UVB Irradiation Induced Cell Damage and Early Onset of Junbb Expression in Zebrafish

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Keeping and Ethics
2.2. Zebrafish UVB Irradiation
2.3. Histology
2.4. Acridine Orange Staining
2.5. ROS Detection
2.6. Whole-Mount Immunostaining
2.7. Quantitative Real-Time-PCR
2.8. Image Acquisition, Quantification, and Statistics
2.9. Microarray Analysis
2.10. Statistical Analysis
3. Results
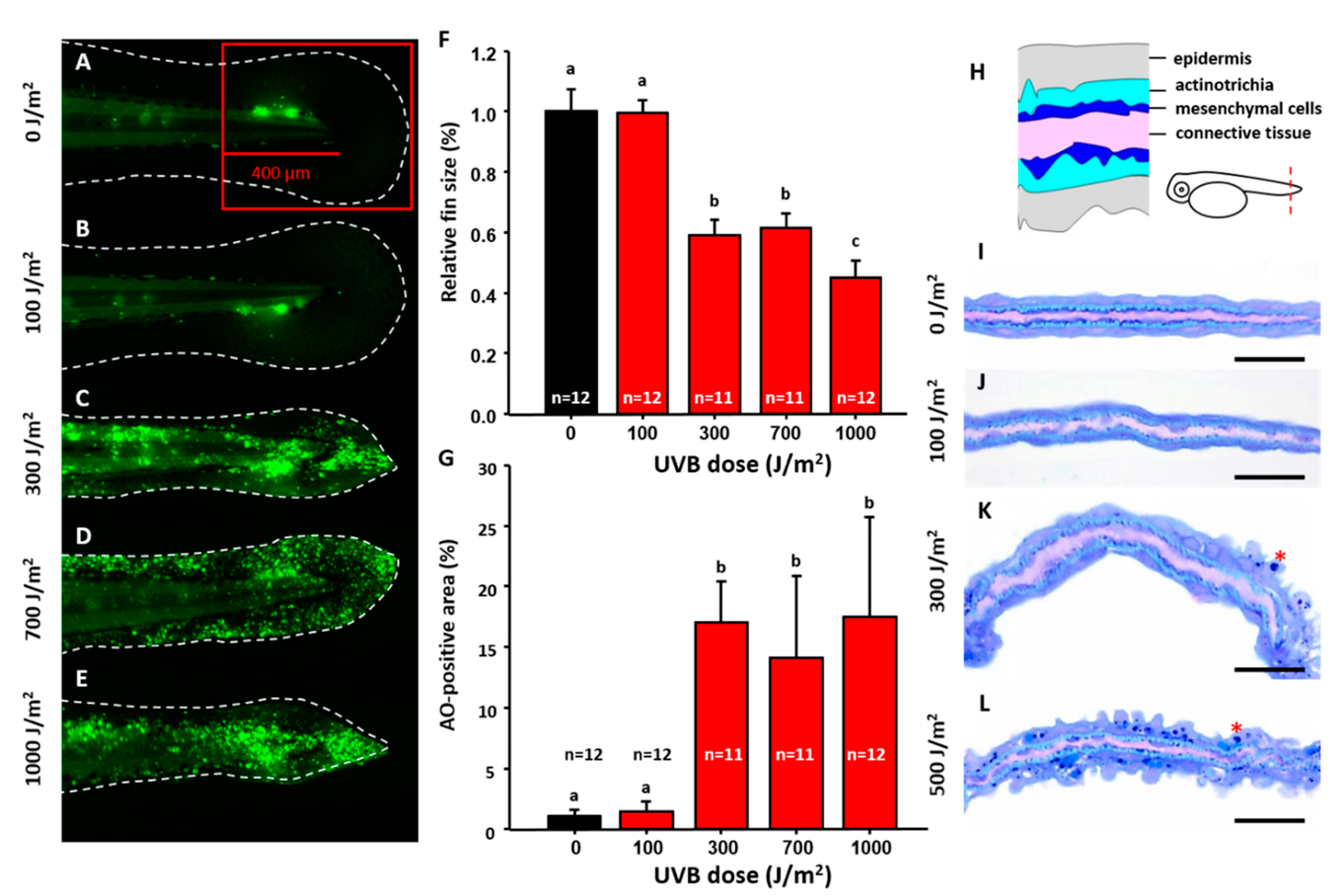
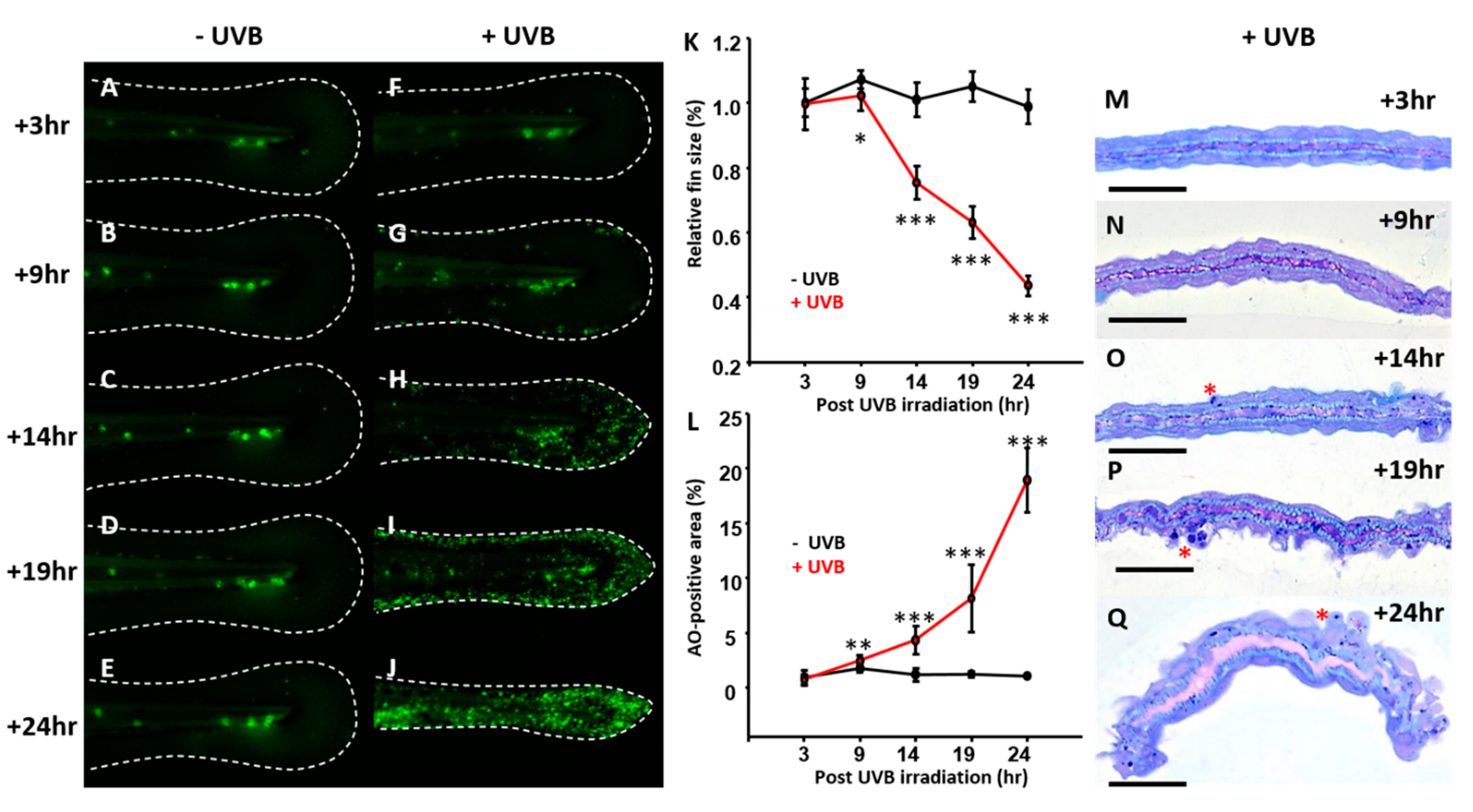
3.1. Optimization of UVB-Induced Zebrafish Model
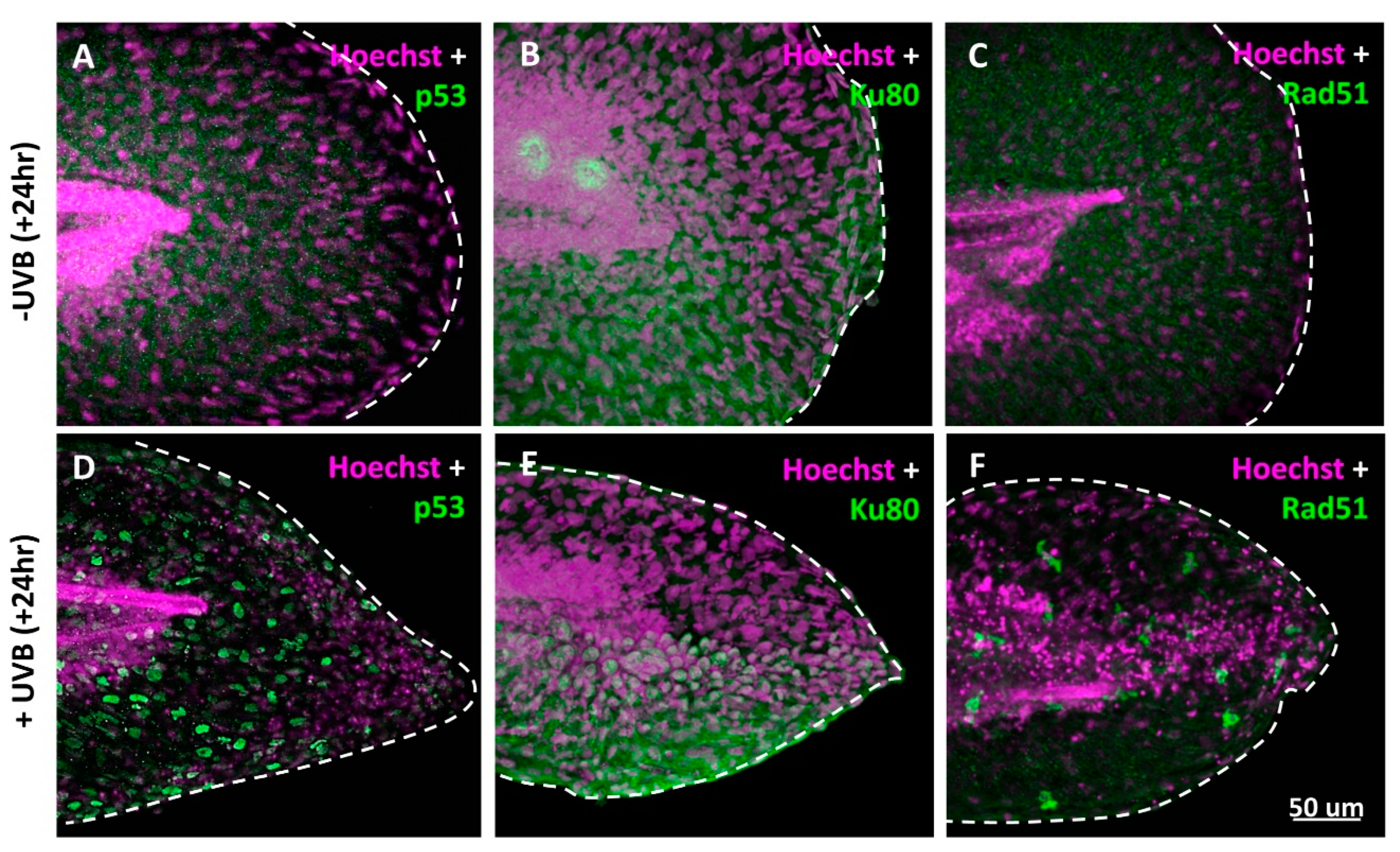
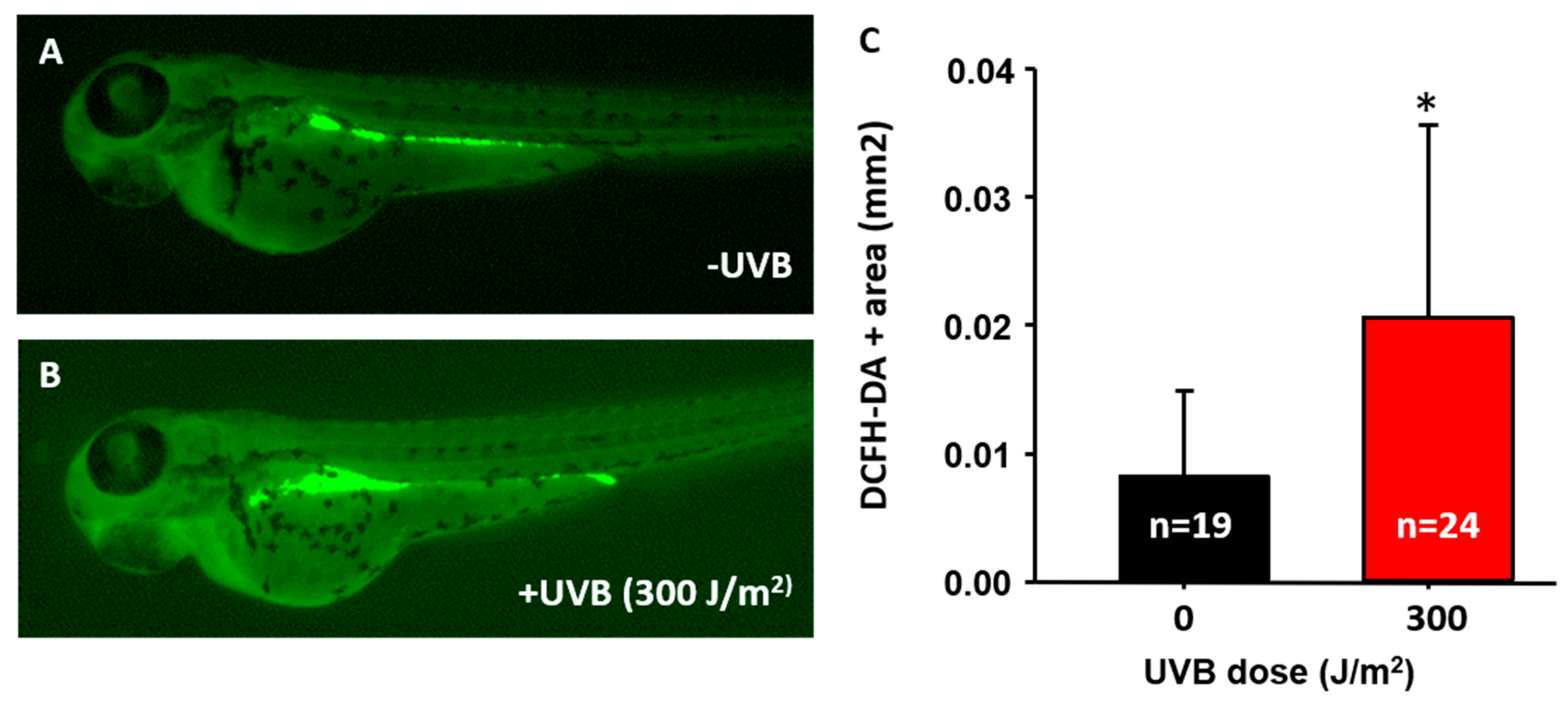
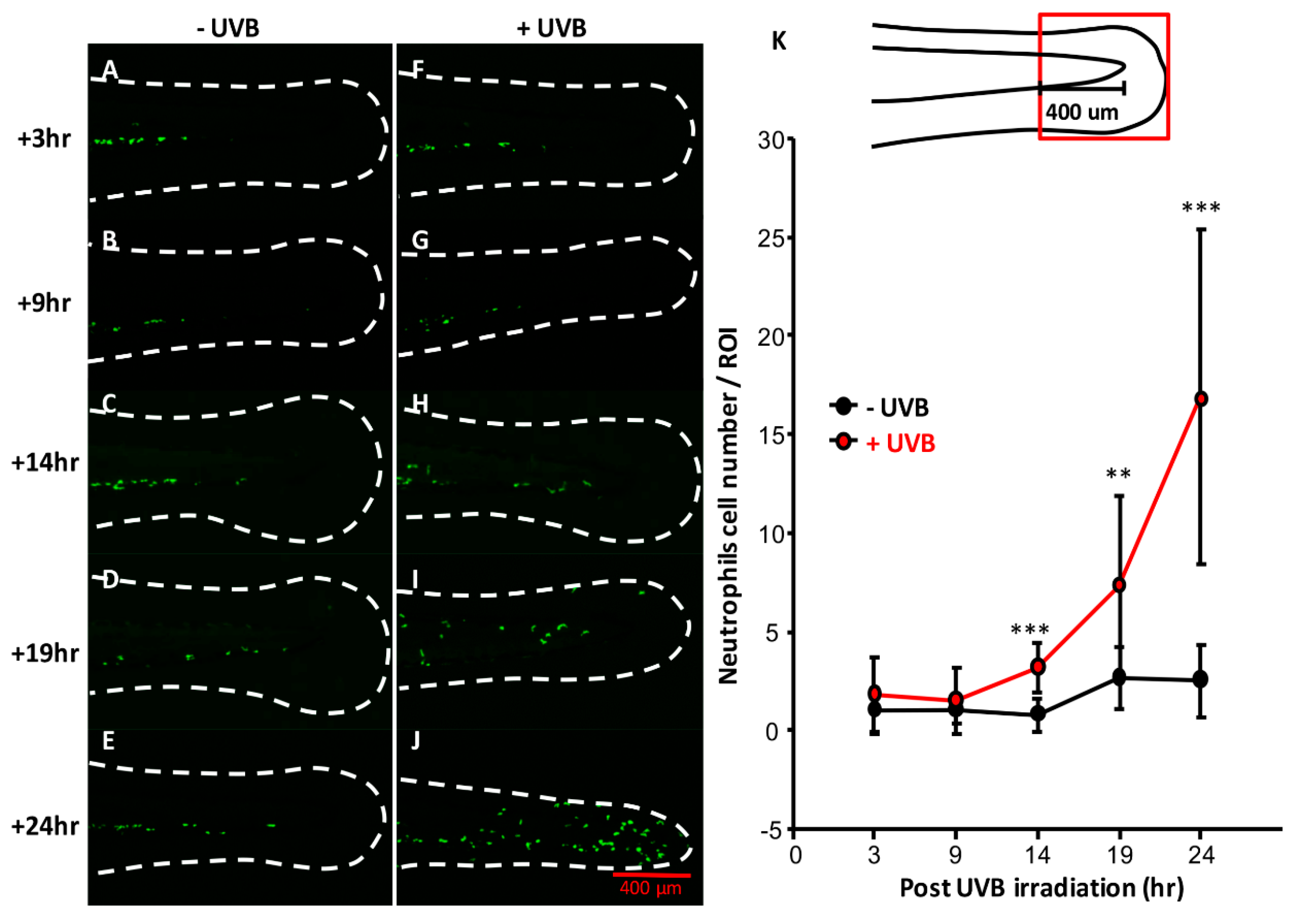
3.2. DNA Damage Induction, ROS Release, and Inflammatory Response after UVB Irradiation
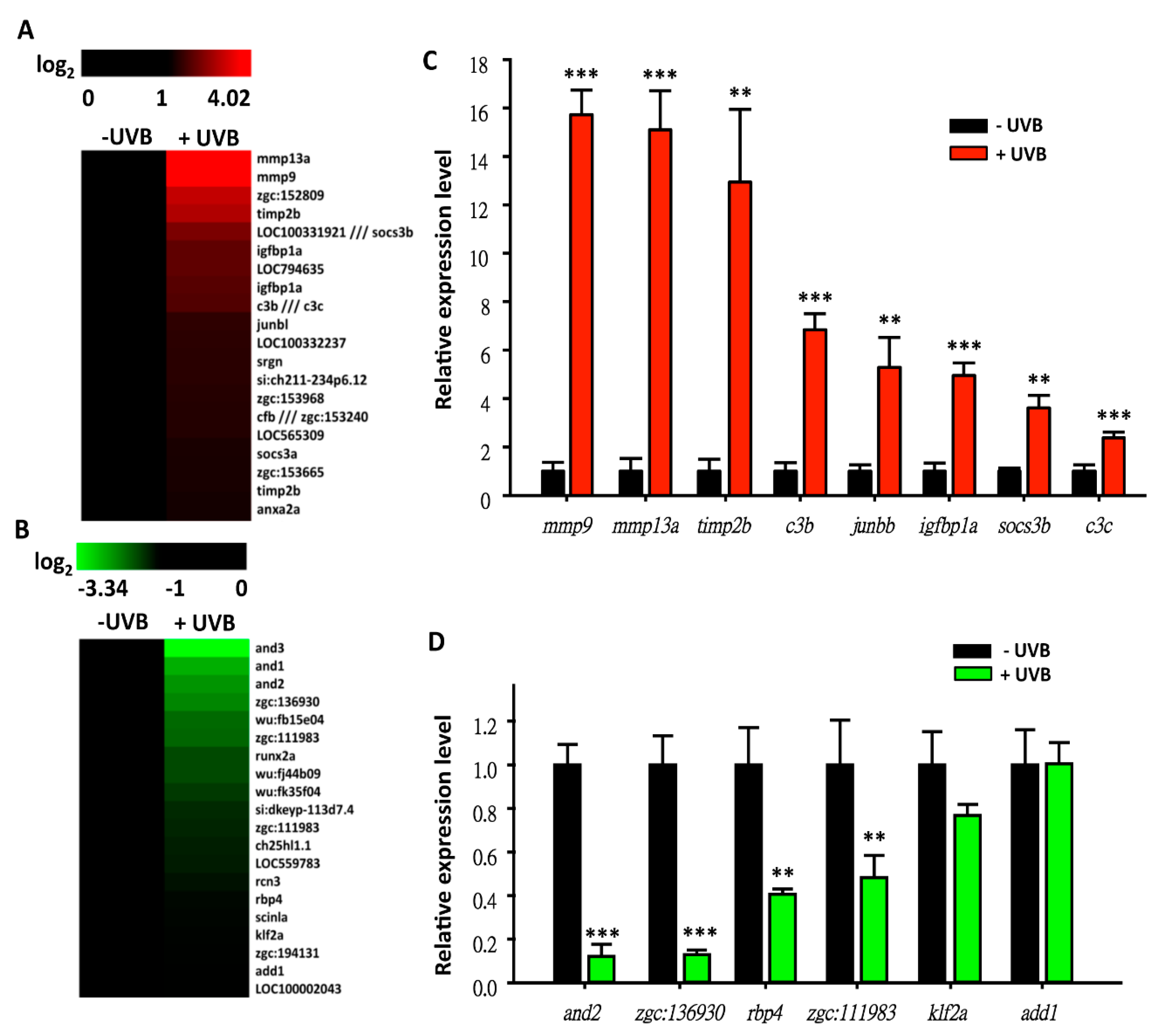
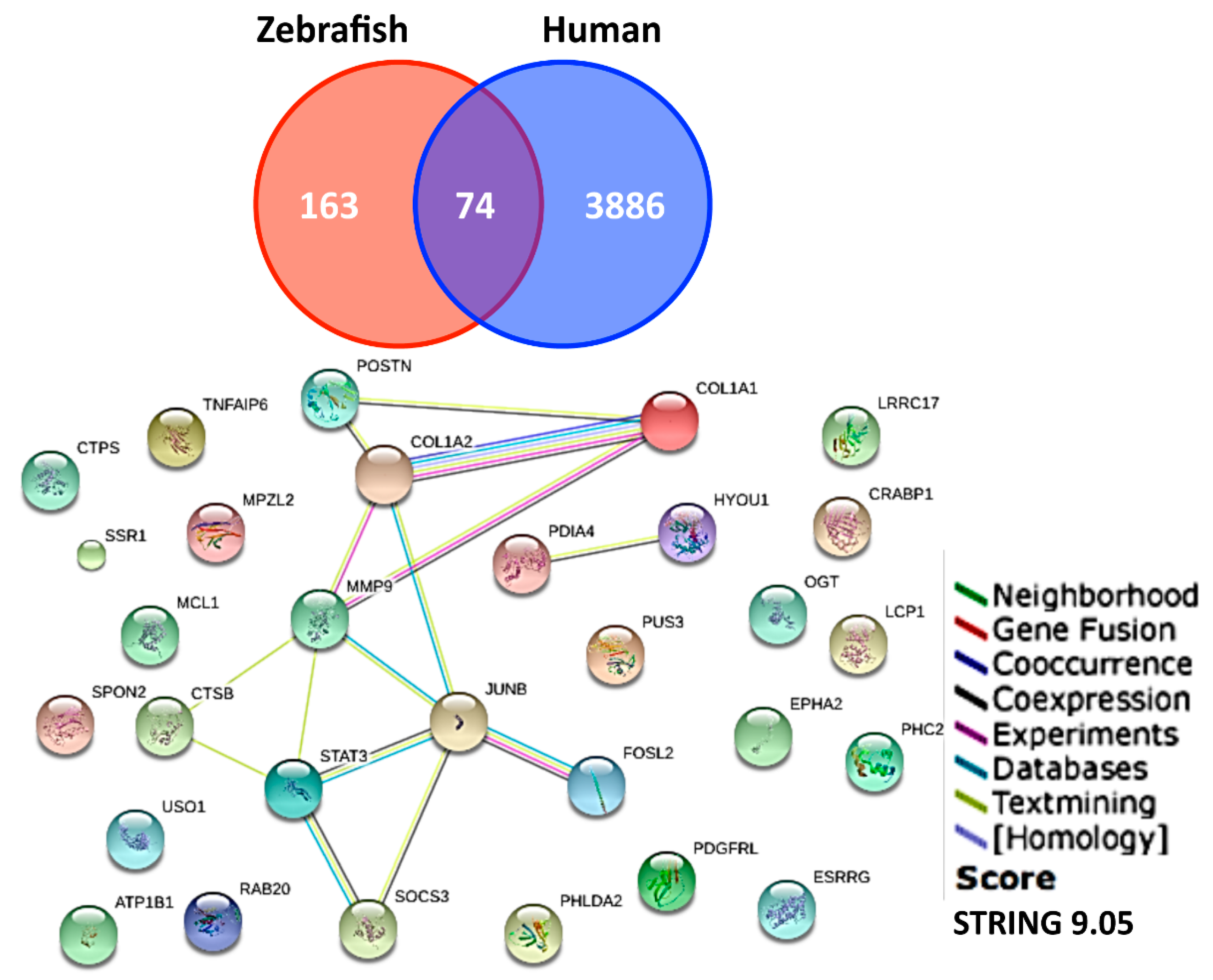
3.3. Screening on Novel UVB-Inducible Markers by Microarray Analysis
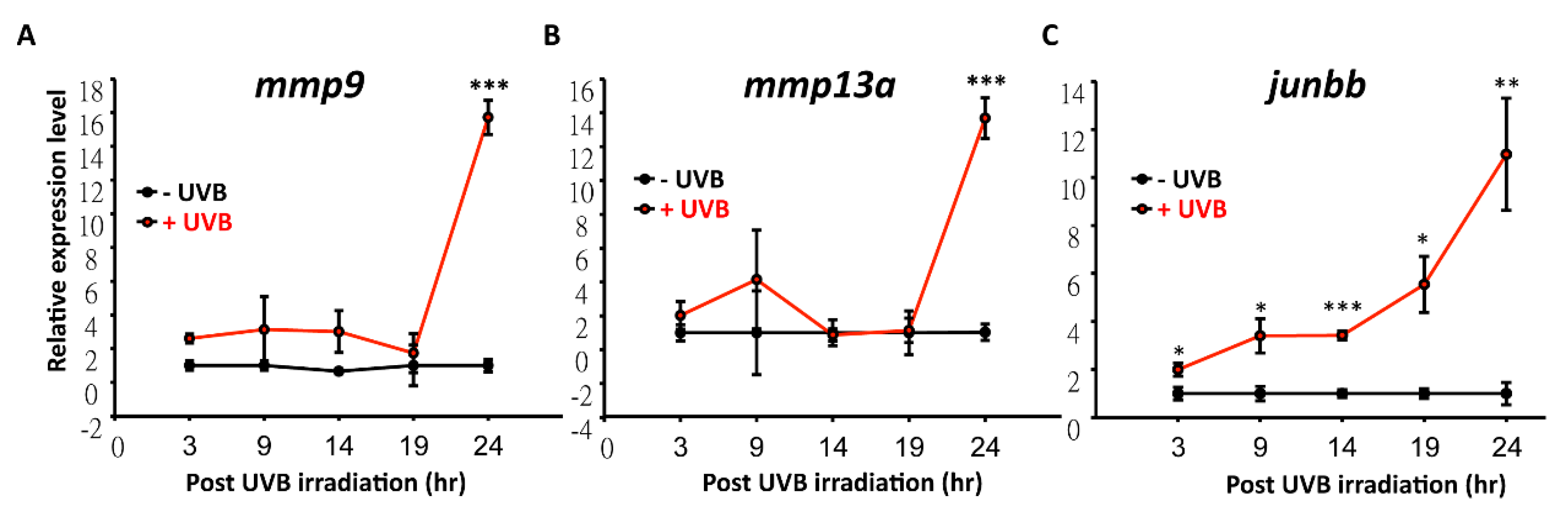
3.4. Temporal and Spatial Expression Patterns of UVB-Inducible Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maverakis, E.; Miyamura, Y.; Bowen, M.P.; Correa, G.; Ono, Y.; Goodarzi, H. Light, including ultraviolet. J. Autoimmun. 2010, 34, J247–J257. [Google Scholar] [CrossRef] [Green Version]
- Diaz, S.; Camilion, C.; Deferrari, G.; Fuenzalida, H.; Armstrong, R.; Booth, C.; Paladini, A.; Cabrera, S.; Casiccia, C.; Lovengreen, C.; et al. Ozone and uv radiation over southern south america: Climatology and anomalies. Photochem. Photobiol. 2006, 82, 834–843. [Google Scholar] [CrossRef]
- El-Nouby, A.M. Effect of stratospheric ozone in uvb solar radiation reaching the earth’s surface at qena, egypt. Atmos. Pollut. Res. 2010, 1, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Holick, M.F. Sunlight, uv-radiation, vitamin d and skin cancer: How much sunlight do we need? Adv. Exp. Med. Biol. 2008, 624, 1–15. [Google Scholar]
- Uchida, Y.; Morimoto, Y.; Uchiike, T.; Kamamoto, T.; Hayashi, T.; Arai, I.; Nishikubo, T.; Takahashi, Y. Phototherapy with blue and green mixed-light is as effective against unconjugated jaundice as blue light and reduces oxidative stress in the gunn rat model. Early Hum. Dev. 2015, 91, 381–385. [Google Scholar] [CrossRef]
- Rajakumar, K.; Greenspan, S.L.; Thomas, S.B.; Holick, M.F. Solar ultraviolet radiation and vitamin d: A historical perspective. Am. J. Public Health 2007, 97, 1746–1754. [Google Scholar] [CrossRef]
- Flament, F.; Bazin, R.; Laquieze, S.; Rubert, V.; Simonpietri, E.; Piot, B. Effect of the sun on visible clinical signs of aging in caucasian skin. Clin. Cosmet. Investig. Dermatol. 2013, 6, 221–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; Little, E.; Quan, H.; Qin, Z.; Voorhees, J.J.; Fisher, G.J. Elevated matrix metalloproteinases and collagen fragmentation in photodamaged human skin: Impact of altered extracellular matrix microenvironment on dermal fibroblast function. J. Investig. Dermatol. 2013, 133, 1362–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skobowiat, C.; Slominski, A.T. Sun-derived infrared a and ultraviolet b radiation: Allies or enemies in melanomagenesis? Exp. Dermatol. 2016, 25, 760–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, F.P.; Zaidi, M.R.; Wolnicka-Glubisz, A.; Anver, M.R.; Bahn, J.; Wielgus, A.; Cadet, J.; Douki, T.; Mouret, S.; Tucker, M.A.; et al. Melanoma induction by ultraviolet a but not ultraviolet b radiation requires melanin pigment. Nat. Commun. 2012, 3, 884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominski, A.T.; Zmijewski, M.A.; Semak, I.; Zbytek, B.; Pisarchik, A.; Li, W.; Zjawiony, J.; Tuckey, R.C. Cytochromes p450 and skin cancer: Role of local endocrine pathways. Anticancer Agents Med. Chem. 2014, 14, 77–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, A.; Johnson, B.E. Dose response, wavelength dependence and rate of excision of ultraviolet radiation-induced pyrimidine dimers in mouse skin DNA. Biochim. Biophys. Acta 1978, 517, 24–30. [Google Scholar] [CrossRef]
- Ikehata, H.; Mori, T.; Kamei, Y.; Douki, T.; Cadet, J.; Yamamoto, M. Wavelength-and tissue-dependent variations in the mutagenicity of cyclobutane pyrimidine dimers in mouse skin. Photochem. Photobiol. 2020, 96, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Sinha, R.P.; Hader, D.P. Uv-induced DNA damage and repair: A review. Photochem. Photobiol. Sci. 2002, 1, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.T.; Zmijewski, M.A.; Skobowiat, C.; Zbytek, B.; Slominski, R.M.; Steketee, J.D. Sensing the environment: Regulation of local and global homeostasis by the skin’s neuroendocrine system. Adv. Anat. Embryol. Cell Biol. 2012, 212, 1–115. [Google Scholar]
- Skobowiat, C.; Slominski, A.T. Uvb activates hypothalamic-pituitary-adrenal axis in c57bl/6 mice. J. Investig. Dermatol. 2015, 135, 1638–1648. [Google Scholar] [CrossRef] [Green Version]
- Grandjean-Laquerriere, A.; Le Naour, R.; Gangloff, S.C.; Guenounou, M. Differential regulation of tnf-alpha, il-6 and il-10 in uvb-irradiated human keratinocytes via cyclic amp/protein kinase a pathway. Cytokine 2003, 23, 138–149. [Google Scholar] [CrossRef]
- Bald, T.; Quast, T.; Landsberg, J.; Rogava, M.; Glodde, N.; Lopez-Ramos, D.; Kohlmeyer, J.; Riesenberg, S.; van den Boorn-Konijnenberg, D.; Homig-Holzel, C.; et al. Ultraviolet-radiation-induced inflammation promotes angiotropism and metastasis in melanoma. Nature 2014, 507, 109–113. [Google Scholar] [CrossRef]
- Banerjee, S.; Leptin, M. Systemic response to ultraviolet radiation involves induction of leukocytic il-1beta and inflammation in zebrafish. J. Immunol. 2014, 193, 1408–1415. [Google Scholar] [CrossRef] [Green Version]
- Kennedy Crispin, M.; Fuentes-Duculan, J.; Gulati, N.; Johnson-Huang, L.M.; Lentini, T.; Sullivan-Whalen, M.; Gilleaudeau, P.; Cueto, I.; Suarez-Farinas, M.; Lowes, M.A.; et al. Gene profiling of narrowband uvb-induced skin injury defines cellular and molecular innate immune responses. J. Investig. Dermatol. 2013, 133, 692–701. [Google Scholar] [CrossRef] [Green Version]
- Skobowiat, C.; Sayre, R.M.; Dowdy, J.C.; Slominski, A.T. Ultraviolet radiation regulates cortisol activity in a waveband-dependent manner in human skin ex vivo. Br. J. Dermatol. 2013, 168, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Enk, C.D.; Jacob-Hirsch, J.; Gal, H.; Verbovetski, I.; Amariglio, N.; Mevorach, D.; Ingber, A.; Givol, D.; Rechavi, G.; Hochberg, M. The uvb-induced gene expression profile of human epidermis in vivo is different from that of cultured keratinocytes. Oncogene 2006, 25, 2601–2614. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Frank, M.; Thisse, C.I.; Thisse, B.V.; Uitto, J. Zebrafish: A model system to study heritable skin diseases. J. Investig. Dermatol. 2011, 131, 565–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sire, J.Y.; Akimenko, M.A. Scale development in fish: A review, with description of sonic hedgehog (shh) expression in the zebrafish (danio rerio). Int. J. Dev. Biol. 2004, 48, 233–247. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Xie, G.; Fan, Q.; Xie, J. Activation of the hedgehog-signaling pathway in human cancer and the clinical implications. Oncogene 2010, 29, 469–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Reilly, K.E.; de Miera, E.V.; Segura, M.F.; Friedman, E.; Poliseno, L.; Han, S.W.; Zhong, J.; Zavadil, J.; Pavlick, A.; Hernando, E.; et al. Hedgehog pathway blockade inhibits melanoma cell growth in vitro and in vivo. Pharmaceuticals 2013, 6, 1429–1450. [Google Scholar] [CrossRef] [PubMed]
- Aksakal, F.I.; Ciltas, A.; Sciences, P. The impact of ultraviolet b (uv-b) radiation in combination with different temperatures in the early life stage of zebrafish (danio rerio). Photochem. Photobiol. Sci. 2018, 17, 35–41. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Wen, C.-C.; Lin, C.-Y.; Chou, C.-Y.; Yang, Z.-S.; Wang, Y.-H. Uv-induced fin damage in zebrafish as a system for evaluating the chemopreventive potential of broccoli and cauliflower extracts. Toxicol. Mech. Methods 2011, 21, 63–69. [Google Scholar] [CrossRef]
- Storer, N.Y.; Zon, L.I. Zebrafish models of p53 functions. Cold Spring Harb. Perspect. Biol. 2010, 2, a001123. [Google Scholar] [CrossRef]
- Vazquez, A.; Bond, E.E.; Levine, A.J.; Bond, G.L. The genetics of the p53 pathway, apoptosis and cancer therapy. Nat. Rev. Drug Discov. 2008, 7, 979–987. [Google Scholar] [CrossRef]
- Tyrrell, R.M. Ultraviolet radiation and free radical damage to skin. Biochem. Soc. Symp. 1995, 61, 47–53. [Google Scholar]
- LeBel, C.P.; Ischiropoulos, H.; Bondy, S.C. Evaluation of the probe 2’,7’-dichlorofluorescin as an indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 1992, 5, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Zhang, Y.; Zhao, F.; Ruan, H.; Huang, H.; Luo, L.; Li, L. Acetylcholine serves as a derepressor in loperamide-induced opioid-induced bowel dysfunction (oibd) in zebrafish. Sci. Rep. 2014, 4, 5602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, J.J.; Cowing-Zitron, C.; Nakatsuji, T.; Muehleisen, B.; Muto, J.; Borkowski, A.W.; Martinez, L.; Greidinger, E.L.; Yu, B.D.; Gallo, R.L. Ultraviolet radiation damages self noncoding rna and is detected by tlr3. Nat. Med. 2012, 18, 1286. [Google Scholar] [CrossRef]
- Renshaw, S.A.; Loynes, C.A.; Trushell, D.M.; Elworthy, S.; Ingham, P.W.; Whyte, M.K. A transgenic zebrafish model of neutrophilic inflammation. Blood 2006, 108, 3976–3978. [Google Scholar] [CrossRef]
- Waster, P.; Orfanidis, K.; Eriksson, I.; Rosdahl, I.; Seifert, O.; Ollinger, K. Uv radiation promotes melanoma dissemination mediated by the sequential reaction axis of cathepsins-tgf-beta1-fap-alpha. Br. J. Cancer 2017, 117, 535–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Pal, A.S.; Hsu, A.Y.; Gurol, T.; Zhu, X.; Wirbisky-Hershberger, S.E.; Freeman, J.L.; Kasinski, A.L.; Deng, Q. Microrna-223 suppresses the canonical nf-kappab pathway in basal keratinocytes to dampen neutrophilic inflammation. Cell Rep. 2018, 22, 1810–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galindo-Villegas, J.; Montalban-Arques, A.; Liarte, S.; de Oliveira, S.; Pardo-Pastor, C.; Rubio-Moscardo, F.; Meseguer, J.; Valverde, M.A.; Mulero, V. Trpv4-mediated detection of hyposmotic stress by skin keratinocytes activates developmental immunity. J. Immunol. 2016, 196, 738–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berghmans, S.; Murphey, R.D.; Wienholds, E.; Neuberg, D.; Kutok, J.L.; Fletcher, C.D.; Morris, J.P.; Liu, T.X.; Schulte-Merker, S.; Kanki, J.P.; et al. Tp53 mutant zebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci. USA 2005, 102, 407–412. [Google Scholar] [CrossRef] [Green Version]
- Assefa, Z.; Van Laethem, A.; Garmyn, M.; Agostinis, P. Ultraviolet radiation-induced apoptosis in keratinocytes: On the role of cytosolic factors. Biochim. Biophys. Acta 2005, 1755, 90–106. [Google Scholar] [CrossRef]
- Skobowiat, C.; Dowdy, J.C.; Sayre, R.M.; Tuckey, R.C.; Slominski, A. Cutaneous hypothalamic-pituitary-adrenal axis homolog: Regulation by ultraviolet radiation. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E484–E493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noonan, F.P.; Dudek, J.; Merlino, G.; De Fabo, E.C. Animal models of melanoma: An hgf/sf transgenic mouse model may facilitate experimental access to uv initiating events. Pigment. Cell Res. 2003, 16, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Skobowiat, C.; Slominski, A.T. Ultraviolet b stimulates proopiomelanocortin signalling in the arcuate nucleus of the hypothalamus in mice. Exp. Dermatol. 2016, 25, 120–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skobowiat, C.; Postlethwaite, A.E.; Slominski, A.T. Skin exposure to ultraviolet b rapidly activates systemic neuroendocrine and immunosuppressive responses. Photochem. Photobiol. 2016, 93, 1008–1015. [Google Scholar] [CrossRef]
- Yang, H.M.; Ham, Y.M.; Yoon, W.J.; Roh, S.W.; Jeon, Y.J.; Oda, T.; Kang, S.M.; Kang, M.C.; Kim, E.A.; Kim, D.; et al. Quercitrin protects against ultraviolet b-induced cell death in vitro and in an in vivo zebrafish model. J. Photochem. Photobiol. B 2012, 114, 126–131. [Google Scholar] [CrossRef]
- Wang, Y.H.; Wen, C.C.; Yang, Z.S.; Cheng, C.C.; Tsai, J.N.; Ku, C.C.; Wu, H.J.; Chen, Y.H. Development of a whole-organism model to screen new compounds for sun protection. Mar. Biotechnol. N. Y. 2009, 11, 419–429. [Google Scholar] [CrossRef]
- Tsai, I.T.; Yang, Z.S.; Lin, Z.Y.; Wen, C.C.; Cheng, C.C.; Chen, Y.H. Flavone is efficient to protect zebrafish fins from uv-induced damage. Drug Chem. Toxicol. 2012, 35, 341–346. [Google Scholar] [CrossRef]
- Zeng, Z.; Richardson, J.; Verduzco, D.; Mitchell, D.L.; Patton, E.E. Zebrafish have a competent p53-dependent nucleotide excision repair pathway to resolve ultraviolet b-induced DNA damage in the skin. Zebrafish 2009, 6, 405–415. [Google Scholar] [CrossRef] [Green Version]
- Siegenthaler, B.; Defila, C.; Muzumdar, S.; Beer, H.D.; Meyer, M.; Tanner, S.; Bloch, W.; Blank, V.; Schafer, M.; Werner, S. Nrf3 promotes uv-induced keratinocyte apoptosis through suppression of cell adhesion. Cell Death Differ. 2018, 25, 1749–1765. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.M.; Timme-Laragy, A.R.; Goldstone, J.V.; McArthur, A.G.; Stegeman, J.J.; Smolowitz, R.M.; Hahn, M.E. Developmental expression of the nfe2-related factor (nrf) transcription factor family in the zebrafish, danio rerio. PLoS ONE 2013, 8, e79574. [Google Scholar] [CrossRef]
- Latonen, L.; Laiho, M. Cellular uv damage responses--functions of tumor suppressor p53. Biochim. Biophys Acta 2005, 1755, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Sandrini, J.Z.; Trindade, G.S.; Nery, L.E.; Marins, L.F. Time-course expression of DNA repair-related genes in hepatocytes of zebrafish (danio rerio) after uv-b exposure. Photochem. Photobiol. 2009, 85, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Koivukangas, V.; Kallioinen, M.; Autio-Harmainen, H.; Oikarinen, A. Uv irradiation induces the expression of gelatinases in human skin in vivo. Acta Derm. Venereol. 1994, 74, 279–282. [Google Scholar] [PubMed]
- Chatterjee, R.; Benzinger, M.J.; Ritter, J.L.; Bissett, D.L. Chronic ultraviolet b radiation-induced biochemical changes in the skin of hairless mice. Photochem. Photobiol. 1990, 51, 91–97. [Google Scholar] [CrossRef]
- Brenneisen, P.; Wenk, J.; Klotz, L.O.; Wlaschek, M.; Briviba, K.; Krieg, T.; Sies, H.; Scharffetter-Kochanek, K. Central role of ferrous/ferric iron in the ultraviolet b irradiation-mediated signaling pathway leading to increased interstitial collagenase (matrix-degrading metalloprotease (mmp)-1) and stromelysin-1 (mmp-3) mrna levels in cultured human dermal fibroblasts. J. Biol. Chem. 1998, 273, 5279–5287. [Google Scholar]
- Vincenti, M.P.; Brinckerhoff, C.E. Transcriptional regulation of collagenase (mmp-1, mmp-13) genes in arthritis: Integration of complex signaling pathways for the recruitment of gene-specific transcription factors. Arthritis Res. 2002, 4, 157–164. [Google Scholar] [CrossRef]
- Mignatti, P.; Rifkin, D.B. Biology and biochemistry of proteinases in tumor invasion. Physiol. Rev. 1993, 73, 161–195. [Google Scholar] [CrossRef]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef]
- Ishida, T.; Nakajima, T.; Kudo, A.; Kawakami, A. Phosphorylation of junb family proteins by the jun n-terminal kinase supports tissue regeneration in zebrafish. Dev. Biol. 2010, 340, 468–479. [Google Scholar] [CrossRef] [Green Version]
- Piechaczyk, M.; Farras, R. Regulation and Function of Junb in Cell Proliferation; Portland Press Ltd.: London, UK, 2008. [Google Scholar]
- Passegué, E.; Jochum, W.; Behrens, A.; Ricci, R.; Wagner, E.F. Junb can substitute for jun in mouse development and cell proliferation. Nat. Genet. 2002, 30, 158–166. [Google Scholar] [CrossRef]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, R.-Y.; Lin, C.-J.; Liang, S.-T.; Villalobos, O.; Villaflores, O.B.; Lou, B.; Lai, Y.-H.; Hsiao, C.-D. UVB Irradiation Induced Cell Damage and Early Onset of Junbb Expression in Zebrafish. Animals 2020, 10, 1096. https://doi.org/10.3390/ani10061096
Chen R-Y, Lin C-J, Liang S-T, Villalobos O, Villaflores OB, Lou B, Lai Y-H, Hsiao C-D. UVB Irradiation Induced Cell Damage and Early Onset of Junbb Expression in Zebrafish. Animals. 2020; 10(6):1096. https://doi.org/10.3390/ani10061096
Chicago/Turabian StyleChen, Rui-Yi, Chun-Ju Lin, Sung-Tzu Liang, Omar Villalobos, Oliver B. Villaflores, Bao Lou, Yu-Heng Lai, and Chung-Der Hsiao. 2020. "UVB Irradiation Induced Cell Damage and Early Onset of Junbb Expression in Zebrafish" Animals 10, no. 6: 1096. https://doi.org/10.3390/ani10061096