Map-A-Mole: Greenspace Area Influences the Presence and Abundance of the European Mole Talpa europaea in Urban Habitats

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
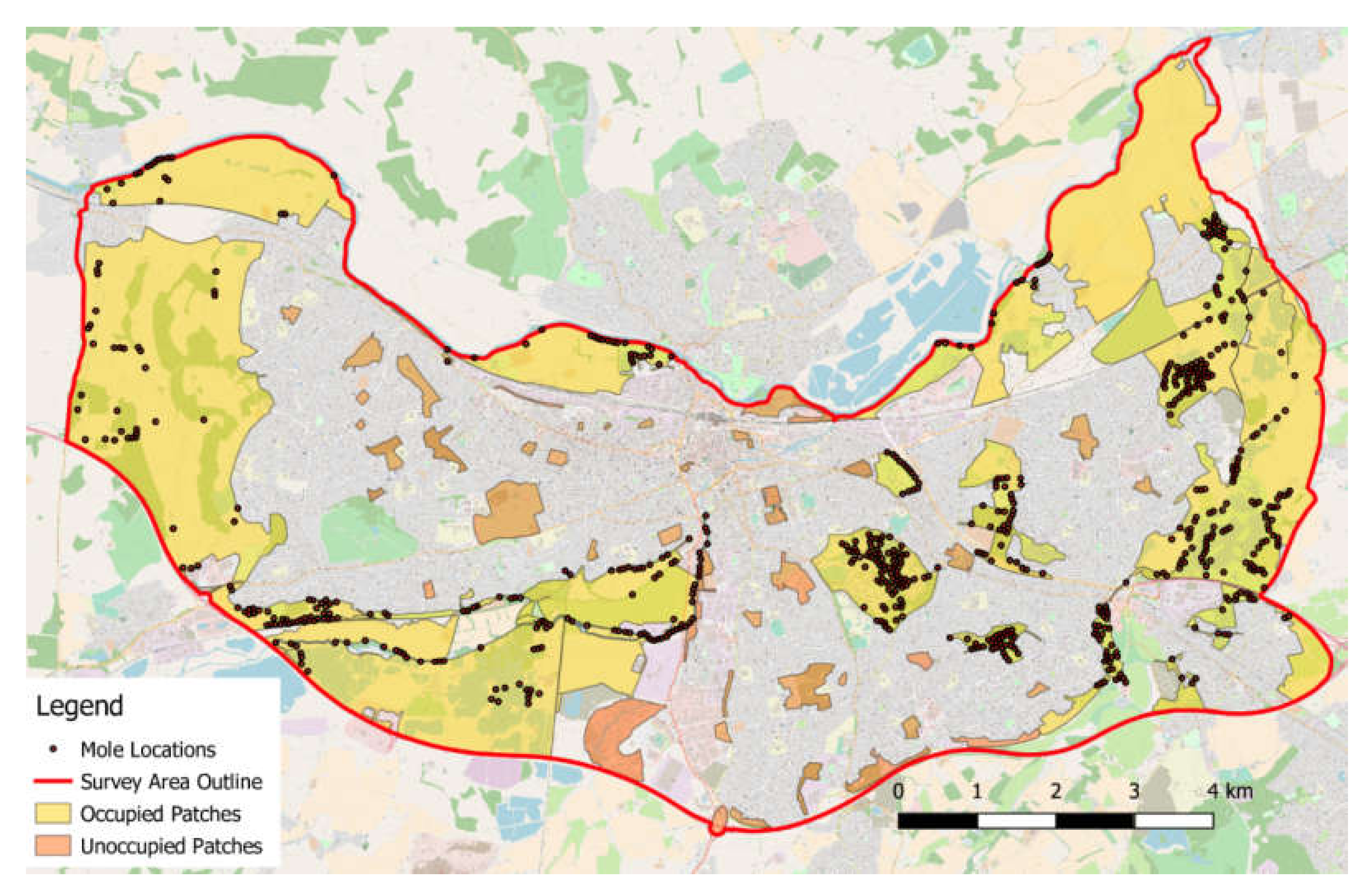
2.1. Study Sites
2.2. Mole Surveying
2.3. Other Variables
2.4. GIS Data Analysis
2.5. Statistical Analysis
3. Results
3.1. Surveying Results
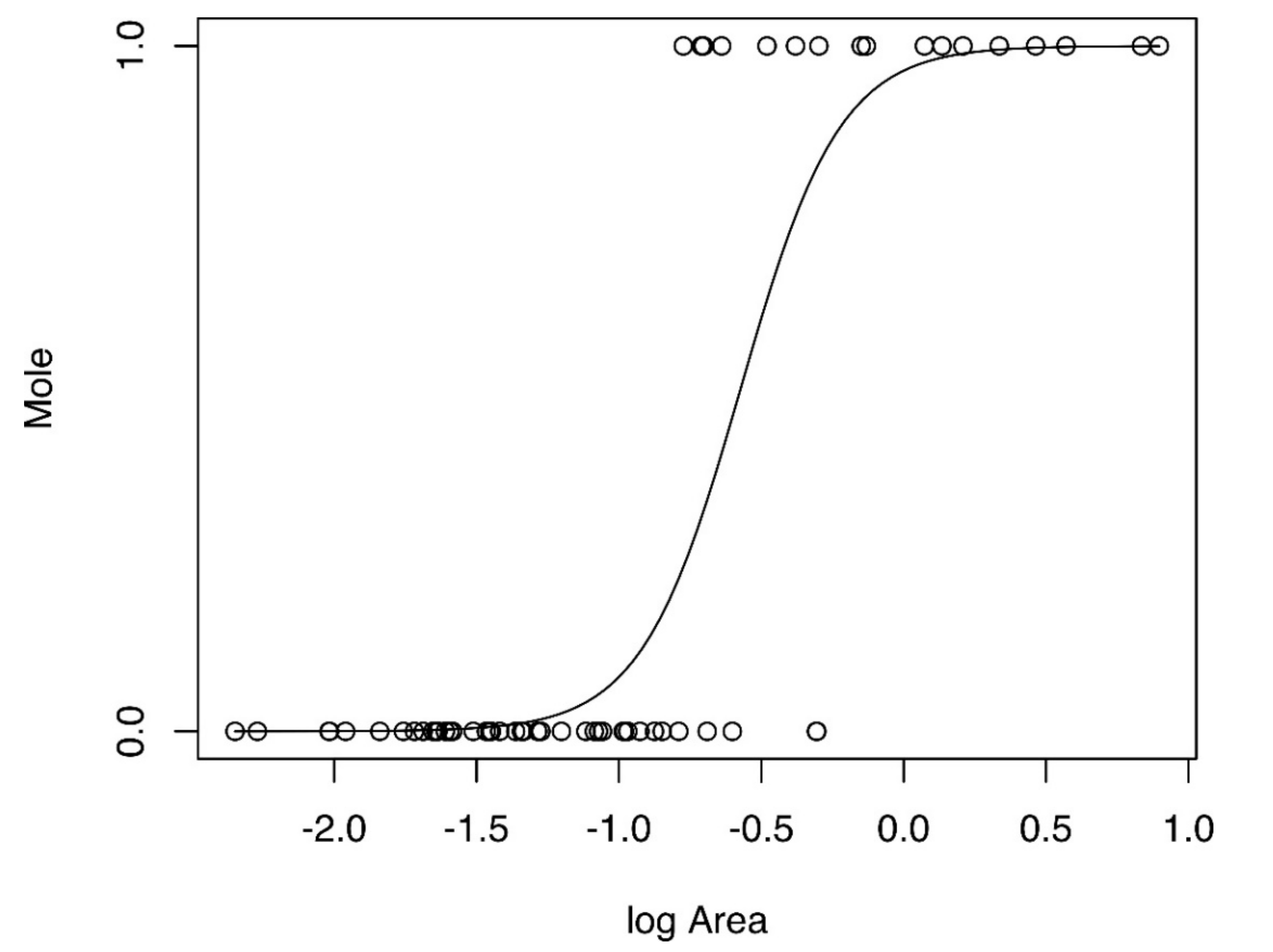
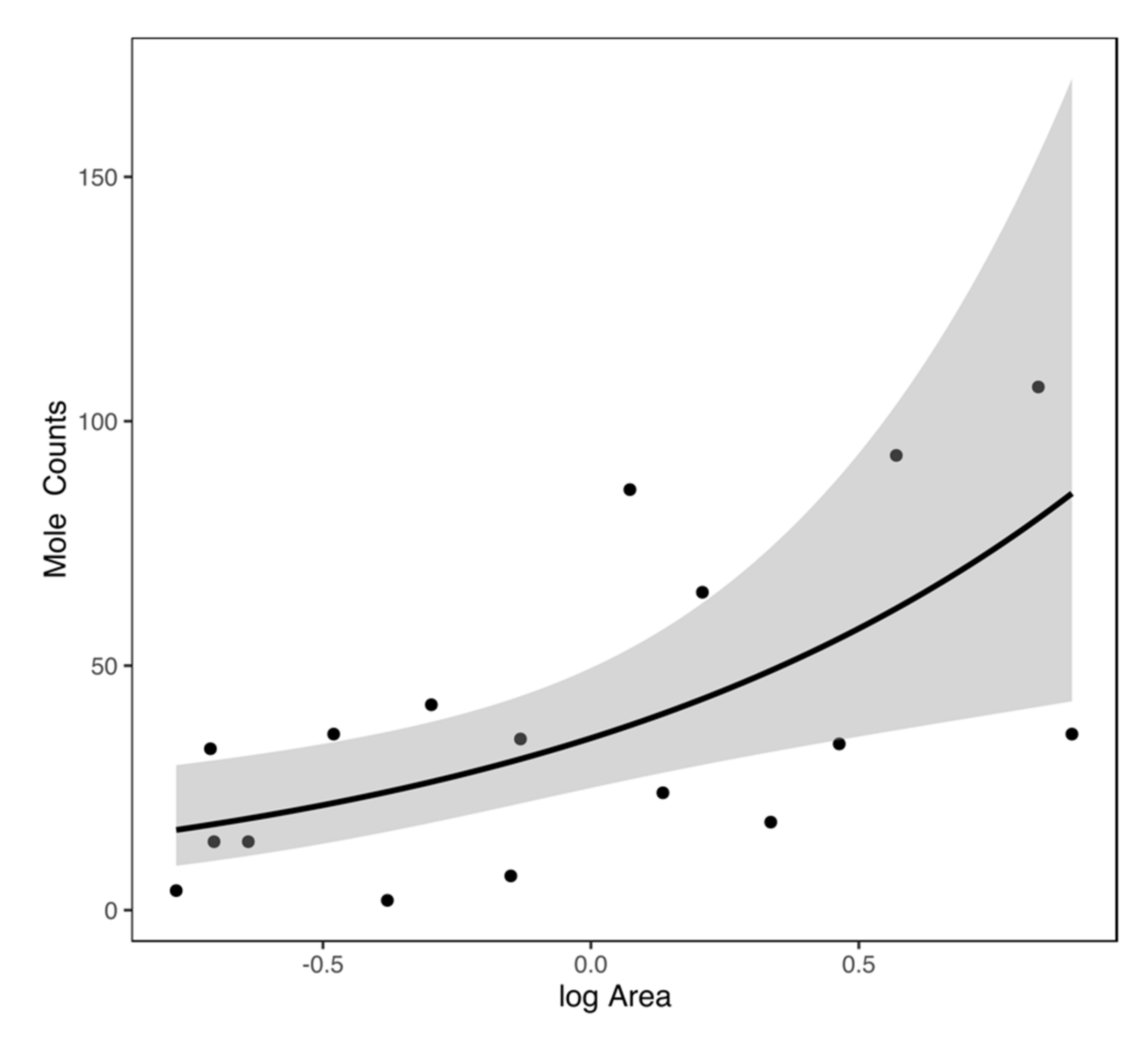
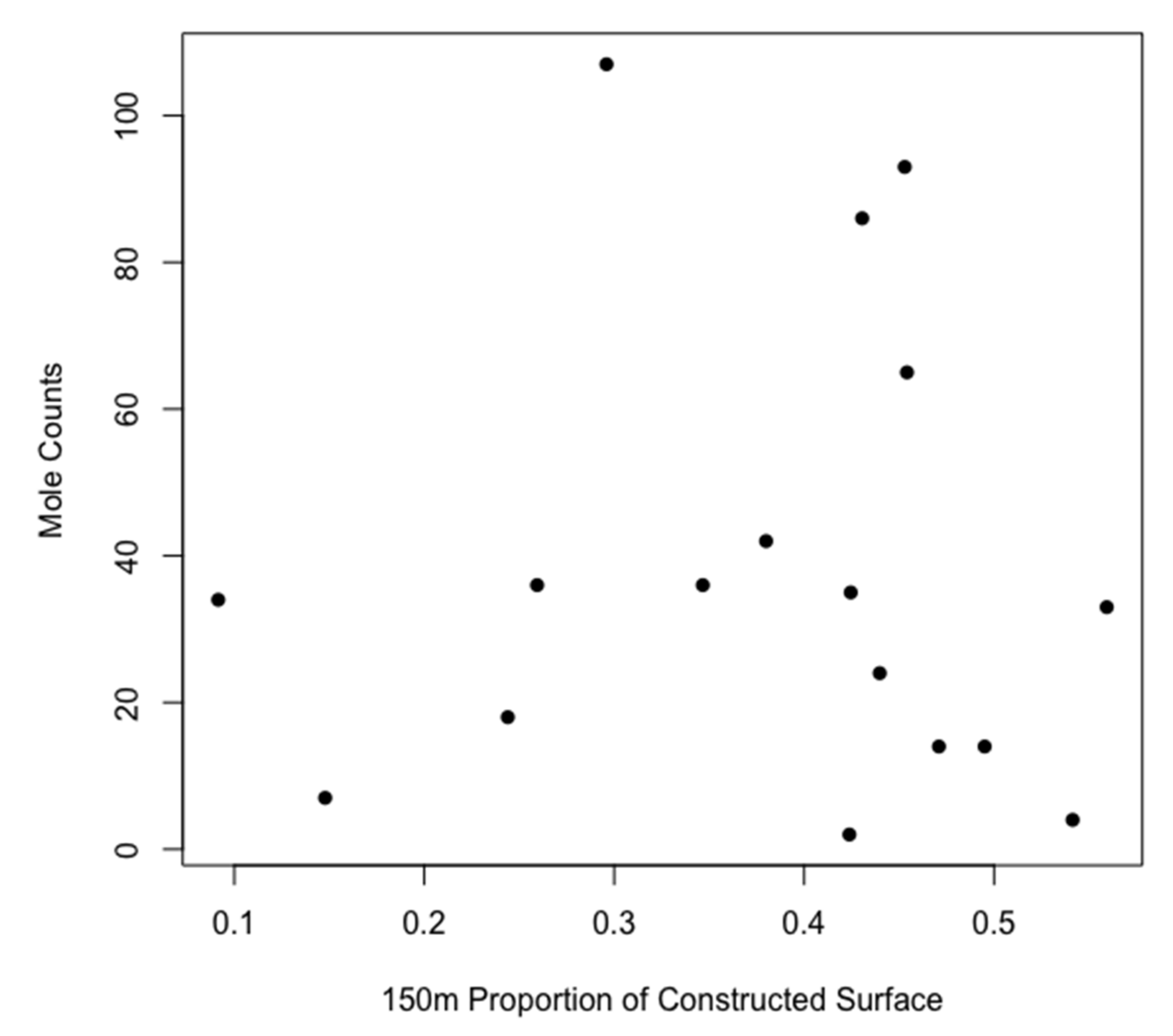
3.2. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McKinney, M.L. Urbanization, biodiversity, and conservation. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an ‘urban exploiter’? J. Biogeog. 2007, 34, 638–651. [Google Scholar] [CrossRef]
- Blair, R.B.; Johnson, E.M. Suburban habitats and their role for birds in the urban-rural habitat network: Points of local invasion and extinction? Landsc. Ecol. 2008, 23, 1157–1169. [Google Scholar] [CrossRef]
- Markovchick-Nicholls, L.; Regan, H.M.; Deustchman, D.H.; Widyanata, A.; Martin, B.; Noreke, L.; Ann Hunt, T. Relationships between human disturbance and wildlife land use in urban habitat fragments. Conserv. Biol. 2017, 22, 99–109. [Google Scholar] [CrossRef]
- Czech, B.; Krausman, P.R.; Devers, P.K. Economic associations among causes of species endangerment in the United States. BioScience 2000, 50, 593–601. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Juricic, E.; Jokimäki, J. A habitat island approach to conserving birds in urban landscapes: Case studies from southern and northern Europe. Biodivers. Conserv. 2001, 10, 2023–2043. [Google Scholar] [CrossRef]
- Crooks, K.R. Relative sensitivities of mammalian carnivores to habitat fragmentation. Conserv. Biol. 2002, 16, 488–502. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Rocha, E.A.; Fellowes, M.D.E. Does urbanization explain differences in interactions between an insect herbivore and its natural enemies and mutualists? Urban Ecosyst. 2018, 21, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Rocha, E.A.; Fellowes, M.D.E. Urbanization alters ecological interactions: Ant mutualists increase and specialist insect predators decrease on an urban gradient. Sci. Rep. 2020, 10, 6406. [Google Scholar] [CrossRef] [PubMed]
- Serrano, M.; Sanz, L.; Puig, J.; Pons, J. Landscape fragmentation caused by the transport network in Navarra (Spain): Two-scale analysis and landscape integration assessment. Landsc. Urban Plan. 2002, 58, 113–123. [Google Scholar] [CrossRef]
- Tillmann, J.E. Habitat fragmentation and ecological networks in Europe. GAIA 2005, 14, 119–123. [Google Scholar] [CrossRef]
- Laurance, W.F.; Gopalasamy, R.C.; Sloan, S.; O’Connell, C.S.; Mueller, N.D.; Goosem, M.; Venter, O.; Edwards, D.P.; Phalan, B.; Balmford, A.; et al. A global strategy for road building. Nature 2014, 513, 229–232. [Google Scholar] [CrossRef] [PubMed]
- Shepard, D.B.; Kuhns, A.R.; Dreslik, M.J.; Phillips, C.A. Roads as barriers to animal movement in fragmented landscapes. Anim. Conserv. 2008, 11, 288–296. [Google Scholar] [CrossRef]
- D’Amico, M.; Periquet, S.; Roman, J.; Revilla, E. Road avoidance responses determine the impact of heterogeneous road networks at a regional scale. J. Appl. Ecol. 2016, 53, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Angeoletto, F.H.S.; Fellowes, M.D.E.; Essi, L.; Santos, J.W.M.C.; Johann, J.M.; da Silva Leandro, D.; Mendonça, N.M. Ecología urbana y planificación: Una convergéncia ineludible. REGET 2019, 23, 17. [Google Scholar] [CrossRef]
- Rumble, H.; Angeoletto, F.; Connop, S.; Goddard, M.A.; Nash, C. Understanding and applying ecological principles in cities. In Planning Cities with Nature; Springer: Cham, Switzerland, 2019; pp. 217–234. [Google Scholar]
- Angeoletto, F.; Leandro, D.D.S.; Fellowes, M.D.E. The consequences of Brazil’s lack of transport planning is written in the blood of sparrows. Urban Geogr. 2019, 40, 1191–1197. [Google Scholar] [CrossRef]
- Orros, M.E.; Fellowes, M.D.E. Supplementary feeding of wild birds indirectly affects the local abundance of arthropod prey. Basic Appl. Ecol. 2012, 13, 286–293. [Google Scholar] [CrossRef]
- Orros, M.E.; Fellowes, M.D.E. Wild bird feeding in an urban area: Intensity, economics and numbers of individuals supported. Acta Ornithol. 2015, 50, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Angeoletto, F.; Sanz, J.P.R.; Albertin, R.; Silva, F.F.D. The grass is always greener on the other side of the fence: The flora in urban backyards of different social classes. Ambient. Soc. 2017, 20, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Jung, K.; Threlfall, C.G. Urbanization and its effects on bats—A global meta-analysis. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Springer: Cham, Switzerland, 2016; pp. 13–33. [Google Scholar] [CrossRef] [Green Version]
- IUCN. Talpa europaea. 2017. Available online: http://www.iucnredlist.org/details/41481/0 (accessed on 24 August 2018).
- The Mammal Society. Species Mole. Talpa europaea. 2008. Available online: http://www.mammal.org.uk/discover-mammals/species-mole/ (accessed on 7 June 2020).
- Mathews, F.; Kubasiewicz, L.M.; Gurnell, J.; Harrower, C.A.; McDonald, R.A.; Shore, R.F. A Review of the Population and Conservation Status of British Mammals; Natural England: Peterborough, UK, 2018. [Google Scholar]
- NBN Atlas. Talpa europaea Linnaeus, 1758, European Mole. Occurrence Records. 2015. Available online: https://records.nbnatlas.org/occurrences/search?q=lsid%3ANBNSYS0000005079&fq=year%3A%222015%22#tab_mapView (accessed on 7 June 2020).
- Loy, A.; Corti, C. Distribution of Talpa europea (Mammalia, Insectivora, Talpidae) in Europe: A biogeographic hypothesis based on morphometric data. Ital. J. Zool. 1996, 63, 277–284. [Google Scholar] [CrossRef]
- Godfrey, G.; Crowcroft, P. The Life of the Mole; Museum Press Limited: London, UK, 1960. [Google Scholar]
- Gorman, M.L.; Stone, R.D. The Natural History of Moles; Christopher Helm Publishers Limited: Bromley, UK, 1990. [Google Scholar]
- Goszczyński, J. Digging activity and estimation of the population density of moles. Acta Theriol. 1983, 28, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Funmilayo, O. Food consumption, preferences and storage in the Mole. Acta Theriol. 1979, 24, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Paoletti, M.G. The role of earthworms for assessment of sustainability and as bioindicators. Agric. Ecosyst. Environ. 1999, 74, 137–155. [Google Scholar] [CrossRef]
- Zurawska-Seta, E.; Barczak, T. The influence of field margins on the presence and spatial distribution of the European mole Talpa europaea L. within the agricultural landscape of northern Poland. Arch. Biol. Sci. 2012, 64, 971–980. [Google Scholar] [CrossRef]
- Gryz, J.; Krauze, D.; Goszczyński, J. The small mammals of Warsaw as based on the analysis of tawny owl (Strix aluco) pellets. Ann. Zool. Fenn. 2008, 45, 281–285. [Google Scholar] [CrossRef]
- Baker, S.E.; Maw, S.A.; Johnson, P.J.; Macdonald, D.W. Not in my backyard: Public perceptions of wildlife and ‘pest control’ in and around UK homes, and Local Authority ‘pest control’. Animals 2020, 10, 222. [Google Scholar] [CrossRef] [Green Version]
- Baker, S.E.; Ellwood, S.A.; Johnson, P.J.; Macdonald, D.W. Moles and mole control on British farms, amenities and gardens after strychnine withdrawal. Animals 2016, 6, 39. [Google Scholar] [CrossRef] [Green Version]
- QGIS.org. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2020. Available online: http://qgis.org (accessed on 1 May 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing. 2014. Available online: https://www.r-project.org/ (accessed on 22 April 2020).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2004. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Edwards, G.R.; Crawley, M.J.; Heard, M.S. Factors influencing molehill distribution in grassland: Implications for controlling the damage caused by molehills. J. Appl. Ecol. 1999, 36, 434–442. [Google Scholar] [CrossRef]
- Seifan, M.; Tielbörger, K.; Schloz-Murer, D.; Seifan, T. Contribution of molehill disturbances to grassland community composition along a productivity gradient. Acta Oecol. 2010, 36, 569–577. [Google Scholar] [CrossRef]
- Suárez-Seoane, S.; Virgós, E.; Terroba, O.; Pardavila, X.; Barea-Azcón, J.M. Scaling of species distribution models across spatial resolutions and extents along a biogeographic gradient. The case of the Iberian mole Talpa occidentalis. Ecography 2013, 37, 279–292. [Google Scholar] [CrossRef]
- Lee, E.J.; Rhim, S.J. Influence of vegetation area and edge length on mammals in urban woods. Anim. Cells Syst. 2017, 21, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Bowers, M.A.; Matter, S.F. Landscape ecology of mammals: Relationships between density and patch size. J. Mammal. 1997, 78, 999–1013. [Google Scholar] [CrossRef] [Green Version]
- Stone, R.D.; Gorman, M.L. Social organization of the European mole (Talpa europaea) and the Pyrenean desman (Galemys pyrenaicus). Mamm. Rev. 1985, 15, 35–42. [Google Scholar] [CrossRef]
- Nesterkova, D.V. Distribution and abundance of European mole (Talpa europaea L.) in areas affected by two Ural copper smelters. Russ. J. Ecol. 2013, 45, 429–436. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mole Presence | ||||
|---|---|---|---|---|
| Buffer Size | Explanatory Variable | Coefficient Value ± SE | p | AIC |
| 50 m | Intercept | 3.288 ± 1.286 | 0.010 | 27.291 |
| logArea | 5.742 ± 1.752 | 0.001 | ||
| 100 m | Intercept | 9.194 ± 4.706 | 0.051 | 26.286 |
| logArea | 5.312 ± 1.930 | 0.006 | ||
| 100 m CS | −12.846 ± 8.603 | 0.135 | ||
| 150 m | Intercept | 8.041 ± 4.217 | 0.056 | 26.889 |
| logArea | 5.235 ± 1.791 | 0.003 | ||
| 150 m CS | −10.443 ± 7.676 | 0.174 | ||
| Mole Counts | ||||
|---|---|---|---|---|
| Buffer Size | Explanatory Variable | Coefficient Value ± SE | p | AIC |
| 50 m | Intercept | 3.560 ± 0.174 | 0.000 | 156.43 |
| logArea | 0.986 ± 0.331 | 0.003 | ||
| 100 m | Intercept | 3.560 ± 0.174 | 0.000 | 156.43 |
| logArea | 0.986 ± 0.331 | 0.003 | ||
| 150 m | Intercept | 2.919 ± 0.558 | 0.000 | 156.39 |
| logArea | 1.035 ± 0.343 | 0.002 | ||
| logDistance NP | −0.417 ± 0.252 | 0.098 | ||
| 150 m CS | 3.090 ± 1.483 | 0.038 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fellowes, M.D.E.; Acquaah-Harrison, K.; Angeoletto, F.; Santos, J.W.M.C.; da Silva Leandro, D.; Rocha, E.A.; Pirie, T.J.; Thomas, R.L. Map-A-Mole: Greenspace Area Influences the Presence and Abundance of the European Mole Talpa europaea in Urban Habitats. Animals 2020, 10, 1097. https://doi.org/10.3390/ani10061097
Fellowes MDE, Acquaah-Harrison K, Angeoletto F, Santos JWMC, da Silva Leandro D, Rocha EA, Pirie TJ, Thomas RL. Map-A-Mole: Greenspace Area Influences the Presence and Abundance of the European Mole Talpa europaea in Urban Habitats. Animals. 2020; 10(6):1097. https://doi.org/10.3390/ani10061097
Chicago/Turabian StyleFellowes, Mark D.E., Kojo Acquaah-Harrison, Fabio Angeoletto, Jeater W.M.C. Santos, Deleon da Silva Leandro, Elise A. Rocha, Tara J. Pirie, and Rebecca L. Thomas. 2020. "Map-A-Mole: Greenspace Area Influences the Presence and Abundance of the European Mole Talpa europaea in Urban Habitats" Animals 10, no. 6: 1097. https://doi.org/10.3390/ani10061097