
First Report of Haemoproteus (Haemosporida, Haemoproteidae) Megalomeronts in the Brain of an Avian Host, with Description of Megalomerogony of Haemoproteus Pastoris, the Blood Parasite of the Common Starling


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Samples
2.2. Histological Examination
2.3. DNA Extraction, PCR and Sequencing
2.4. Phylogenetic Analysis
3. Results
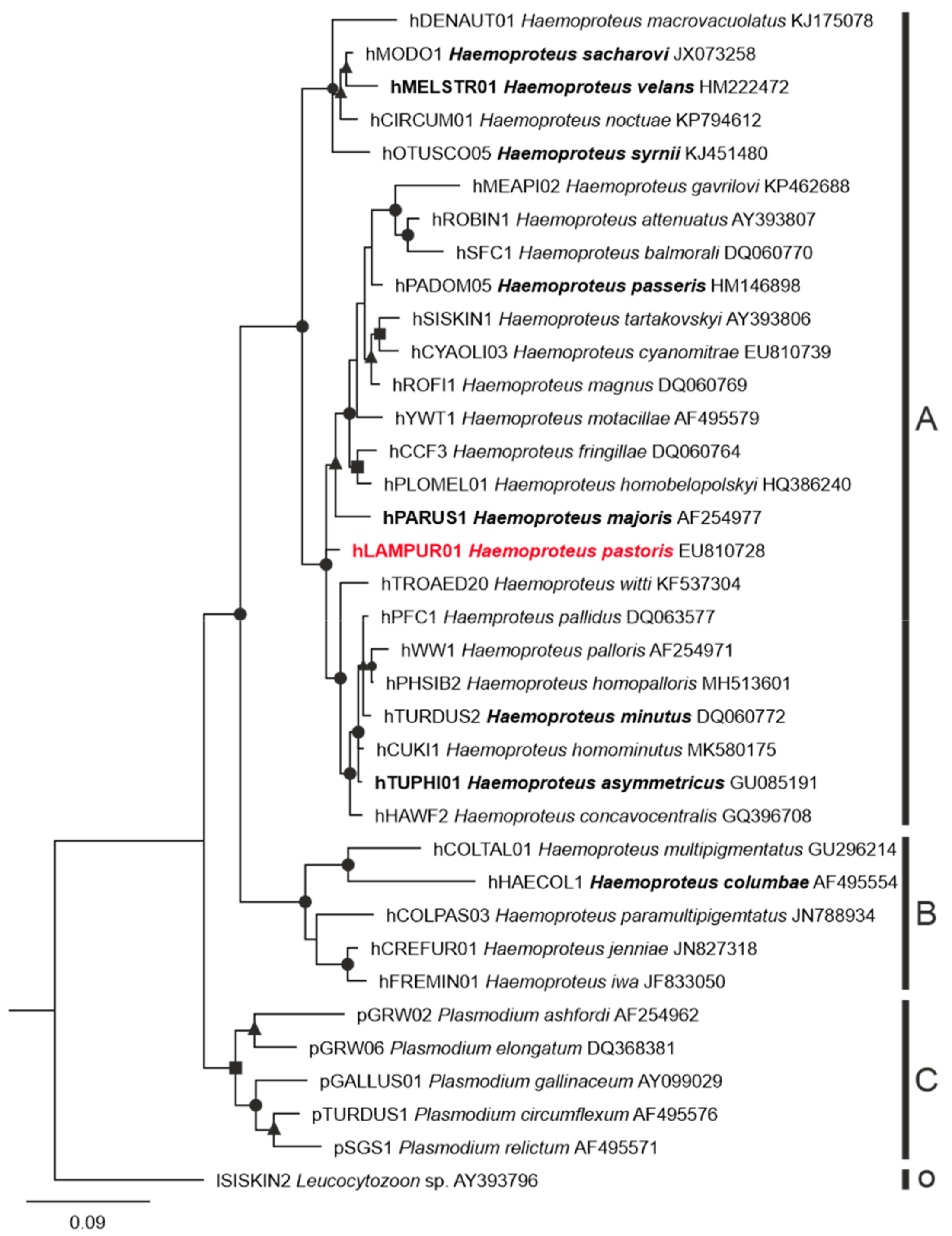
3.1. Parasite Identification and Phylogenetic Relationships
3.2. Description of Megalomeronts
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC Press: Boca Raton, FL, USA, 2005; ISBN 0415300975. [Google Scholar]
- Valkiūnas, G.; Atkinson, C.T. Introduction to life cycles, taxonomy, distribution, and basic research techniques. In Avian Malaria and Related Parasites in the Tropics: Ecology, Evolution and Systematics; Santiago-Alarcon, D., Marzal, A., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 45–80. [Google Scholar]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef]
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Quillfeldt, P.; Arriero, E.; Martínez, J.; Masello, J.F.; Merino, S. Prevalence of blood parasites in seabirds—A review. Front. Zool. 2011, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Shirihai, H.; Svensson, L. Handbook of Western Palearctic birds, Volume II: Passerines: Flycatchers to Bundings; HELM; Bloomsbury Publishing: London, UK, 2018; ISBN 978-1-4729-3757-7. [Google Scholar]
- Chagas, C.R.F.; Bukauskaitė, D.; Ilgūnas, M.; Bernotienė, R.; Iezhova, T.; Valkiūnas, G. Sporogony of four Haemoproteus species (Haemosporida: Haemoproteidae), with report of in vitro ookinetes of Haemoproteus hirundinis: Phylogenetic inference indicates patterns of haemosporidian parasite ookinete development. Parasites Vectors 2019, 12, 422. [Google Scholar] [CrossRef]
- Bukauskaitė, D.; Chagas, C.R.F.; Bernotienė, R.; Žiegytė, R.; Ilgūnas, M.; Iezhova, T.; Valkiūnas, G. A new methodology for sporogony research of avian haemoproteids in laboratory-reared Culicoides spp., with a description of the complete sporogonic development of Haemoproteus pastoris. Parasites Vectors 2019, 12, 582. [Google Scholar] [CrossRef]
- De Beaurepaire de Aragão, H. Über den Entwicklungsgang und die Übertragung von Haemoproteus columbae. Arch. für Protiskenkd. 1908, 12, 154–167. [Google Scholar]
- Atkinson, C.T.; Forrester, D.J.; Greiner, E.C. Pathogenicity of Haemoproteus meleagridis (Haemosporina: Haemoproteidae) in experimentally infected domestic turkeys. J. Parasitol. 1988, 74, 228. [Google Scholar] [CrossRef]
- Miltgen, F.; Landau, I.; Ratanaworabhan, N.; Yenbutra, S. Parahaemoproteus desseri n. sp.; Gamétogonie et schizogonie chez l’hote naturel: Psittacula roseata de Thaïland, et sporogonie expérimentale chez Culicoides nubeculosus. Annu. Parasitol. Hum. Comp. 1981, 56, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Farmer, J.N. Gizzard lesions associated with Haemoproteus sacharovi infections of pigeons. Proc. Iowa Acad. Sci. 1964, 71, 537–542. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A. Exo-erythrocytic development of avian malaria and related haemosporidian parasites. Malar. J. 2017, 16, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, G.F.; Peirce, M.A.; Ashford, R.W. Avian haematozoa: Mortality and pathogenicity. J. Nat. Hist. 1993, 27, 993–1001. [Google Scholar] [CrossRef]
- Cardona, C.J.; Ihejirika, A.; McClellan, L. Haemoproteus lophortyx infection in bobwhite quail. Avian Dis. 2002, 46, 249–255. [Google Scholar] [CrossRef]
- Peirce, M.A.; Lederer, R.; Adlard, R.D.; O’Donoghue, P.J. Pathology associated with endogenous development of haematozoa in birds from southeast Queensland. Avian Pathol. 2004, 33, 445–450. [Google Scholar] [CrossRef]
- Ferrell, S.T.; Snowden, K.; Marlar, A.B.; Garner, M.; Lung, N.P. Fatal hemoprotozoal infections in multiple avian species in a zoological park. J. Zoo Wildl. Med. 2007, 38, 309–316. [Google Scholar] [CrossRef]
- Donovan, T.A.; Schrenzel, M.; Tucker, T.A.; Pessier, A.P.; Stalis, I.H. Hepatic hemorrhage, hemocoelom, and sudden death due to Haemoproteus infection in passerine birds: Eleven cases. J. Vet. Diagn. Investig. 2008, 20, 304–313. [Google Scholar] [CrossRef] [Green Version]
- Olias, P.; Wegelin, M.; Freter, S.; Gruber, A.D.; Klopfleisch, R. Avian malaria deaths in parrots, Europe. Emerg. Infect. Dis. 2011, 17, 950–952. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Catedral, L.; Brunton, D.; Stidworthy, M.F.; Elsheikha, H.M.; Pennycott, T.; Schulze, C.; Braun, M.; Wink, M.; Gerlach, H.; Pendl, H.; et al. Haemoproteus minutus is highly virulent for Australasian and South American parrots. Parasites Vectors 2019, 12, 40. [Google Scholar] [CrossRef] [PubMed]
- Barino, G.T.M.; Rossi, M.F.; de Oliveira, L.; Reis Junior, J.L.; D’Agosto, M.; Dias, R.J.P. Haemoproteus syrnii (Haemosporida: Haemoproteidae) in owls from Brazil: Morphological and molecular characterization, potential cryptic species, and exo-erythrocytic stages. Parasitol. Res. 2021, 120, 243–255. [Google Scholar] [CrossRef]
- Duc, M.; Ilgūnas, M.; Valkiūnas, G. Patterns of Haemoproteus majoris (Haemosporida, Haemoproteidae) megalomeront development. Acta Trop. 2020, 212, 105706. [Google Scholar] [CrossRef]
- Groff, T.C.; Lorenz, T.J.; Crespo, R.; Iezhova, T.; Valkiūnas, G.; Sehgal, R.N.M. Haemoproteosis lethality in a woodpecker, with molecular and morphological characterization of Haemoproteus velans (Haemosporida, Haemoproteidae). Int. J. Parasitol. Parasites Wildl. 2019, 10, 93–100. [Google Scholar] [CrossRef]
- Yoshimoto, M.; Ozawa, K.; Kondo, H.; Echigoya, Y.; Shibuya, H.; Sato, Y.; Sehgal, R.N.M. A fatal case of a captive snowy owl (Bubo scandiacus) with Haemoproteus infection in Japan. Parasitol. Res. 2021, 120, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Himmel, T.; Harl, J.; Kübber-Heiss, A.; Konicek, C.; Fernández, N.; Juan-Sallés, C.; Ilgūnas, M.; Valkiūnas, G.; Weissenböck, H. Molecular probes for the identification of avian Haemoproteus and Leucocytozoon parasites in tissue sections by chromogenic in situ hybridization. Parasites Vectors 2019, 12, 282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilgūnas, M.; Romeiro Fernandes Chagas, C.; Bukauskaitė, D.; Bernotienė, R.; Iezhova, T.; Valkiūnas, G. The life-cycle of the avian haemosporidian parasite Haemoproteus majoris, with emphasis on the exoerythrocytic and sporogonic development. Parasites Vectors 2019, 12, 516. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, C.T.; Greiner, E.C.; Forrester, D.J. Pre-erythrocytic development and associated host responses to Haemoproteus meleagridis (Haemosporina: Haemoproteidae) in experimentally infected domestic turkeys. J. Protozool. 1986, 33, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Godfrey, R.D.; Fedynich, A.M.; Pence, D.B. Quantification of hematozoa in blood smears. J. Wildl. Dis. 1987, 23, 558–565. [Google Scholar] [CrossRef]
- Lillie, R.D. Histopathologic Technic and Practical Histochemistry, 3rd ed.; Blakiston Division, McGraw-Hill: New York, NY, USA, 1965. [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning; A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Bensch, S.; Stjernman, M.; Hasselquist, D.; Ostman, O.; Hansson, B.; Westerdahl, H.; Pinheiro, R.T. Host specificity in avian blood parasites: A study of Plasmodium and Haemoproteus mitochondrial DNA amplified from birds. Proc. R. Soc. B Biol. Sci. 2000, 267, 1583–1589. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Paperna, I.; Gill, H. Schizogonic stages of Haemoproteus from Wenyon’s Baghdad sparrows are also found in Passer domesticus biblicus in Israel. Parasitol. Res. 2003, 91, 486–490. [Google Scholar] [CrossRef]
- Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.A.; Dinhopl, N.; Nedorost, N.; Weissenbacher-Lang, C.; Weissenböck, H.; Valkiūnas, G. Mortality and pathology in birds due to Plasmodium (Giovannolaia) homocircumflexum infection, with emphasis on the exoerythrocytic development of avian malaria parasites. Malar. J. 2016, 15, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desser, S.S.; Fallis, A.M. The cytological development and encapsulation of megaloschizonts of Leucocytozoon simondi. Can. J. Zool. 1967, 45, 1061–1065. [Google Scholar] [CrossRef] [PubMed]
- Huff, C.G. Schizogony and gametocyte development in Leucocytozoon simondi, and comparisons with Plasmodium and Haemoproteus. J. Infect. Dis. 1942, 71, 18–32. [Google Scholar] [CrossRef]
- Ilgūnas, M.; Bukauskaitė, D.; Palinauskas, V.; Iezhova, T.; Fragner, K.; Platonova, E.; Weissenböck, H.; Valkiūnas, G. Patterns of Plasmodium homocircumflexum virulence in experimentally infected passerine birds. Malar. J. 2019, 18, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleischman, R.W.; Squire, R.A.; Sladen, W.J.L.; Moore, J. Pathologic confirmation of malaria (Plasmodium elongatum) in African penguins (Spheniscus demersus). Bull. Wildl. Dis. Assoc. 1968, 4, 133–135. [Google Scholar] [CrossRef] [PubMed]
- Howe, L.; Castro, I.C.; Schoener, E.R.; Hunter, S.; Barraclough, R.K.; Alley, M.R. Malaria parasites (Plasmodium spp.) infecting introduced, native and endemic New Zealand birds. Parasitol. Res. 2012, 110, 913–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desser, S.S. Schizogony and gametogony of Leucocytozoon simondi and associated reactions in the avian host. J. Protozool. 1967, 14, 244–254. [Google Scholar] [CrossRef]
- Karstad, L. A case of leucocytozoonosis in a wild. Wildl. Dis. Assoc. 1965, 1, 33–34. [Google Scholar]
- Valkiūnas, G.; Ilgūnas, M.; Bukauskaitė, D.; Duc, M.; Iezhova, T.A. Description of Haemoproteus asymmetricus n. sp. (Haemoproteidae), with remarks on predictability of the DNA haplotype networks in haemosporidian parasite taxonomy research. Acta Trop. 2021, 218, 105905. [Google Scholar] [CrossRef]
- Valkiūnas, G.; Žičkus, T.; Shapoval, A.P.; Iezhova, T.A. Effect of Haemoproteus belopolskyi (Haemosporida: Haemoproteidae) on body mass of the blackcap Sylvia atricapilla. J. Parasitol. 2006, 92, 1123–1125. [Google Scholar] [CrossRef] [PubMed]
- Garnham, P.C.C. Malaria Parasites and Other Haemosporidia; Blackwell Scientific Publications: Oxford, UK, 1966. [Google Scholar]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. Parasitic Diseases of Wild Birds; Wiley-Blackwell: Oxford, UK, 2008; ISBN 9780813820811. [Google Scholar]
- Valkiūnas, G.; Iezhova, T.A.; Loiseau, C.; Sehgal, R.N.M. Nested cytochrome b Polymerase Chain Reaction diagnostics detect sporozoites of Hemosporidian parasites in peripheral blood of naturally infected birds. J. Parasitol. 2009, 95, 1512–1515. [Google Scholar] [CrossRef] [PubMed]
- Morii, T. A review of Leucocytozoon caulleryi infection in chickens. J. Protozool. Res. 1992, 2, 128–133. [Google Scholar]
- Nilsson, E.; Taubert, H.; Hellgren, O.; Huang, X.; Palinauskas, V.; Markovets, M.Y.; Valkiūnas, G.; Bensch, S. Multiple cryptic species of sympatric generalists within the avian blood parasite Haemoproteus majoris. J. Evol. Biol. 2016, 29, 1812–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
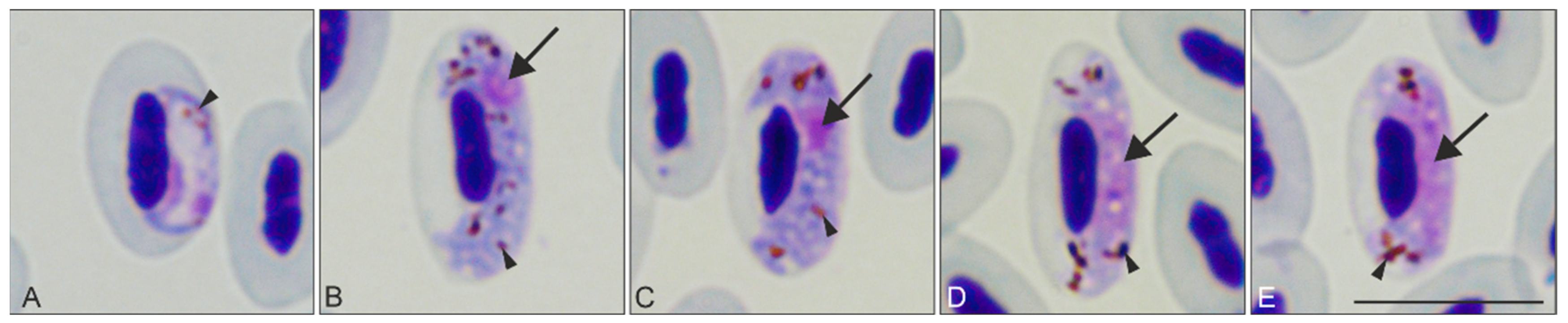
 )—parasite nucleus. Triangle arrowhead (
)—parasite nucleus. Triangle arrowhead (  )—pigment granules. Images were taken using Giemsa-stained blood films at ×1000 magnification. Scale bar = 10 μm.
)—parasite nucleus. Triangle arrowhead ( )—pigment granules. Images were taken using Giemsa-stained blood films at ×1000 magnification. Scale bar = 10 μm.
)—pigment granules. Images were taken using Giemsa-stained blood films at ×1000 magnification. Scale bar = 10 μm.
)—parasite nucleus. Triangle arrowhead ( )—pigment granules. Images were taken using Giemsa-stained blood films at ×1000 magnification. Scale bar = 10 μm.

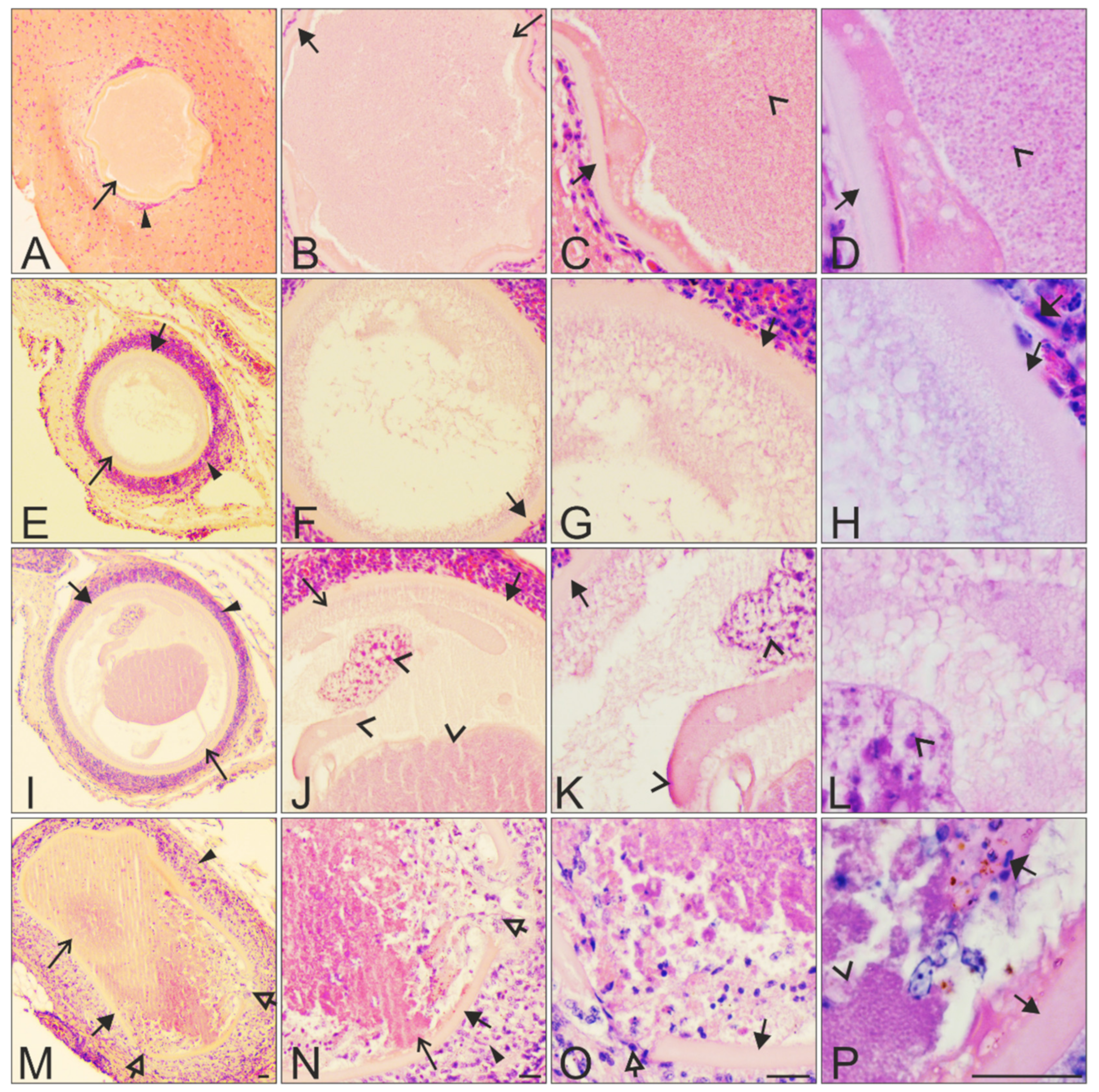
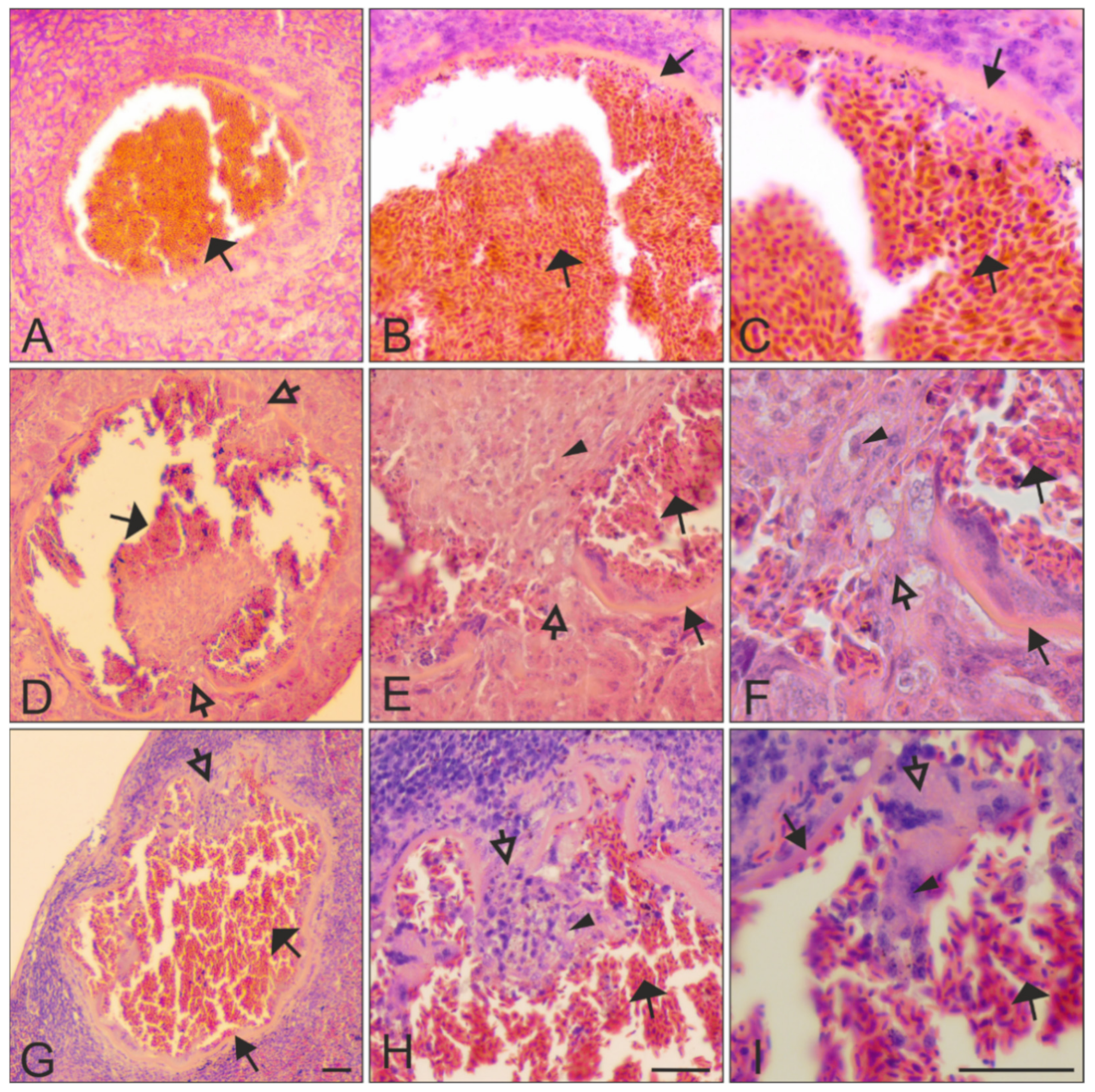
 )—megalomeronts. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows (
)—megalomeronts. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows (  )—rupture of the capsular-like wall. Flat triangle arrows (
)—rupture of the capsular-like wall. Flat triangle arrows (  )—red blood cells inside or outside the megalomeront. Triangle arrowheads ( )—deformed adjacent tissue cells suppressed by the megalomeront. Simple arrowheads (
)—red blood cells inside or outside the megalomeront. Triangle arrowheads ( )—deformed adjacent tissue cells suppressed by the megalomeront. Simple arrowheads (  )—merozoites. All scale bars = 20 μm.
)—megalomeronts. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells inside or outside the megalomeront. Triangle arrowheads ( )—deformed adjacent tissue cells suppressed by the megalomeront. Simple arrowheads ( )—merozoites. All scale bars = 20 μm.
)—merozoites. All scale bars = 20 μm.
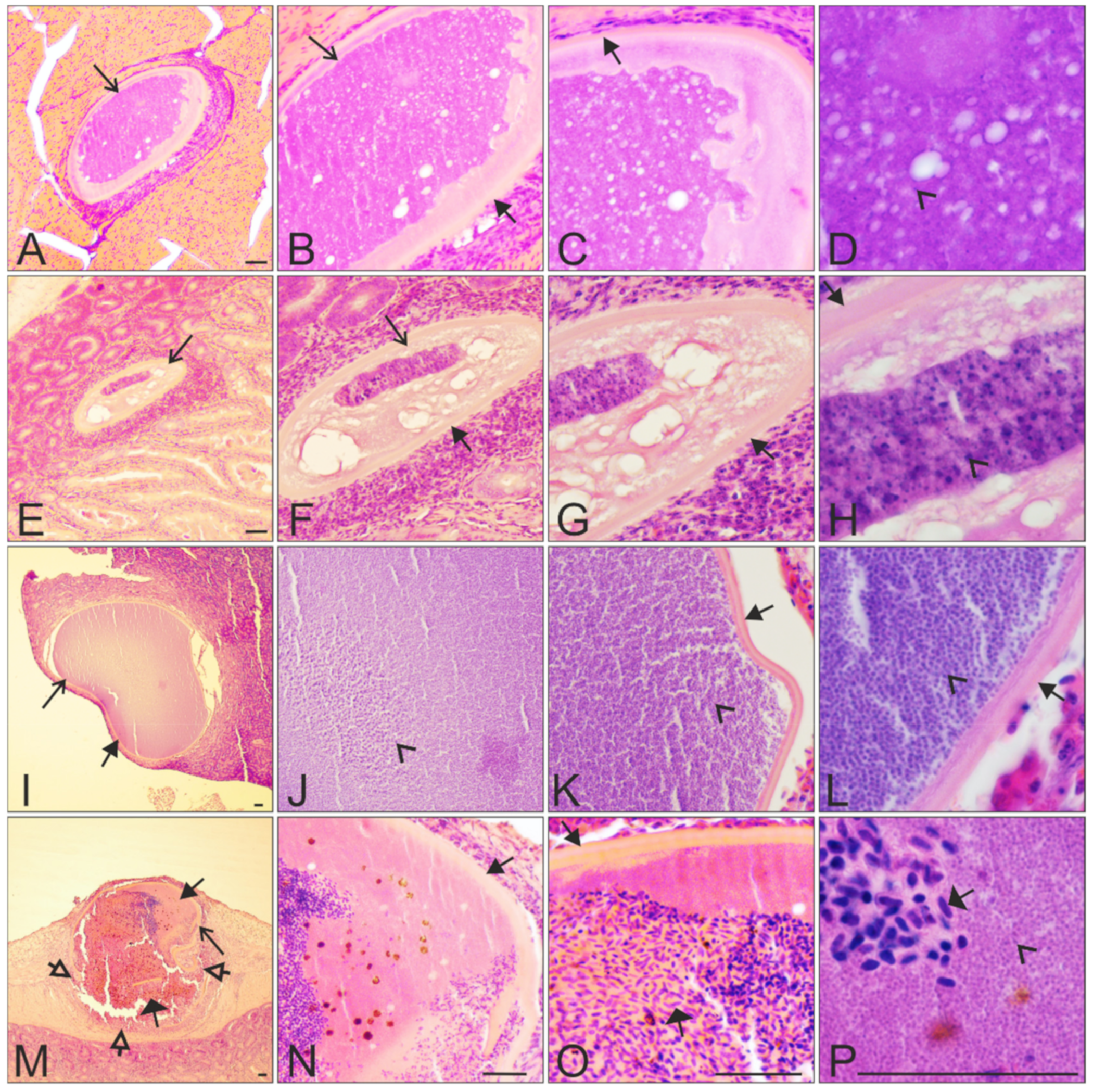
)—megalomeronts. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells inside or outside the megalomeront. Triangle arrowheads ( )—deformed adjacent tissue cells suppressed by the megalomeront. Simple arrowheads ( )—merozoites. All scale bars = 20 μm. )—megalomeront. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Simple arrowheads ( )—merozoite. All scale bars = 40 μm.
)—megalomeront. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Simple arrowheads ( )—merozoite. All scale bars = 40 μm.
)—megalomeront. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Simple arrowheads ( )—merozoite. All scale bars = 40 μm.
)—megalomeront. Filled-black triangle arrows ( )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Simple arrowheads ( )—merozoite. All scale bars = 40 μm. )—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Triangle arrowheads ( )—other cells invading the megalomeront. All scale bars = 40 μm.
)—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Triangle arrowheads ( )—other cells invading the megalomeront. All scale bars = 40 μm.
)—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Triangle arrowheads ( )—other cells invading the megalomeront. All scale bars = 40 μm.
)—capsular-like wall. Contoured-black triangle arrows ( )—rupture of the capsular-like wall. Flat triangle arrows ( )—red blood cells invading the megalomeront. Triangle arrowheads ( )—other cells invading the megalomeront. All scale bars = 40 μm.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual Number | Parasitemia (%) | Brain | Intestine | Pancreas | Kidneys | Lungs | Esophagus | Spleen | Gizzard | Trachea |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 1 | 0 a | growing (2) | 0 | growing (1) | 0 | ruptured (1) | growing (1) ruptured (3) | 0 | 0 |
| 2 | 2 | 0 | growing (1) | 0 | 0 | 0 | 0 | growing (1) | 0 | growing (1) |
| 3 | 3 | 0 | growing (4) ruptured (4) | growing (1) mature (1) ruptured (1) | 0 | growing (1) | 0 | 0 | growing (1) | 0 |
| 4 | 10 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| 5 | 26 | mature (1) | ruptured (2) | 0 | 0 | 0 | growing (1) ruptured (1) | 0 | growing (2) | 0 |
| Parasite Species (Lineage) | Recorded | Bird Species (Family) | Country (Continent) |
|---|---|---|---|
| H. pastoris (hLAMPUR01) | 1 | Bulgaria (Europe) | |
| 1 | Sturnus vulgaris (Sturnidae) | Turkey (Asia) | |
| 1 | Iran (Asia) | ||
| H. pastoris (hLAMPUR01) | 1 | Sturnus roseus (Sturnidae) | Bulgaria (Europe) |
| H. pastoris (hLAMPUR01) | 1 | Lamprotonis pupureiceps (Sturnidae) | Gabon (Africa) |
| H. sp. (hLAMPUR02) a | 1 | Lamprotonis pupureiceps (Sturnidae) | Gabon (Africa) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duc, M.; Ilgūnas, M.; Kubiliūnaitė, M.; Valkiūnas, G. First Report of Haemoproteus (Haemosporida, Haemoproteidae) Megalomeronts in the Brain of an Avian Host, with Description of Megalomerogony of Haemoproteus Pastoris, the Blood Parasite of the Common Starling. Animals 2021, 11, 2824. https://doi.org/10.3390/ani11102824
Duc M, Ilgūnas M, Kubiliūnaitė M, Valkiūnas G. First Report of Haemoproteus (Haemosporida, Haemoproteidae) Megalomeronts in the Brain of an Avian Host, with Description of Megalomerogony of Haemoproteus Pastoris, the Blood Parasite of the Common Starling. Animals. 2021; 11(10):2824. https://doi.org/10.3390/ani11102824
Chicago/Turabian StyleDuc, Mélanie, Mikas Ilgūnas, Monika Kubiliūnaitė, and Gediminas Valkiūnas. 2021. "First Report of Haemoproteus (Haemosporida, Haemoproteidae) Megalomeronts in the Brain of an Avian Host, with Description of Megalomerogony of Haemoproteus Pastoris, the Blood Parasite of the Common Starling" Animals 11, no. 10: 2824. https://doi.org/10.3390/ani11102824