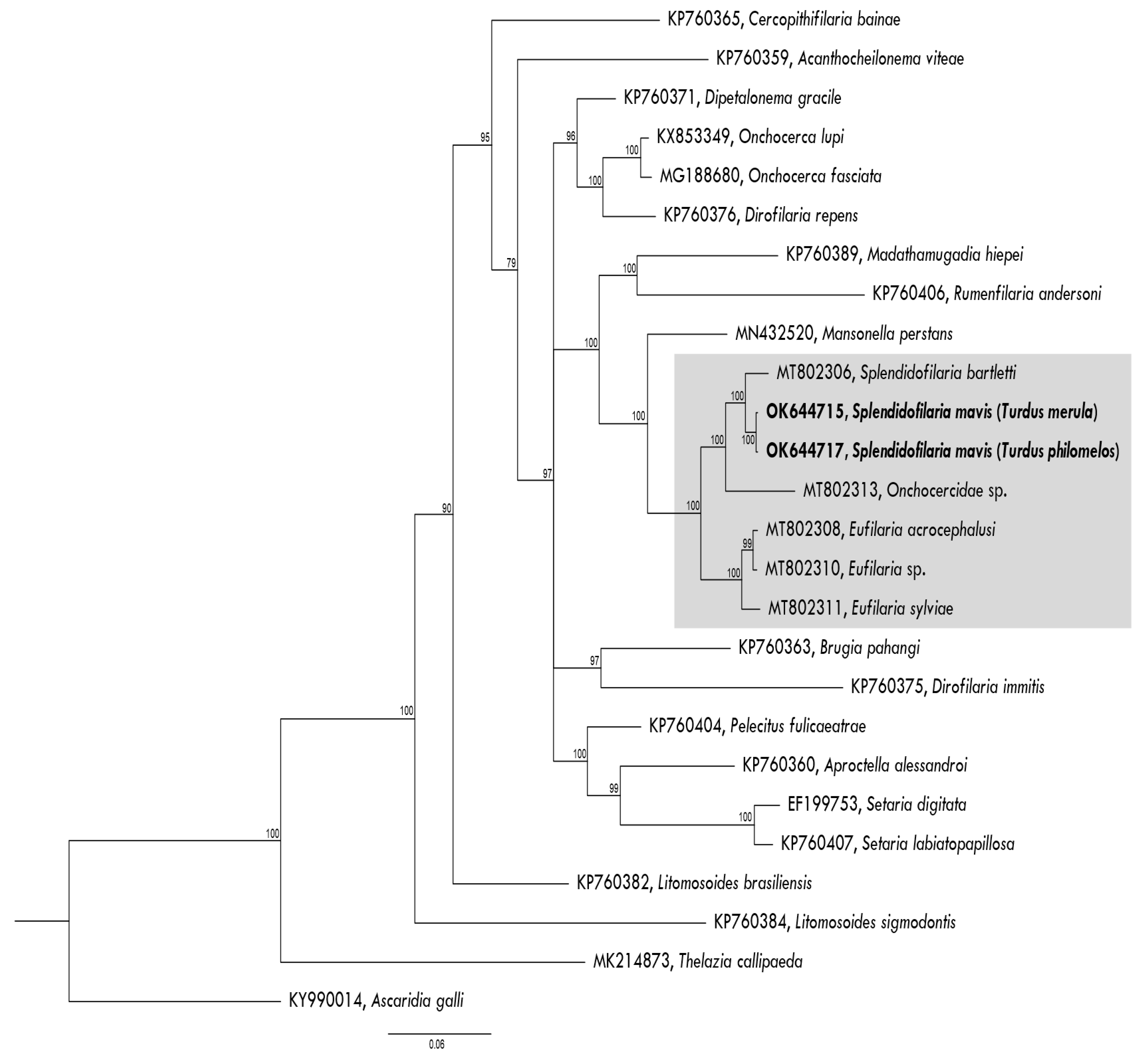
2.5. Plylogenetic Analysis
3. Results
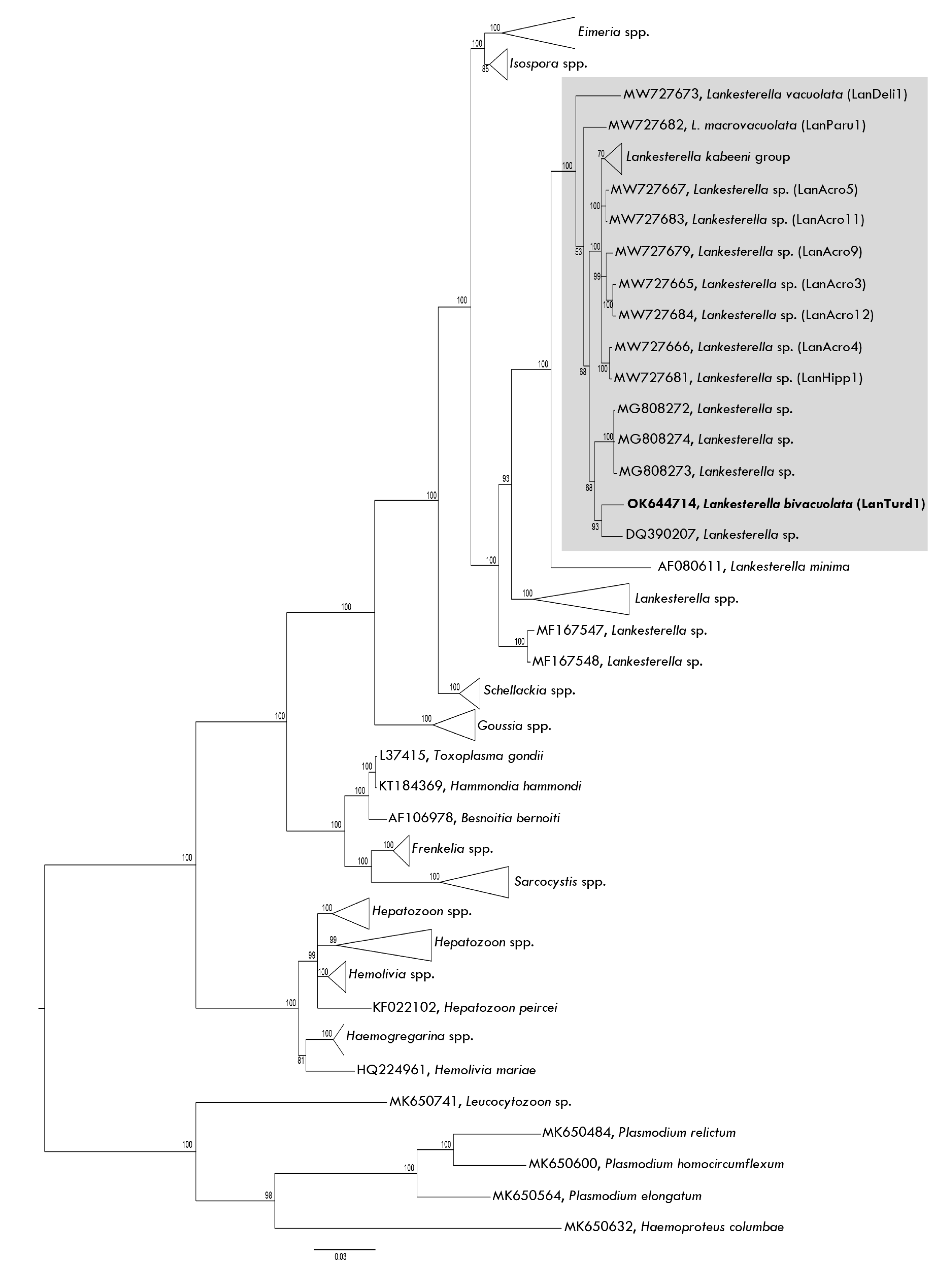
3.1. Parasite Morphological and Molecular Identification
3.1.1. Eurasian Blackbird Infections
3.1.2. Song Thrush Infections
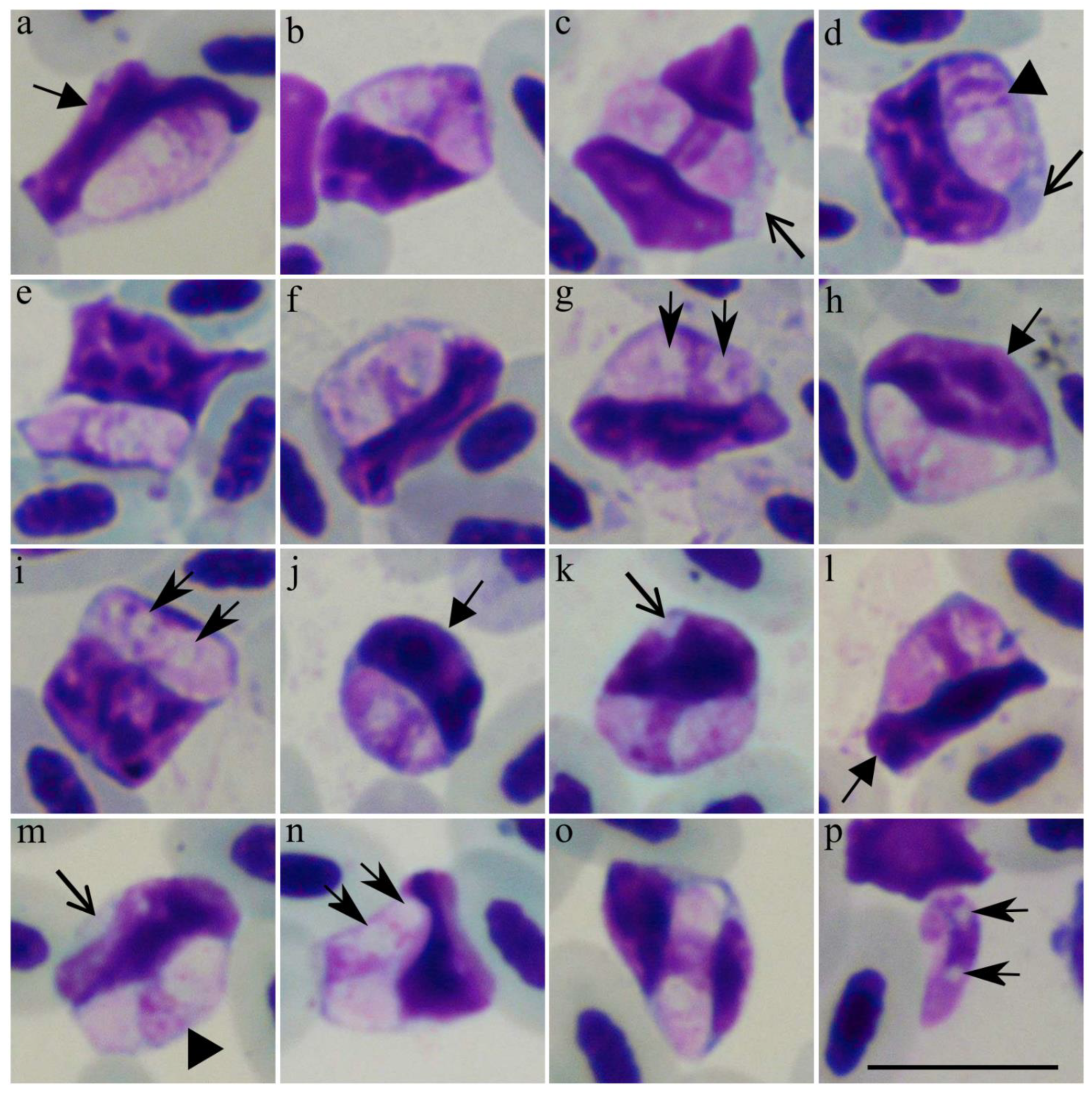
3.2. Description of New Lankesterella Species (Eimeriorina, Apicomplexa)
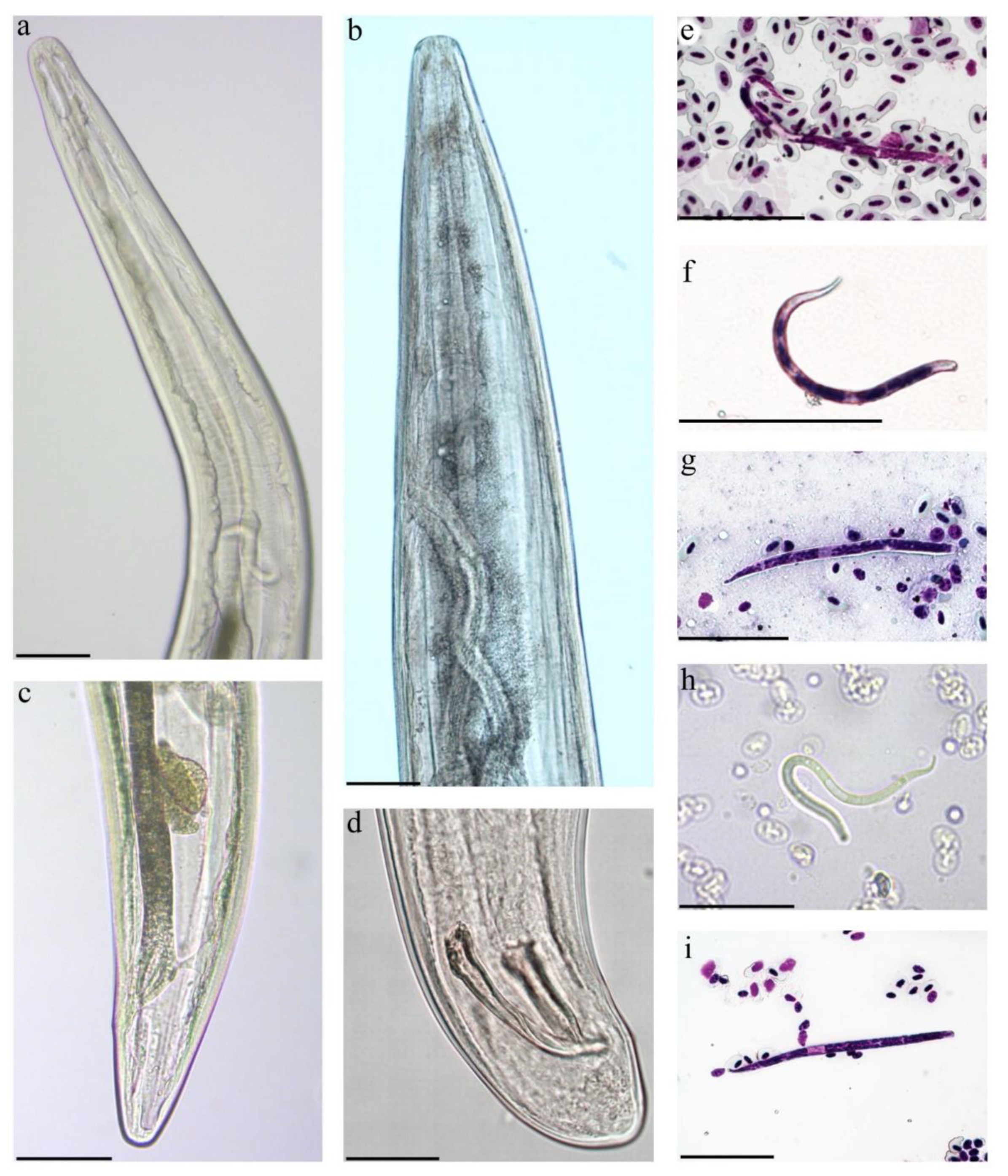
3.3. Description of Splendidofilaria mavis
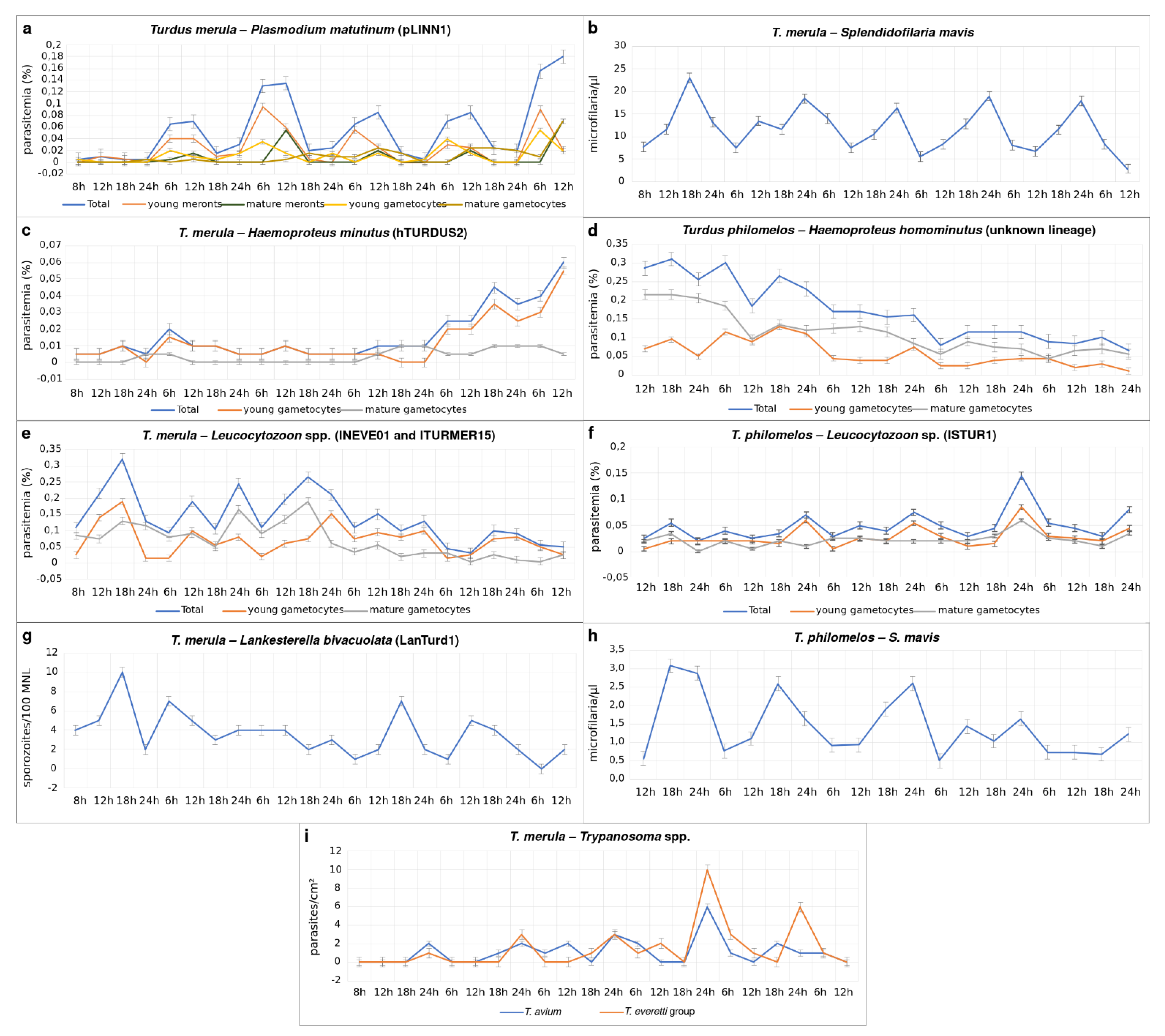
3.4. Circadian Rhythms
3.4.1. Parasites of the Eurasian Blackbird
3.4.2. Parasites of the Song Thrush
4. Discussion
4.1. Description of New Lankesterella Parasite
4.2. Description and Molecular Characterization of Splendidofilaria mavis
4.3. Circadian Rhythms of Blood Parasites during Co-Infections
4.3.1. Plasmodium Infections
4.3.2. Haemoproteus Infections
4.3.3. Leucocytozoon Infections
4.3.4. Lankesterella Infection
4.3.5. Trypanosoma Infections
4.3.6. Microfilariae Infections
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atkinson, C.T.; Thomas, N.; Hunter, B. Parasitic Disease of Wild Birds; John Wiley & Sons: Ames, IA, USA, 2008. [Google Scholar]
- Santiago-Alarcon, D.; Marzal, A. Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Springer: Cham, Switzerland, 2020; ISBN 978-3-030-51632-1. [Google Scholar]
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Binkienė, R.; Chagas, C.R.F.; Bernotienė, R.; Valkiūnas, G. Molecular and Morphological Characterization of Three New Species of Avian Onchocercidae (Nematoda) with Emphasis on Circulating Microfilariae. Parasit. Vectors 2021, 14, 137. [Google Scholar] [CrossRef]
- Chagas, C.R.F.; Binkienė, R.; Ilgūnas, M.; Iezhova, T.; Valkiūnas, G. The Buffy Coat Method: A Tool for Detection of Blood Parasites without Staining Procedures. Parasit. Vectors 2020, 13, 104. [Google Scholar] [CrossRef] [Green Version]
- Clark, N.J.; Wells, K.; Dimitrov, D.; Clegg, S.M. Co-Infections and Environmental Conditions Drive the Distributions of Blood Parasites in Wild Birds. J. Anim. Ecol. 2016, 85, 1461–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, S.; Melo, M.; Covas, R.; Doutrelant, C.; Pereira, H.; de Lima, R.; Loiseau, C. Influence of Land Use and Host Species on Parasite Richness, Prevalence and Co-Infection Patterns. Int. J. Parasitol. 2021, 51, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Van Rooyen, J.; Lalubin, F.; Glaizot, O.; Christe, P. Avian Haemosporidian Persistence and Co-Infection in Great Tits at the Individual Level. Malar. J. 2013, 12, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soares, L.; Ellis, V.A.; Ricklefs, R.E. Co-Infections of Haemosporidian and Trypanosome Parasites in a North American Songbird. Parasitology 2016, 143, 1930–1938. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Iezhova, T.A.; Shapoval, A.P. High Prevalence of Blood Parasites in Hawfinch Coccothraustes Coccothraustes. J. Nat. Hist. 2003, 37, 2647–2652. [Google Scholar] [CrossRef]
- Bernotienė, R.; Palinauskas, V.; Iezhova, T.; Murauskaitė, D.; Valkiūnas, G. Avian Haemosporidian Parasites (Haemosporida): A Comparative Analysis of Different Polymerase Chain Reaction Assays in Detection of Mixed Infections. Exp. Parasitol. 2016, 163, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Ciloglu, A.; Ellis, V.A.; Bernotienė, R.; Valkiūnas, G.; Bensch, S. A New One-Step Multiplex PCR Assay for Simultaneous Detection and Identification of Avian Haemosporidian Parasites. Parasitol. Res. 2019, 118, 191–201. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Iezhova, T.A.; Loiseau, C.; Chasar, A.; Smith, T.B.; Sehgal, R.N.M. New Species of Haemosporidian Parasites (Haemosporida) from African Rainforest Birds, with Remarks on Their Classification. Parasitol. Res. 2008, 103, 1213–1228. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Križanauskienė, A.; Iezhova, T.A.; Hellgren, O.; Bensch, S. Molecular Phylegenetic Analysis of Circumnuclear Hemoproteids (Haemosporida: Haemoproteidae) of Sylviid Birds, with a Description of Haemoproteus Parabelopolskyi Sp. No. J. Parasitol. 2007, 93, 680–687. [Google Scholar] [CrossRef] [PubMed]
- Hawking, F. Circadian and other Rhythms of Parasites. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 1975; Volume 13, pp. 123–182. ISBN 978-0-12-031713-4. [Google Scholar]
- Palinauskas, V.; Žiegytė, R.; Ilgūnas, M.; Iezhova, T.A.; Bernotienė, R.; Bolshakov, C.; Valkiūnas, G. Description of the First Cryptic Avian Malaria Parasite, Plasmodium Homocircumflexum n. Sp., with Experimental Data on Its Virulence and Development in Avian Hosts and Mosquitoes. Int. J. Parasitol. 2015, 45, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Pigeault, R.; Caudron, Q.; Nicot, A.; Rivero, A.; Gandon, S. Timing Malaria Transmission with Mosquito Fluctuations. Evol. Lett. 2018, 2, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Bakker, M.; Helm, B. The Influence of Biological Rhythms on Host–Parasite Interactions. Trends Ecol. Evol. 2015, 30, 314–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rijo-Ferreira, F.; Takahashi, J.S.; Figueiredo, L.M. Circadian Rhythms in Parasites. PLOS Pathog. 2017, 13, e1006590. [Google Scholar] [CrossRef] [PubMed]
- Westwood, M.L.; O’Donnell, A.J.; de Bekker, C.; Lively, C.M.; Zuk, M.; Reece, S.E. The Evolutionary Ecology of Circadian Rhythms in Infection. Nat. Ecol. Evol. 2019, 3, 552–560. [Google Scholar] [CrossRef]
- Anderson, R.C. Nematode Parasites of Vertebrates: Their Development and Transmission; Cabi: Wallingford, UK, 2000. [Google Scholar]
- Dolnik, O.V.; Metzger, B.J.; Loonen, M.J.J.E. Keeping the Clock Set under the Midnight Sun: Diurnal Periodicity and Synchrony of Avian Isospora Parasites Cycle in the High Arctic. Parasitology 2011, 138, 1077–1081. [Google Scholar] [CrossRef] [Green Version]
- Calistri, P.; Iannetti, S.; Danzetta, M.L.; Narcisi, V.; Cito, F.; Di Sabatino, D.; Bruno, R.; Sauro, F.; Atzeni, M.; Carvelli, A.; et al. The Components of ‘One World-One Health’ Approach. Transbound. Emerg. Dis. 2013, 60, 4–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godfrey, R.D.; Fedynich, A.M.; Pence, D.B. Quantification of Hematozoan in Blood Smears. J. Wildl. Dis. 1987, 23, 558–565. [Google Scholar] [CrossRef]
- Hellgren, O.; Waldenström, J.; Bensch, S. A New PCR Assay for Simultaneous Studies of Leucocytozoon, Plasmodium and Haemoproteus from Avian Blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Harl, J.; Preikša, V.; Bukauskaitė, D.; Ilgūnas, M.; Weissenböck, H.; Valkiūnas, G. Lankesterella (Apicomplexa, Lankesterellidae) Blood Parasites of Passeriform Birds: Prevalence, Molecular and Morphological Characterization, with Notes on Sporozoite Persistence In Vivo and Development In Vitro. Animals 2021, 11, 1451. [Google Scholar] [CrossRef] [PubMed]
- Bernotienė, R.; Iezhova, T.A.; Bukauskaitė, D.; Chagas, C.R.F.; Kazak, M.; Valkiūnas, G. Development of Trypanosoma Everetti in Culicoides Biting Midges. Acta Trop. 2020, 210, 105555. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.N. Studies on the Helminth Parasites of Birds in Hyderabad State; Nematoda. J. Helminthol. 1948, 22, 77–92. [Google Scholar] [CrossRef]
- Bartlett, C.M.; Anderson, R.C. Lemdana Wernaarti n.Sp. and Other Filarioid Nematodes from Bubo Virginianus and Asio Otus (Strigiformes) in Ontario, Canada, with a Revision of Lemdana and a Key to Avian Filarioid Genera. Can. J. Zool. 1987, 65, 1100–1109. [Google Scholar] [CrossRef]
- Leiper, R.Th. Description of Filaria Mavis n. Sp. from the Trush. Zoologist 1909, 13, 337–339. [Google Scholar]
- Olsen, O.W.; Braun, C.E. New Species of Splendidofilaria and Chandlerella (Filarioidea: Nematoda), with Keys to the Species, from the Band-Tailed Pigeon (Columba Fasciata Fasciata) in the Rocky Mountain Region. Gt. Basin Nat. 1976, 36, 445–457. [Google Scholar]
- Sambrook, J.; Russel, D.W. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Spring Harbour Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Stunžėnas, V.; Petkevičiūtė, R.; Stanevičiūtė, G. Phylogeny of Sphaerium Solidum (Bivalvia) Based on Karyotype and Sequences of 16S and ITS1 RDNA. Open Life Sci. 2011, 6, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Petkevičiūtė, R.; Stunžėnas, V.; Stanevičiūtė, G. Differentiation of European Freshwater Bucephalids (Digenea: Bucephalidae) Based on Karyotypes and DNA Sequences. Syst. Parasitol. 2014, 87, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 98/98/NT. Nucleic. Acid Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Ronquist, R.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT Online Service: Multiple Sequence Alignment, Interactive Sequence Choice and Visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author: Evolutionary Biology Centre; Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Rambout, A. FigTree: Tree Figure Drawing Tool Version 1.4.0; 2006. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 March 2021).
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Bernotienė, R.; Iezhova, T.A. Molecular Characterization and Distribution of Plasmodium Matutinum, a Common Avian Malaria Parasite. Parasitology 2017, 144, 1726–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, J.R. Biology of the trypanosomes of birds. In Biology of the Kinetoplastida; Academic Press: London, UK; New York, NY, USA, , 1976; Volume 1, pp. 131–174. ISBN 978-0-12-460201-4. [Google Scholar]
- Molyneux, D.H. Trypanosoma Everetti Sp. Nov. a Trypanosome from the Black-Rumped Waxbill Estrilda t. Troglodytes Lichtenstein. Ann. Trop. Med. Parasitol. 1973, 67, 219–222. [Google Scholar] [CrossRef] [PubMed]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Chagas, C.R.F.; Bernotienė, R.; Himmel, T.; Harl, J.; Weissenböck, H.; Iezhova, T.A. Molecular Characterization of Six Widespread Avian Haemoproteids, with Description of Three New Haemoproteus Species. Acta Trop. 2019, 197, 105051. [Google Scholar] [CrossRef] [PubMed]
- Clark, P.; Boardman, W.S.J.; Raidal, S. Atlas of Clinical Avian Hematology; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Bennett, G.F.; Earlé, R.A.; Peirce, M.A. New Species of Avian Hepatozoon (Apicomplexa: Haemogregarinidae) and a Re-Description of Hepatozoon Neophrontis (Todd & Wohlbach, 1912) Wenyon, 1926. Syst. Parasitol. 1992, 23, 183–193. [Google Scholar] [CrossRef]
- Biedrzycka, A.; Kloch, A.; Migalska, M.; Bielański, W. Molecular Characterization of Putative Hepatozoon Sp. from the Sedge Warbler (Acrocephalus Schoenobaenus). Parasitology 2013, 140, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Merino, S.; Martínez, J.; Martínez-de la Puente, J.; Criado-Fornelio, Á.; Tomás, G.; Morales, J.; Lobato, E.; García-Fraile, S. Molecular Characterization of the 18S RDNA Gene of an Avian Hepatozoon Reveals That It Is Closely Related to Lankesterella. J. Parasitol. 2006, 92, 1330–1335. [Google Scholar] [CrossRef] [PubMed]
- Gönnert, R.V. Zur Frage Der Artzugehörigkeit von Filaria Mavis Leiper, 1909. Festschr. Bernhard Nocht Zum 1937, 80, 159–162. [Google Scholar]
- Martínez, J.; Merino, S.; Badás, E.P.; Almazán, L.; Moksnes, A.; Barbosa, A. Hemoparasites and Immunological Parameters in Snow Bunting (Plectrophenax Nivalis) Nestlings. Polar Biol. 2018, 41, 1855–1866. [Google Scholar] [CrossRef]
- Bartlett, C.M. New, Known and Unidentified Species of Eulimdana (Nematoda): Additional Information on Biologically Unusual Filarioids of Charadriiform Birds. Syst. Parasitol. 1992, 23, 209–230. [Google Scholar] [CrossRef]
- Bartlett, C.M.; Anderson, R.C. Filarioid Nematodes (Filarioidea: Onchocercidae) of Corvus Brachyrhynchos Brachyrhynchos Brehm in Southern Ontario, Canada and a Consideration of the Epizootiology of Avian Filariasis. Syst. Parasitol. 1980, 2, 77–102. [Google Scholar] [CrossRef]
- Bartlett, C.M.; Bain, O. New Avian Filarioids (Nematoda: Splendidofilariinae): Dessetfilaria Guianensis Gen. n., Sp. n., Andersonfilaria Africanus Gen. n., Sp. n., and Splendidofilaria Chandenieri Sp. n. Proc. Helminthol. Soc. Wash. 1987, 54, 1–14. [Google Scholar]
- Haas, M.; Baruš, V.; Benedikt, V.; Literák, I. Microfilariae in Birds in the Czech Republic, Including a Note on Adult Nematodes Eufilaria Delicata in a Song Thrush Turdus Philomelos. Parasitol. Res. 2011, 109, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Sonin, M.D. Filariata of Animals and Man and Diseases Caused by Them. 2: Diplotriaenoidea; Osnovy nematodologii; Israel Program for Scientific Translations: Jerusalem, Israel, 1975; ISBN 978-0-7065-1504-6. [Google Scholar]
- Okulewicz, A. Ornithofilaria mavis (Leiper 1909) Gönnert, 1937 in blackbird (Turdus merula L.) in Poland]. Wiad. Parazytol. 1981, 27, 669–671. [Google Scholar] [PubMed]
- Chabaud, A.G.; Golvan, Y. Nouvelle Filaire Parasite Des Grives En France. Ann. Parasitol. Hum. Comparée 1956, 31, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Lopez Caballero, E.J. Incidencia del parasitismo por formas de la superfamilia Filarioidea (Nematoda: Spirurina) en aves del genero Turdus (Passeriformes: Musacapidae). Boletin Estac. Cent. Ecol. 1978, 7, 67–71. [Google Scholar]
- Hauptmanová, K.; Baruš, V.; Literák, I.; Benedikt, V. Haemoproteids and Microfilariae in Hawfinches in the Czech Republic. Helminthologia 2004, 4, 125–133. [Google Scholar]
- Supperer, R. Zwei neue Filarien (s. l.), Eufilaria delicata spec. nov. und Ornithofilaria böhmi spec. nov. aus der Misteldrossel, Turdus viscivorus L. Z. Für Parasitenkd. 1958, 18. [Google Scholar] [CrossRef]
- Fecchio, A.; Chagas, C.R.F.; Bell, J.A.; Kirchgatter, K. Evolutionary Ecology, Taxonomy, and Systematics of Avian Malaria and Related Parasites. Acta Trop. 2020, 204, 105364. [Google Scholar] [CrossRef]
- Southworth, G.C.; Mason, G.; Seed, J.R. Studies on Frog Trypanosomiasis. I. A 24-Hour Cycle in the Parasitemia Level of Trypanosoma Rotatorium in Rana Clamitans from Louisiana. J. Parasitol. 1968, 54, 255. [Google Scholar] [CrossRef]
- Di Cesare, A.; Otranto, D.; Di Giulio, E.; Simonato, G.; Latrofa, M.S.; La Torre, F.; Coccia, G.; Traversa, D. Microfilarial Periodicity of Dirofilaria Repens in Naturally Infested Dogs. Parasitol. Res. 2013, 112, 4273–4279. [Google Scholar] [CrossRef] [PubMed]
- Gooneratne, B.W.M. On Cardiofilaria Nilesi in Experimentally Infected Chickens with a Note on the Morphology and Periodicity of the Microfilariae. J. Helminthol. 1969, 43, 311–317. [Google Scholar] [CrossRef]
- Garnham, P.C.C. Malaria Parasites and Other Haemosporidia; Blackwell Scientific: Oxford, UK, 1966; ISBN 978-0-632-01770-6. [Google Scholar]
- Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.A.; Dinhopl, N.; Nedorost, N.; Weissenbacher-Lang, C.; Weissenböck, H.; Valkiūnas, G. Mortality and Pathology in Birds Due to Plasmodium (Giovannolaia) Homocircumflexum Infection, with Emphasis on the Exoerythrocytic Development of Avian Malaria Parasites. Malar. J. 2016, 15, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palinauskas, V.; Valkiūnas, G.; Bolshakov, C.V.; Bensch, S. Plasmodium Relictum (Lineage P-SGS1): Effects on Experimentally Infected Passerine Birds. Exp. Parasitol. 2008, 120, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Palinauskas, V.; Žiegytė, R.; Šengaut, J.; Bernotienė, R. Different Paths—The Same Virulence: Experimental Study on Avian Single and Co-Infections with Plasmodium Relictum and Plasmodium Elongatum. Int. J. Parasitol. 2018, 48, 1089–1096. [Google Scholar] [CrossRef] [PubMed]
- Harl, J.; Himmel, T.; Valkiūnas, G.; Ilgūnas, M.; Bakonyi, T.; Weissenböck, H. Geographic and Host Distribution of Haemosporidian Parasite Lineages from Birds of the Family Turdidae. Malar. J. 2020, 19, 335. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.K.; Maney, D.L.; Maerz, J.C. The Use of Leukocyte Profiles to Measure Stress in Vertebrates: A Review for Ecologists. Funct. Ecol. 2008, 22, 760–772. [Google Scholar] [CrossRef]
- Romano, M.C.; Jiménez, P.; Miranda-Brito, C.; Valdez, R.A. Parasites and Steroid Hormones: Corticosteroid and Sex Steroid Synthesis, Their Role in the Parasite Physiology and Development. Front. Neurosci. 2015, 9, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appleby, B.M.; Anwar, M.A.; Petty, S.J. Short-term and Long-term Effects of Food Supply on Parasite Burdens in Tawny Owls, Strix Aluco. Funct. Ecol. 1999, 13, 315–321. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Duc, M.; Iezhova, T.A. Description of Haemoproteus Asymmetricus n. Sp. (Haemoproteidae), with Remarks on Predictability of the DNA Haplotype Networks in Haemosporidian Parasite Taxonomy Research. Acta Trop. 2021, 218, 105905. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.F.; Valkiūnas, G.; Nery, C.V.C.; Henrique, P.C.; Gonzalez, I.H.L.; Monteiro, E.F.; de Oliveira Guimarães, L.; Romano, C.M.; Kirchgatter, K. Plasmodium (Novyella) Nucleophilum from an Egyptian Goose in São Paulo Zoo, Brazil: Microscopic Confirmation and Molecular Characterization. Int. J. Parasitol. Parasites Wildl. 2013, 2, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Addler, P.H.; McCreadie, J.W. Black Flies (Simuliidae). In Medical and Veterinary Entomology; Elsevier: San Diego, CA, USA, 2019; ISBN 978-0-12-814043-7. [Google Scholar]
- Svobodová, M.; Dolnik, O.V.; Čepička, I.; Rádrová, J. Biting Midges (Ceratopogonidae) as Vectors of Avian Trypanosomes. Parasit. Vectors 2017, 10, 224. [Google Scholar] [CrossRef] [PubMed]
- Votýpka, J.; Svobodová, M. Trypanosoma Avium: Experimental Transmission from Black Flies to Canaries. Parasitol. Res. 2004, 92, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Mullen, G.R.; Murphree, C.S. Biting Midges (Ceratopogonidae). In Medical and Veterinary Entomology; Elsevier: San Diego, CA, USA, 2019; pp. 213–236. ISBN 978-0-12-814043-7. [Google Scholar]
- Cornford, E.M.; Freeman, B.J.; MacInnis, A.J. Physiological Relationships and Circadian Periodicities in Rodent Trypanosomes. Trans. R. Soc. Trop. Med. Hyg. 1976, 70, 238–243. [Google Scholar] [CrossRef]
- Hawking, F. Circadian Rhythms of Trypanosoma Congolense in Laboratory Rodents. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 592–595. [Google Scholar] [CrossRef]
- Zídková, L.; Cepicka, I.; Szabová, J.; Svobodová, M. Biodiversity of Avian Trypanosomes. Infect. Genet. Evol. 2012, 12, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Hawking, F. The 24-Hour Periodicity of Microfilariae: Biological Mechanisms Responsible for Its Production and Control. Proc. R. Soc. Lond. B Biol. Sci. 1967, 169, 59–76. [Google Scholar] [CrossRef]
- Kelly-Hope, L.; Paulo, R.; Thomas, B.; Brito, M.; Unnasch, T.R.; Molyneux, D. Loa Loa Vectors Chrysops Spp.: Perspectives on Research, Distribution, Bionomics, and Implications for Elimination of Lymphatic Filariasis and Onchocerciasis. Parasit. Vectors 2017, 10, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bain, O. Recherches Sur La Morphogénèse Des Filaires Chez l’hôte Intermédiaire. Ann. Parasitol. Hum. Comparée 1972, 47, 251–303. [Google Scholar] [CrossRef]
- Campbell, J.R.; Soekartono; Purnomo; Atmosoedjono, S.; Marwoto, H. Experimental Wuchereria Kalimantani Infection in the Leaf Monkey, Presbytis Cristata. Ann. Trop. Med. Parasitol. 1986, 80, 141–142. [Google Scholar] [CrossRef]
- Hibler, C.P. Onchocercidae (Nematoda: Filarioidea) of the American Magpie, Pica Pica Hudsonia (Sabine), in Northern Colorado; Colorado State University: Fort Collins, CO, USA, 1964. [Google Scholar]
- Kobayasi, H. On the development of Microfilaria bancrofli in the body of the mosquito (Culex fatigans). Acta Jpn. Med. Trop. 1940, 2, 63–88. [Google Scholar]
- Manguin, S.; Bangs, M.J.; Pothikasikorn, J.; Chareonviriyaphap, T. Review on Global Co-Transmission of Human Plasmodium Species and Wuchereria Bancrofti by Anopheles Mosquitoes. Infect. Genet. Evol. 2010, 10, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Webber, W.A.F. Dirofilaria Aethiops Webber, 1955, a Filarial Parasite of Monkeys III. The Larval Development in Mosquitoes. Parasitology 1955, 45, 388–400. [Google Scholar] [CrossRef] [PubMed]
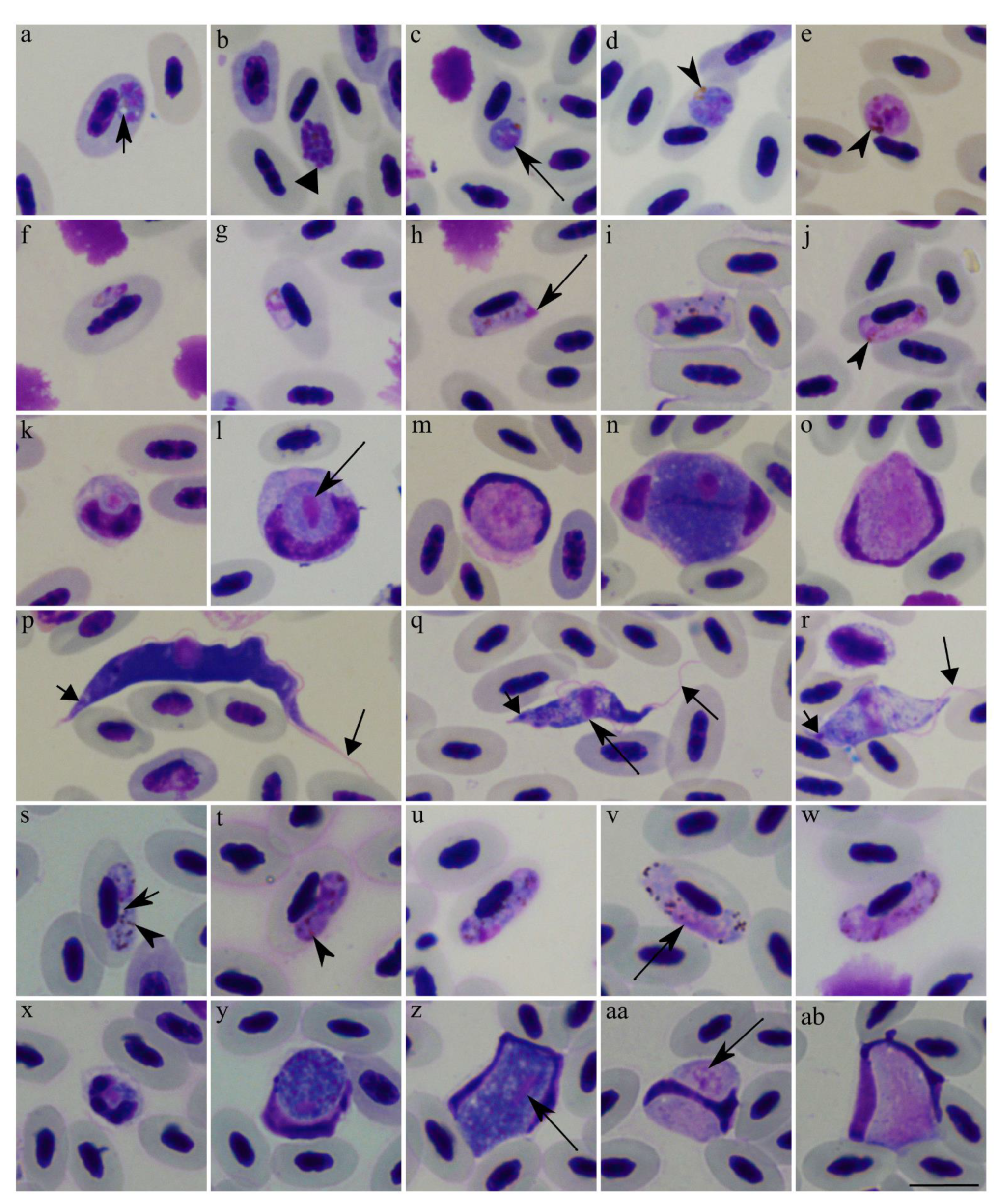
 ), vacuole. Arrowhead (
), vacuole. Arrowhead (  ), pigment granules. Long arrow (
), pigment granules. Long arrow (  ), parasite nucleus. Short triangle arrow (
), parasite nucleus. Short triangle arrow (  ), kinetoplast. Long triangle arrow (
), kinetoplast. Long triangle arrow (  ), flagellum. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), vacuole. Arrowhead ( ), pigment granules. Long arrow ( ), parasite nucleus. Short triangle arrow ( ), kinetoplast. Long triangle arrow ( ), flagellum. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), flagellum. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), vacuole. Arrowhead ( ), pigment granules. Long arrow ( ), parasite nucleus. Short triangle arrow ( ), kinetoplast. Long triangle arrow ( ), flagellum. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films. ), vacuole-like structures. Short triangle arrow ( ), host cell nuclei. Simple wide arrow (
), vacuole-like structures. Short triangle arrow ( ), host cell nuclei. Simple wide arrow (  ), host cell cytoplasm. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), vacuole-like structures. Short triangle arrow ( ), host cell nuclei. Simple wide arrow ( ), host cell cytoplasm. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), host cell cytoplasm. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.
), vacuole-like structures. Short triangle arrow ( ), host cell nuclei. Simple wide arrow ( ), host cell cytoplasm. Scale bar: 10 µm. Methanol fixed and Giemsa-stained blood films.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasite Genus | Parasitemia Intensity Determination | PCR-Protocol | Gene Amplified |
|---|---|---|---|
| Plasmodium | Infected cells on 20,000 erythrocytes [24] | [25] | Cytochrome b (cytb) |
| Haemoproteus | Infected cells on 20,000 erythrocytes [24] | [25] | cytb |
| Leucocytozoon | Infected cells on 20,000 erythrocytes [24] | [25] | cytb |
| Lankesterella | Infected cells on 100 mononuclear leukocytes [26] | [26] | 18S ribosomal RNA (18S) |
| Trypanosoma | Trypomastigotes seen in 1 cm2 of blood film | [27] | 18S |
| Splendidofilaria | Microfilariae seen 1 µL of blood | [4] | 28S nuclear (28S) and cytochrome oxidase I (coxI) |
| Feature | Measurements (n = 21) a |
|---|---|
| Sporozoite | |
| Length | 8.2–9.9 (9.1 ± 0.4) b |
| Width | 2.6–4.0 (3.4 ± 0.4) |
| Area | 21.9–29.3 (25.7 ± 2.2) |
| Sporozoite nucleus | |
| Length | 1.0–2.5 (2.0 ± 0.4) |
| Width | 1.5–3.6 (2.7 ± 0.6) |
| Area | 2.7–5.4 (3.9 ± 0.7) |
| Sporozoite vacuole-like structure | |
| Length | 1.0–2.0 (1.5 ± 0.2) |
| Width | 1.4–2.9 (2.2 ± 0.4) |
| Area | 1.2–3.8 (2.8 ± 0.7) |
| Turdus merula | Turdus philomelos | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Parameter | Blood (n = 25) | Lungs (n = 20) | Uterus (n = 7) | Blood (n = 10) | Lungs (n = 17) | |||||
| MC | % | MC | % | MC | % | MC | % | MC | % | |
| Length | 83–120 | - | 86–143 | - | 75–95 | - | 96–124 | - | 73–113 | - |
| Maximum width | 4–6 | - | 5–6 | - | 3–5 | - | 4–6 | - | 4–6 | - |
| Inner body length | 6–14 | - | 5–15 | - | 2–5 | - | 6–12 | - | 4–13 | - |
| Cephalic space | 1–4 | - | 1–7 | - | 2–6 | - | 1–3 | - | 1–7 | - |
| Distance from AE to: | ||||||||||
| nerve ring | 16–26 | 18–24 | 17–34 | 19–28 | 18–25 | 22–26 | 18–27 | 17–24 | 16–27 | 18–25 |
| excretory pore | 24–47 | 28–35 | 26–49 | 30–35 | 24–31 | 30–36 | 31–40 | 29–35 | 22–39 | 26–36 |
| inner body anterior end | 44–70 | 47–56 | 44–81 | 46–57 | 41–53 | 52–56 | 53–69 | 51–56 | 39–62 | 48–55 |
| inner body posterior end | 50–85 | 57–65 | 51–90 | 55–65 | 44–58 | 55–61 | 60–79 | 58–64 | 43–73 | 58–67 |
| anal pore | 67–108 | 81–87 | 70–119 | 80–85 | 60–78 | 79–82 | 82–105 | 83–87 | 59–96 | 80–88 |
| Feature | Turdus merula | Turdus philomelos | ||
|---|---|---|---|---|
| Male (n = 1) | Female (n = 4) | Male (n = 1) | Female (n = 1) | |
| Body length | 8 | 11–17 | 8 | 15 |
| Maximum width | 345 | 320–403 | 253 | 380 |
| Oesophagus length | 890 | 1010–1026 | 729 | 791 |
| Vagina length | - | 108–128 | - | 82 |
| Left spicule length | 116 | - | 103 | - |
| Right spicule length | 82 | - | 91 | - |
| Nerve ring from AE | 118 | 128–182 | 118 | 126 |
| Vulva from AE | - | 466–646 | - | 795 |
| Testes from AE | 547 | - | 546 | - |
| Ovary from PE | - | 86–239 | - | 447 |
| Cloaca/Anus from PE | 53 | 73–110 | 70 | 153 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chagas, C.R.F.; Binkienė, R.; Valkiūnas, G. Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections. Animals 2021, 11, 3490. https://doi.org/10.3390/ani11123490
Chagas CRF, Binkienė R, Valkiūnas G. Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections. Animals. 2021; 11(12):3490. https://doi.org/10.3390/ani11123490
Chicago/Turabian StyleChagas, Carolina Romeiro Fernandes, Rasa Binkienė, and Gediminas Valkiūnas. 2021. "Description and Molecular Characterization of Two Species of Avian Blood Parasites, with Remarks on Circadian Rhythms of Avian Haematozoa Infections" Animals 11, no. 12: 3490. https://doi.org/10.3390/ani11123490