Influences of Immunocastration on Endocrine Parameters, Growth Performance and Carcass Quality, as Well as on Boar Taint and Penile Injuries

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Housing Conditions and Slaughtering
2.3. Sample Collection
2.3.1. Blood Sampling
2.3.2. Feces Sampling
2.3.3. Adipose Tissue Sampling
2.3.4. Sampling of Penis, Testis, and Accessory Glands
2.4. Analytical Methods
2.4.1. Determination of Boar Taint Compounds
2.4.2. Determination of Fatty Acid Composition
2.4.3. Quantification of GnRH Binding in Serum Samples
2.4.4. Determination of Testosterone Concentrations in Serum
2.4.5. Determination of Testosterone Concentrations in Feces
2.5. Statistical Analysis
3. Results
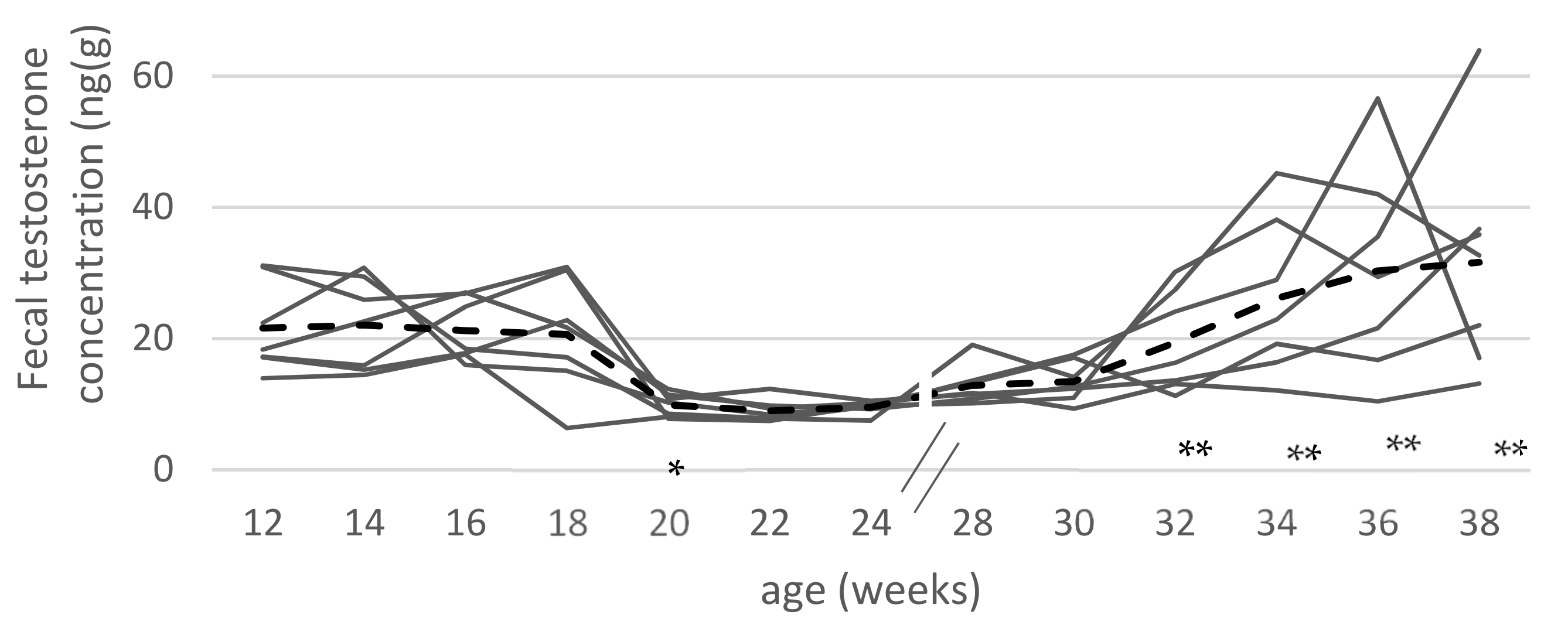
3.1. Fecal Testosterone Concentrations
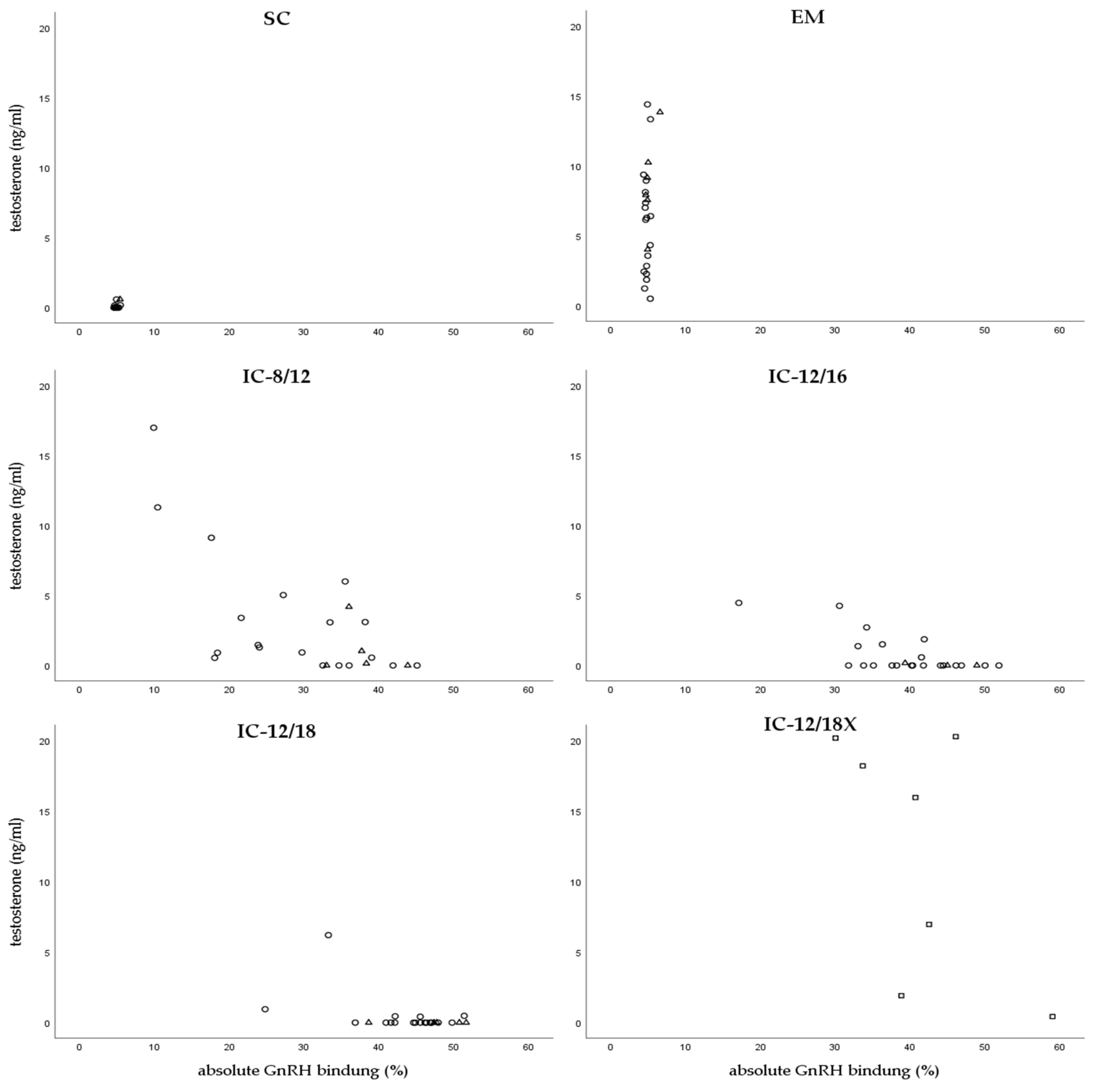
3.2. GnRH Binding and Serum Testosterone Concentrations
3.3. Fattening Performance and Carcass Quality
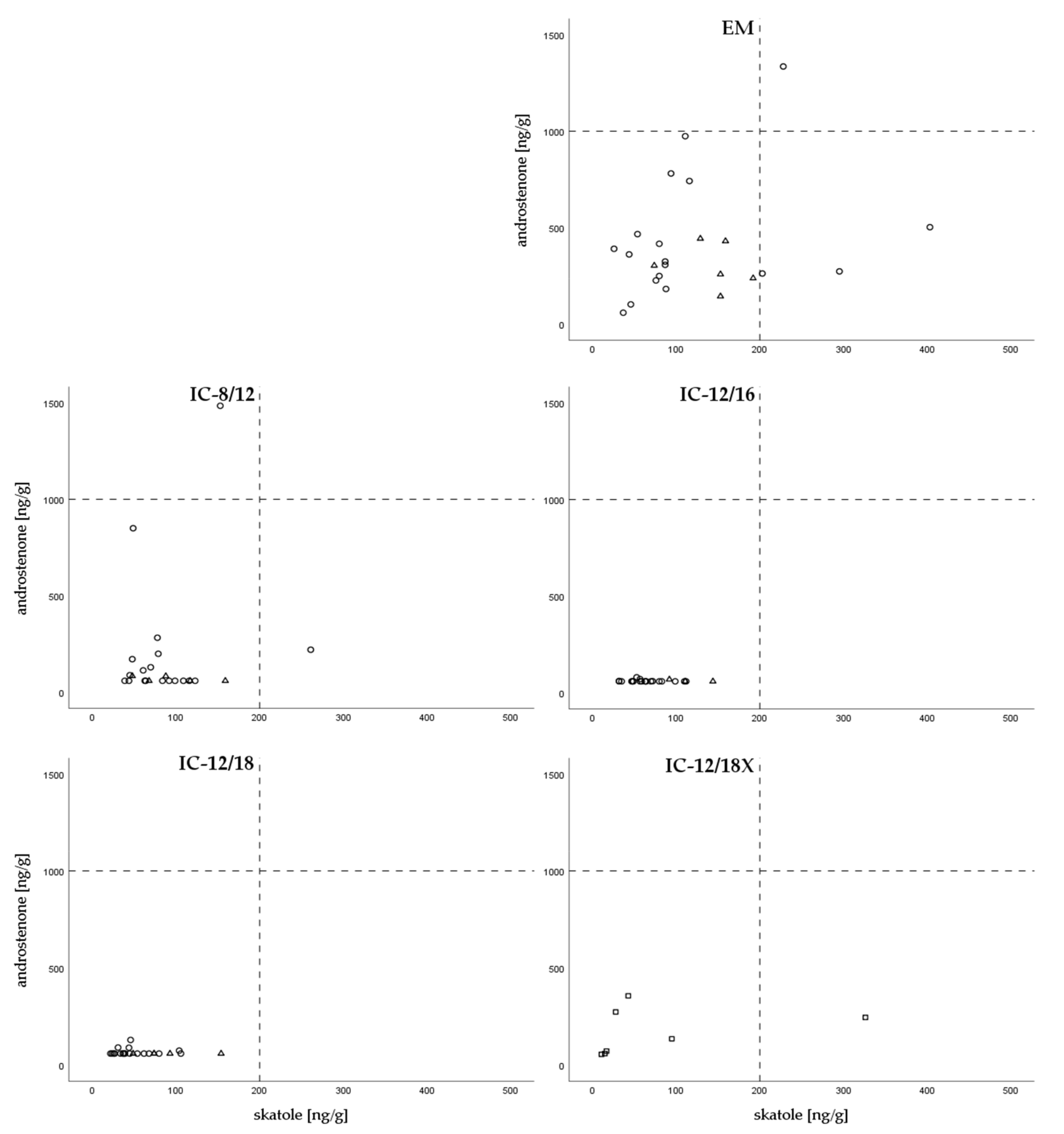
3.4. Levels of Boar Taint Compounds
3.5. Penile Injuries
3.6. Accessory Glands
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zöls, S.; Ritzmann, M.; Heinritzi, K. Einfluss von Schmerzmitteln bei der Kastration männlicher Ferkel. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 193–196. [Google Scholar] [PubMed]
- Prunier, A.; Bonneau, M.; Borell, E.H.; Cinotti, S.; Gunn, M.; Fredriksen, B.; Giersing, M.; Morton, D.B.; Tuyttens, F.A.M.; Velarde, A. A review of the welfare consequences of surgical castration in piglets and the evaluation of non-surgical methods. Anim. Welf. 2006, 15, 277–289. [Google Scholar]
- von Borell, E.; Baumgartner, J.; Giersing, M.; Jäggin, N.; Prunier, A.; Tuyttens, F.A.M.; Edwards, S.A. Animal welfare implications of surgical castration and its alternatives in pigs. Animal 2009, 3, 1488–1496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunshea, F.R.; Colantoni, C.; Howard, K.; McCauley, I.; Jackson, P.; Long, K.A.; Lopaticki, S.; Nugent, E.A.; Simons, J.A.; Walker, J.; et al. Vaccination of boars with a GnRH vaccine (Improvac) eliminates boar taint and increases growth performance. J. Anim. Sci. 2001, 79, 2524–2535. [Google Scholar] [CrossRef] [PubMed]
- Rydhmer, L.; Lundström, K.; Andersson, K. Immunocastration reduces aggressive and sexual behaviour in male pigs. Animal 2010, 4, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.; Claus, R. Involvement of glucocorticoids in testicular involution after active immunization of boars against GnRH. Reproduction 2004, 127, 275–283. [Google Scholar] [CrossRef] [Green Version]
- Andersson, K.; Brunius, C.; Zamaratskaia, G.; Lundstrom, K. Early vaccination with Improvac(R): Effects on performance and behaviour of male pigs. Animal 2012, 6, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Brunius, C.; Zamaratskaia, G.; Andersson, K.; Chen, G.; Norrby, M.; Madej, A.; Lundström, K. Early immunocastration of male pigs with Improvac®—Effect on boar taint, hormones and reproductive organs. Vaccine 2011, 29, 9514–9520. [Google Scholar] [CrossRef]
- Einarsson, S.; Brunius, C.; Wallgren, M.; Lundstrom, K.; Andersson, K.; Zamaratskaia, G.; Rodriguez-Martinez, H. Effects of early vaccination with Improvac(R) on the development and function of reproductive organs of male pigs. Anim. Reprod. Sci. 2011, 127, 50–55. [Google Scholar] [CrossRef] [Green Version]
- Barton-Gade, P.A. Meat and fat quality in boars, castrates and gilts. Livest. Prod. Sci. 1987, 16, 187–196. [Google Scholar] [CrossRef]
- Weiler, U.; Isernhagen, M.; Stefanski, V.; Ritzmann, M.; Kress, K.; Hein, C.; Zols, S. Penile Injuries in Wild and Domestic Pigs. Animals (Basel) 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Elsinghorst, P.W.; Bücking, M.; Tholen, E.; Petersen, B.; Wüst, M. Development of a Candidate Reference Method for the Simultaneous Quantitation of the Boar Taint Compounds Androstenone, 3α-Androstenol, 3β-Androstenol, Skatole, and Indole in Pig Fat by Means of Stable Isotope Dilution Analysis–Headspace Solid-Phase Microextraction–Gas Chromatography/Mass Spectrometry. Anal. Chem. 2011, 83, 6785–6791. [Google Scholar] [PubMed]
- Walstra, P.; Claudi-Magnussen, C.; Chevillon, P.; Seth, G.V.; Diestre, A.; Matthews, K.R.; Homer, D.B.; Bonneau, M. An international study on the importance of androstenone and skatole for boar taint: Levels of androstenone and skatole by country and season. Livest. Prod. Sci. 1999, 62, 15–28. [Google Scholar] [CrossRef]
- Schulte, E.; Weber, K. Schnelle Herstellung der Fettsäuremethylester aus Fetten mit Trimethylsulfoniumhydroxid oder Natriummethylat. Eur. J. Lipid Sci. Technol. 1989, 91, 181–183. [Google Scholar] [CrossRef]
- Schwalm, A.; Bauer, A.; Dederer, I.; Well, C.; Bussemas, R.; Weißmann, F. Effects of three genotypes and two roughages in organic heavy pig production for dry fermented sausage manufacture 2. Meat quality, fatty acid pattern, and product quality. Landbauforsch Volk 2013, 63, 273–284. [Google Scholar]
- Wesoly, R.; Jungbluth, I.; Stefanski, V.; Weiler, U. Pre-slaughter conditions influence skatole and androstenone in adipose tissue of boars. Meat Sci. 2015, 99, 60–67. [Google Scholar] [CrossRef]
- Conover, W.; Iman, R. [Rank Transformations as a Bridge between Parametric and Nonparametric Statistics]: Rejoinder. Am. Stat. 1981, 35, 124–129. [Google Scholar] [CrossRef]
- Bonastre, C.; Mitjana, O.; Tejedor, M.T.; Calavia, M.; Yuste, A.G.; Ubeda, J.L.; Falceto, M.V. Acute physiological responses to castration-related pain in piglets: The effect of two local anesthetics with or without meloxicam. Animal 2016, 10, 1474–1481. [Google Scholar] [CrossRef] [Green Version]
- Hay, M.; Vulin, A.; Génin, S.; Sales, P.; Prunier, A. Assessment of pain induced by castration in piglets: Behavioral and physiological responses over the subsequent 5 days. Appl. Anim. Behav. Sci. 2003, 82, 201–218. [Google Scholar] [CrossRef]
- Prunier, A.; Mounier, A.M.; Hay, M. Effects of castration, tooth resection, or tail docking on plasma metabolites and stress hormones in young pigs. J. Anim. Sci. 2005, 83, 216–222. [Google Scholar] [CrossRef]
- Puppe, B.; Schön, P.C.; Tuchscherer, A.; Manteuffel, G. Castration-induced vocalisation in domestic piglets, Sus scrofa: Complex and specific alterations of the vocal quality. Appl. Anim. Behav. Sci. 2005, 95, 67–78. [Google Scholar] [CrossRef]
- Hügel, T.; Zöls, S.; Palzer, A.; Kaufmann, S.; Langhoff, R.; Ritzmann, M.; Heinritzi, K. Einfluss einer GnRH-Vakzine (Improvac(R)) auf Gewichtszunahmen und Schlachtkorperqualitat von Ebern. Ein Feldversuch in Bayern. Tierarztl. Prax. 2011, 39, 353–357. [Google Scholar] [CrossRef]
- Aluwe, M.; Tuyttens, F.A.; Millet, S. Field experience with surgical castration with anaesthesia, analgesia, immunocastration and production of entire male pigs: Performance, carcass traits and boar taint prevalence. Animal 2015, 9, 500–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuyttens, F.A.M.; Vanhonacker, F.; Verhille, B.; De Brabander, D.; Verbeke, W. Pig producer attitude towards surgical castration of piglets without anaesthesia versus alternative strategies. Res. Vet. Sci. 2012, 92, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.-K.; Grosse Beilage, E.; Kanitz, E.; Puppe, B.; Traulsen, I.; Krieter, J. Influence of immunisation against GnRF on agonistic and mounting behaviour, serum testosterone concentration and body weight in male pigs compared with boars and barrows. Appl. Anim. Behav. Sci. 2012, 138, 28–35. [Google Scholar] [CrossRef]
- Zamaratskaia, G.; Andersson, H.K.; Chen, G.; Andersson, K.; Madej, A.; Lundstrom, K. Effect of a gonadotropin-releasing hormone vaccine (Improvac) on steroid hormones, boar taint compounds and performance in entire male pigs. Reprod. Domest. Anim. 2008, 43, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Claus, R.; Rottner, S.; Rueckert, C. Individual return to Leydig cell function after GnRH-immunization of boars. Vaccine 2008, 26, 4571–4578. [Google Scholar] [CrossRef]
- Rottner, S.; Claus, R. Return of testicular function after vaccination of boars against GnRH: Consequences on testes histology. Animal 2009, 3, 1279–1286. [Google Scholar] [CrossRef]
- Claus, R.; Hoffman, B.; Karg, H. Determination of 5 -androst-16-en-3-one, a boar taint steroid in pigs, with reference to relationships to testosterone. J. Anim. Sci. 1971, 33, 1293–1297. [Google Scholar] [CrossRef]
- Claus, R.; Weiler, U.; Herzog, A. Physiological aspects of androstenone and skatole formation in the boar-A review with experimental data. Meat Sci. 1994, 38, 289–305. [Google Scholar] [CrossRef]
- Giersing, M.; Lundstrom, K.; Andersson, A. Social effects and boar taint: Significance for production of slaughter boars (Sus scrofa). J. Anim. Sci. 2000, 78, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Bauer, A.; Lacorn, M.; Danowski, K.; Claus, R. Effects of immunization against GnRH on gonadotropins, the GH-IGF-I-axis and metabolic parameters in barrows. Animal 2008, 2, 1215–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claus, R.; Lacorn, M.; Danowski, K.; Pearce, M.C.; Bauer, A. Short-term endocrine and metabolic reactions before and after second immunization against GnRH in boars. Vaccine 2007, 25, 4689–4696. [Google Scholar] [CrossRef] [PubMed]
- Jaros, P.; Bürgi, E.; Stärk, K.D.C.; Claus, R.; Hennessy, D.; Thun, R. Effect of active immunization against GnRH on androstenone concentration, growth performance and carcass quality in intact male pigs. Livest. Prod. Sci. 2005, 92, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Einarsson, S.; Andersson, K.; Wallgren, M.; Lundstrom, K.; Rodriguez-Martinez, H. Short- and long-term effects of immunization against gonadotropin-releasing hormone, using Improvac, on sexual maturity, reproductive organs and sperm morphology in male pigs. Theriogenology 2009, 71, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Doran, E.; Whittington, F.W.; Wood, J.D.; McGivan, J.D. Cytochrome P450IIE1 (CYP2E1) is induced by skatole and this induction is blocked by androstenone in isolated pig hepatocytes. Chem. Biol. Interact. 2002, 140, 81–92. [Google Scholar] [CrossRef]
- Babol, J.; Squires, E.J.; Lundstrom, K. Hepatic metabolism of skatole in pigs by cytochrome P4502E1. J. Anim. Sci. 1998, 76, 822–828. [Google Scholar] [CrossRef]
- Doran, E.; Whittington, F.M.; Wood, J.D.; McGivan, J.D. Characterisation of androstenone metabolism in pig liver microsomes. Chem. Biol. Interact. 2004, 147, 141–149. [Google Scholar] [CrossRef]
- Fàbrega, E.; Velarde, A.; Cros, J.; Gispert, M.; Suárez, P.; Tibau, J.; Soler, J. Effect of vaccination against gonadotrophin-releasing hormone, using Improvac®, on growth performance, body composition, behaviour and acute phase proteins. Livest. Sci. 2010, 132, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Millet, S.; Gielkens, K.; De Brabander, D.; Janssens, G.P.J. Considerations on the performance of immunocastrated male pigs. Animal 2011, 5, 1119–1123. [Google Scholar] [CrossRef] [Green Version]
- Cronin, G.M.; Dunshea, F.R.; Butler, K.L.; McCauley, I.; Barnett, J.L.; Hemsworth, P.H. The effects of immuno- and surgical-castration on the behaviour and consequently growth of group-housed, male finisher pigs. Appl. Anim. Behav. Sci. 2003, 81, 111–126. [Google Scholar] [CrossRef]
- Pauly, C.; Spring, P.; O’Doherty, J.V.; Ampuero Kragten, S.; Bee, G. Growth performance, carcass characteristics and meat quality of group-penned surgically castrated, immunocastrated (Improvac®) and entire male pigs and individually penned entire male pigs. Animal 2009, 3, 1057–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sauer, F. Untersuchungen zu Schlachtkörperqualität und Ebergeruchsstoffen bei mit Einem GnRH-Analogon Geimpften, Chirurgisch Kastrierten und Intakten Männlichen Mastschweinen. Diss. med. vet., Veterinärmedizinische Fakultät der Universität Leipzig, Leipzig, Germany, 2015. [Google Scholar]
- Smithard, R.R.; Smith, W.C.; Ellis, M. A note on the fatty acid composition of backfat from boars in comparison with gilts and barrows. Anim. Sci. 1980, 31, 217–219. [Google Scholar] [CrossRef]
- Bee, G. Dietary conjugated linoleic acids affect tissue lipid composition but not de novo lipogenesis in finishing pigs. Animal 2001, 50, 383–399. [Google Scholar] [CrossRef] [Green Version]
- Kouba, M.; Sellier, P. A review of the factors influencing the development of intermuscular adipose tissue in the growing pig. Meat Sci. 2011, 88, 213–220. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Reiter, S.; Zöls, S.; Ritzmann, M.; Stefanski, V.; Weiler, U. Penile Injuries in Immunocastrated and Entire Male Pigs of One Fattening Farm. Animals (Basel) 2017, 7. [Google Scholar] [CrossRef] [Green Version]
- Hintze, S.; Scott, D.; Turner, S.; Meddle, S.L.; D’Eath, R.B. Mounting behaviour in finishing pigs: Stable individual differences are not due to dominance or stage of sexual development. Appl. Anim. Behav. Sci. 2013, 147, 69–80. [Google Scholar] [CrossRef]
- Batorek, N.; Candek-Potokar, M.; Bonneau, M.; Van Milgen, J. Meta-analysis of the effect of immunocastration on production performance, reproductive organs and boar taint compounds in pigs. Animal 2012, 6, 1330–1338. [Google Scholar] [CrossRef]
- Kubale, V.; Batorek, N.; Skrlep, M.; Prunier, A.; Bonneau, M.; Fazarinc, G.; Candek-Potokar, M. Steroid hormones, boar taint compounds, and reproductive organs in pigs according to the delay between immunocastration and slaughter. Theriogenology 2013, 79, 69–80. [Google Scholar] [CrossRef]
- Rottner, S. Wiederaufnahme der Hodenfunktion bei Ebern Nach Immunisierung gegen das Gonadotropin Releasing Hormon (GnRH) mit dem Kommerziellen Antigen IMPROVAC®. Diss. med. vet., Justus-Liebig-Universität Giessen, Giessen, Germany, 2012. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Group | n | Age (wk) at Vaccination | Weight (kg; Mean ± SD) at Vaccination | Number of Animals Slaughtered at an Age of | ||||
|---|---|---|---|---|---|---|---|---|
| V1 | V2 | V1 | V2 | 24 wk | 26 wk | 39 wk | ||
| SC | 24 | -- | -- | -- | -- | 4 | 19 | -- |
| EM | 24 | -- | -- | -- | -- | 6 | 18 | -- |
| IC-8/12 | 24 | 8 | 12 | 17.2 ± 2.1 | 31.3 ± 4.2 | 5 | 19 | -- |
| IC-12/16 | 24 | 12 | 16 | 31.9 ± 2.8 | 53.9 ± 5.0 | 3 | 21 | -- |
| IC-12/18 | 24 | 12 | 18 | 30.4 ± 3.2 | 65.3 ± 6.8 | 5 | 19 | -- |
| IC-12/18X | 8 | 12 | 18 | nd | nd | -- | -- | 7 |
| Groups | n | Weeks of Age | |||||
|---|---|---|---|---|---|---|---|
| 12th | 14th | 16th | 18th | 20th | 22nd | ||
| SC | 5 | 11.0 ± 1.6 a | 12.2 ± 2.9 ac | 10.2 ± 1.3 a | 10.5 ± 1.7 a | 11.6 ± 1.9 a | 10.8 ± 1.5 a |
| EM | 5 | 20.2 ± 17.6 | 26.2 ± 5.2 b | 34.7 ± 8.4 b | 31.5 ± 6.1 b | 34.4 ± 5.3 b | 37.2 ± 17.3 b |
| IC-8/12 | 5 | 15.0 ± 2.6 | 8.3 ± 1.4 a | 10.6 ± 1.7 ac | 9.1 ± 2.4 a | 8.6 ± 2.0 a | 9.3 ± 1.8 a |
| IC-12/16 | 5 | 20.3 ± 3.0 b | 20.0 ± 7.6 cb | 27.3 ± 9.3 d | 12.3 ± 2.4 a | 8.1 ± 1.3 a | 7.6 ± 2.0 a |
| IC-12/18 | 6 | 18.3 ± 5.8 | 19.2 ± 4.2 c | 21.7 ± 6.5 ad | 31.9 ± 8.9 b | 10.1 ± 2.4 a | 10.4 ± 2.3 a |
| Group | Second Vaccination | Slaughter | ||||
|---|---|---|---|---|---|---|
| n | Testosterone (ng/mL) | Absolute GnRH Binding (%) | n | Testosterone (ng/mL) | Absolute GnRH Binding (%) | |
| SC | - | nd | nd | 23 | 0.06 ± 0.17 a | 4.85 ± 0.24 a |
| EM | - | nd | nd | 24 | 6.63 ± 3.94 b | 4.88 ± 0.44 a |
| IC-8/12 | 5 | 0.90 ± 0.62 | 16.79 ± 7.00 | 24 | 2.88 ± 4.26 a | 30.24 ± 10.13 b |
| IC-12/16 | 5 | 3.22 ± 1.56 a | 20.31 ± 8.91 | 24 | 0.71 ± 1.34 b | 39.58 ± 7.58 c |
| IC-12/18 | 6 | 2.42 ± 1.33 a | 24.58 ± 3.36 | 24 | 0.35 ± 1.27 ab | 44.20 ± 6.12 c |
| IC-12/18X | 7 | 5.21 ± 3.57 | 17.51 ± 9.09 | 7 | 11.98 ± 8.66 c | 41.55 ± 9.41 c |
| Group Parameter | SC (n = 23) | EM (n = 24) | IC-8/12 (n = 24) | IC-12/16 (n = 24) | IC-12/18 (n = 24) | p-Value |
|---|---|---|---|---|---|---|
| Performance parameters | ||||||
| HCW (kg) | 94.4 ± 6.3 | 97.7 ± 4.9 | 97.4 ± 7.4 | 98.5 ± 6.4 | 98,3 ± 7.6 | n.s. |
| ADW (g/d) | 982.5 ± 69.1 | 1002.1 ± 83.6 | 1001.9 ± 73.9 | 989.2 ± 54.1 | 1028.8 ± 98.1 | n.s. |
| FCR (kg/kg) | 2.3 ± 0.1 a | 2.0 ± 0.1 b | 2.7 ± 0.4 c | 2.2 ± 0.1 a | 2.1 ± 0.1 ab | <0.001 |
| Fat quality | ||||||
| IMF (%) | 1.54 ± 0.33 a | 1.07 ± 0.29 b | 1.46 ± 0.31 a | 1.35 ± 0.46 ab | 1.53 ± 0.46 a | <0.001 |
| SFA (%) | 39.0 ± 1.7 ab | 37.9 ± 1.6 b | 39.9 ± 1.9 ca | 41.7 ± 2.5 d | 41.8 ± 2.1 d | <0.001 |
| MUFA (%) | 44.4 ± 1.7 a | 41.8 ± 2.3 b | 43.7 ± 1.6 ac | 43.2 ± 1.4 ac | 42.8 ± 1.2 bc | <0.001 |
| PUFA (%) | 16.6 ± 1.7 a | 20.4 ± 2.6 b | 16.4 ± 2.0 a | 15.2 ± 2.6 a | 15.5 ± 1.7 a | <0.001 |
| IV | 69.0 ± 2.6 a | 73.4 ± 3.2 b | 68.1 ± 3.1 ac | 65.4 ± 4.4 c | 65.5 ± 3.2 c | <0.001 |
| Group | n | Scars/Animal | Wounds/Animal | Animals with Lesions (%) | Animals with Severe Injuries (%) |
|---|---|---|---|---|---|
| EM | 24 | 7.92 ± 4.75a | 1.63 ± 1.69a | 91.67a | 12.5a |
| IC-8/12 | 24 | 0.17 ± 0.64b | 0.08 ± 0.28b | 16.67b | 0.00a |
| IC-12/16 | 24 | 0.54 ± 0.93cb | 0.00 ± 0.00cb | 29.17bc | 0.00a |
| IC-12/18 | 24 | 0.83 ± 1.17dc | 0.08 ± 0.41b | 41.67bc | 4.17a |
| IC-12/18X | 7 | 3.29 ± 3.15ad | 0.71 ± 1.25a | 71.43bc | 0.00a |
| Parameter | Group (n =) | r Spearman Rho (n =) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| IC-8/12 (21) | IC-12/16 (21) | IC-12/18 (18) | (p) | IC-12/18X (6) | EM (2) | Testost. | Androst. | ||
| Weight (g) | 128.3 ± 105.2 | 58.1 ± 26.2 | 68.6 ± 32.6 | 0.128 | 304.2 ± 169.2 | / | 0.626 * (65) | 0.387 * (64) | |
| Testes (cm) | wi | 4.4 ± 1.8 | 3.5 ± 0.9 | 3.8 ± 0.8 | 0.333 | 6.5 ± 2.0 | 5.0/7.5 | 0.550 * (67) | 0.379 * (66) |
| le | 6.5 ± 1.7 | 5.2 ± 0.86 | 5.8 ± 1.1 ~ | 0.210 | 10.3 ± 3.0 | 10.3/8.1 | 0.566 * (67) | 0.435 * (66) | |
| GlBu (cm) | wi | 2.1 ± 0.9a | 1.4 ± 0.6b | 1.6 ± 0.5ab | 0.039 | 2.8 ± 1.0 | 7.5/4.9 | 0.638 * (67) | 0.451 * (66) |
| le | 8.3 ± 2.5a | 6.4 ± 1.2b | 7.2 ± 1.3ab | 0.017 | 11.0 ± 3.5 | 12.2/8.9 | 0.584 * (68) | 0.478 * (67) | |
| GlVe (cm) | wi | 3.2 ± 2.0a | 1.7 ± 0.7b | 2.2 ± 0.9ab | 0.041 | 5.0 ± 2.8 | 5.0/3.1 | 0.561 * (68) | 0.442 * (67) |
| le | 4.9 ± 2.6 | 3.4 ± 0.9 | 3.8 ± 1.0 | 0.079 | 7.3 ± 3.1 | 11.3/7.9 | 0.554 * (68) | 0.396 * (67) | |
| GlPr (cm) | wi | 1.0 ± 0.8 | 0.7 ± 0.4 | 0.9 ± 0.4 | 0.213 | 2.2 ± 1.0 | 6.2/6.9 | 0.486 * (68) | 0.265 * (67) |
| le | 2.5 ± 0.8 | 2.2 ± 1.0 | 2.3 ± 0.7 | 0.440 | 4.0 ± 1.1 | 3.1/3.3 | 0.534 * (68) | 0.362 * (67) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zoels, S.; Reiter, S.; Ritzmann, M.; Weiß, C.; Numberger, J.; Schütz, A.; Lindner, P.; Stefanski, V.; Weiler, U. Influences of Immunocastration on Endocrine Parameters, Growth Performance and Carcass Quality, as Well as on Boar Taint and Penile Injuries. Animals 2020, 10, 346. https://doi.org/10.3390/ani10020346
Zoels S, Reiter S, Ritzmann M, Weiß C, Numberger J, Schütz A, Lindner P, Stefanski V, Weiler U. Influences of Immunocastration on Endocrine Parameters, Growth Performance and Carcass Quality, as Well as on Boar Taint and Penile Injuries. Animals. 2020; 10(2):346. https://doi.org/10.3390/ani10020346
Chicago/Turabian StyleZoels, Susanne, Simon Reiter, Mathias Ritzmann, Christine Weiß, Jasmin Numberger, Aneka Schütz, Peter Lindner, Volker Stefanski, and Ulrike Weiler. 2020. "Influences of Immunocastration on Endocrine Parameters, Growth Performance and Carcass Quality, as Well as on Boar Taint and Penile Injuries" Animals 10, no. 2: 346. https://doi.org/10.3390/ani10020346