Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics Statement
2.2. Experimental Design and Management
2.3. Preparation of Bacterial Strain and MccJ25
2.4. Determination of Minimum Inhibitory Concentration
2.5. Sample Collection and Preparation
2.6. E. coli, Lactid Acid Bacteria, Bifidobacterium Count, and Salmonella Infection Rate in Feces
2.7. Small Intestine Morphology
2.8. Determination of Immunomodulatory factor Concentrations in Serum
2.9. Statistical Analysis
3. Results
3.1. Minimum Inhibitory Concentration (MIC) of MccJ25 Against E. coli AZ1 and Salmonella CVCC519
3.2. Growth Performance
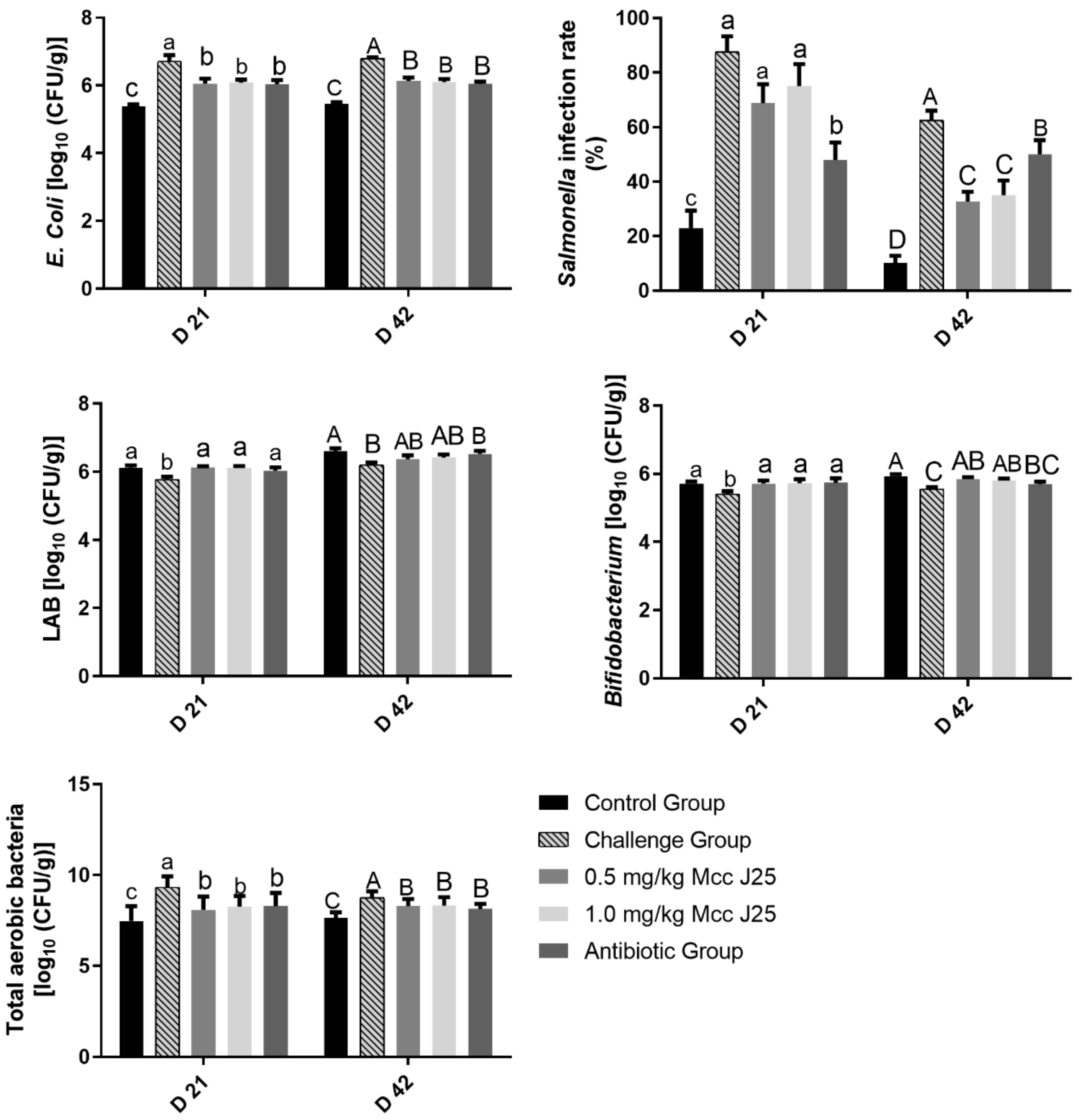
3.3. Count of Feces Bacteria and Salmonella Infection Rate
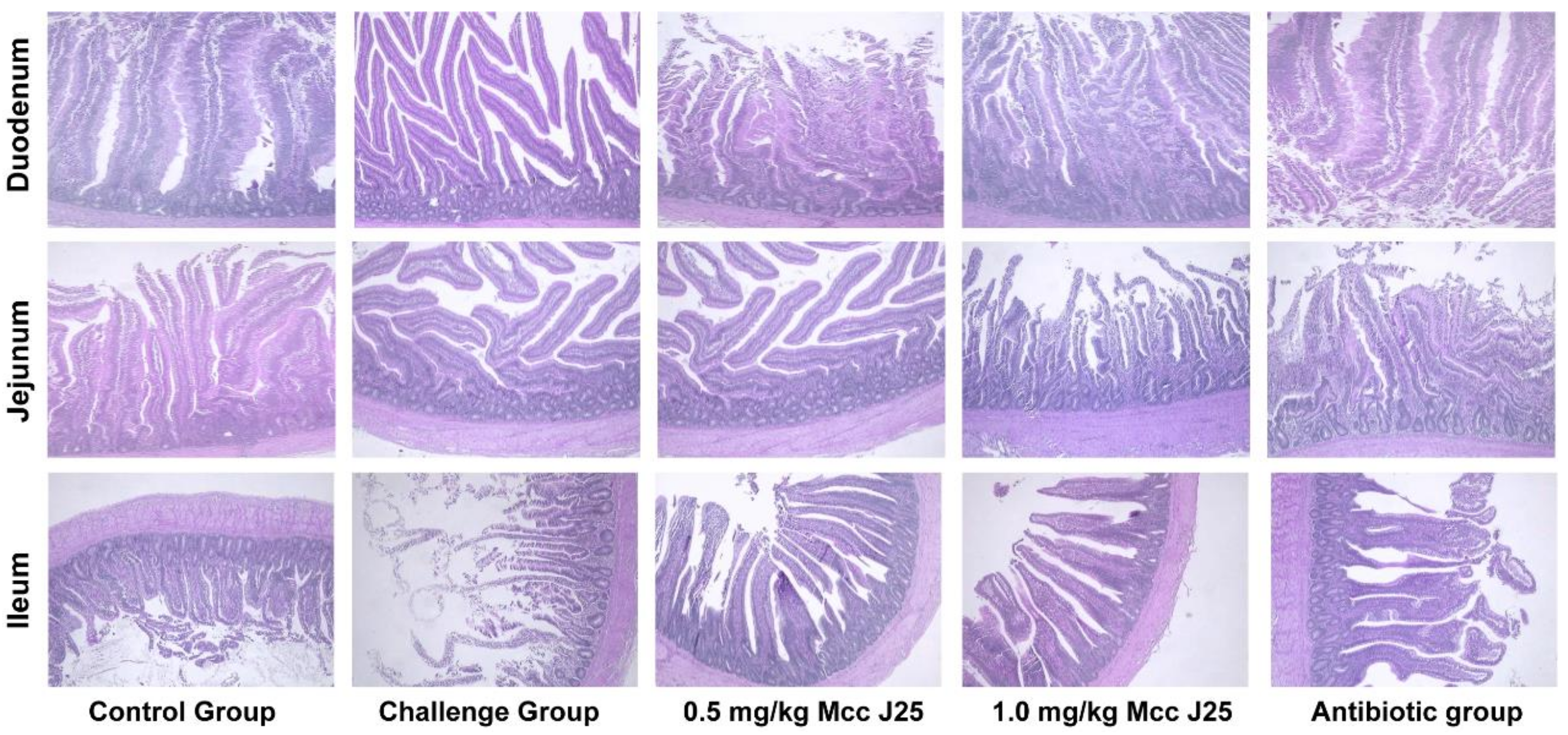
3.4. Intestinal Morphology
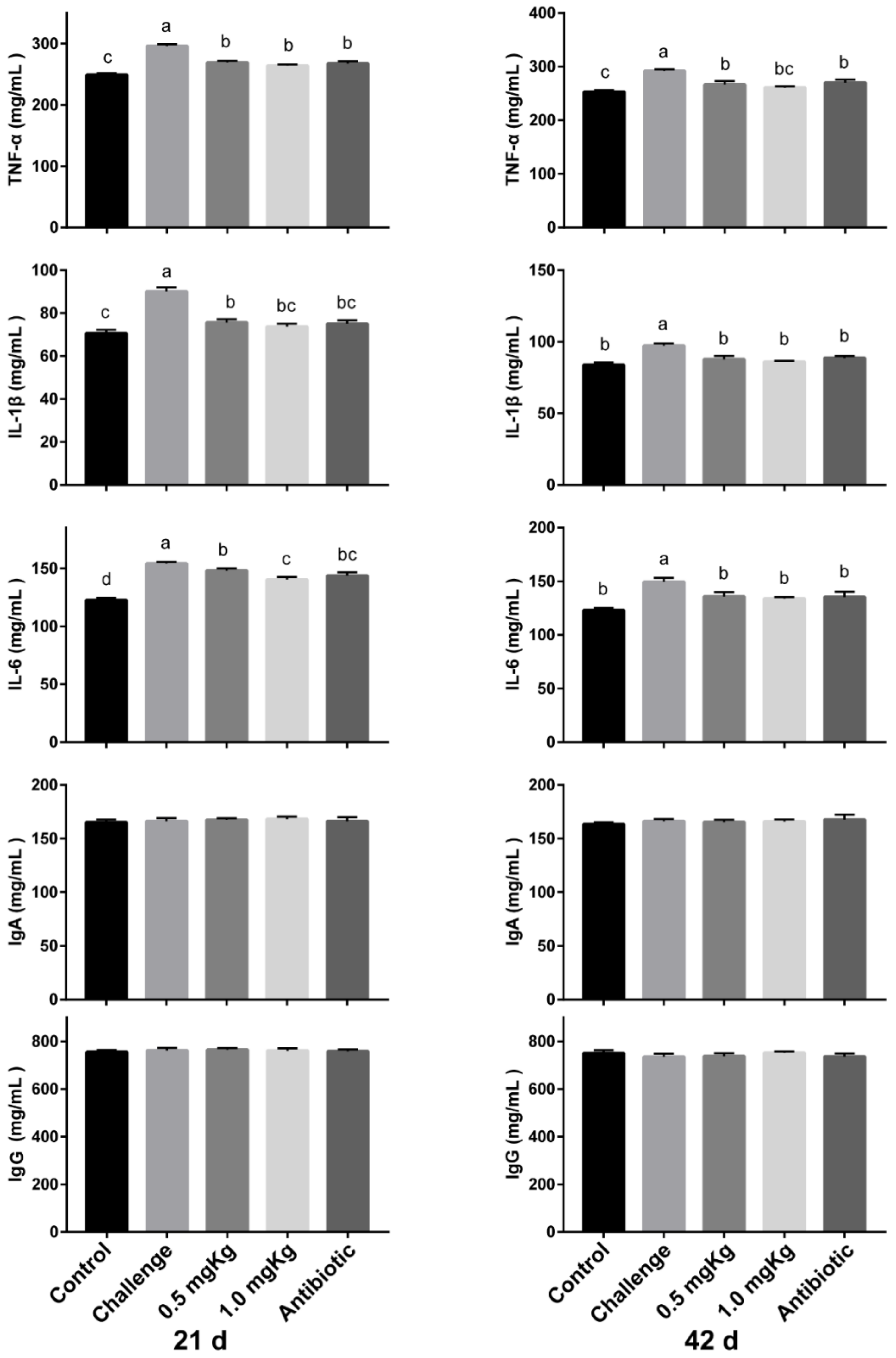
3.5. Immune Factor Concentration in Serum
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Matin, M.A.; Islam, M.A.; Khatun, M.M. Prevalence of colibacillosis in chickens in greater Mymensingh district of Bangladesh. Vet. World 2017, 10, 29–33. [Google Scholar] [CrossRef] [Green Version]
- Levy, S. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. Suppl. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Boman, H.G.; Nilsson, I.; Rasmuson, B. Inducible Antibacterial Defence System in Drosophila. Nature 1972, 237, 232–235. [Google Scholar] [CrossRef]
- Bradshaw, J. Cationic antimicrobial peptides: Issues for potential clinical use. Biodrugs Clin. Immunother. Biopharm. Gene Ther. 2003, 17, 233–240. [Google Scholar] [CrossRef]
- Blazyk, J. A Novel Linear Amphipathic β-Sheet Cationic Antimicrobial Peptide with Enhanced Selectivity for Bacterial Lipids. J. Biol. Chem. 2001, 276, 27899–27906. [Google Scholar] [CrossRef] [Green Version]
- Salomón, R.A.; Farías, R.N. Microcin 25, a novel antimicrobial peptide produced by Escherichia coli. J. Bacteriol. 1992, 174, 7428–7435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, F.E.; Vincent, P.A.; Zenoff, A.M.; Salomon, R.A.; Farias, R.N. Efficacy of microcin J25 in biomatrices and in a mouse model of Salmonella infection. J. Antimicrob. Chemother. 2007, 59, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.T.; Ding, X.L.; Li, N.; Zhang, X.Y.; Zeng, X.F.; Wang, S.; Liu, H.B.; Wang, Y.M.; Jia, H.M.; Qiao, S.Y. Dietary supplemented antimicrobial peptide microcin J25 improves the growth performance, apparent total tract digestibility, fecal microbiota, and intestinal barrier function of weaned pigs. J. Anim. Sci. 2017, 9, 5064–5076. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Shang, L.; Zeng, X.; Li, N.; Liu, H.; Cai, S.; Huang, S.; Wang, G.; Wang, Y.M.; Song, Q.L.; et al. Risks Related to High-Dosage Recombinant Antimicrobial Peptide Microcin J25 in Mice Model: Intestinal Microbiota, Intestinal Barrier Function, and Immune Regulation. J. Agric. Food Chem. 2018, 66, 11301–11310. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Q.; Zeng, X.; Ye, Q.; Huang, S.; Yu, H.; Yang, T.; Qiao, S. Use of the antimicrobial peptide sublancin with combined antibacterial and immunomodulatory activities to protect against methicillin-resistant Staphylococcus aureus infection in mice. J. Agric. Food Chem. 2017, 65, 8595–8605. [Google Scholar] [CrossRef]
- Liu, H.; Hou, C.; Wang, G.; Jia, H.; Yu, H.; Zeng, X.; Thacker, P.A.; Zhang, G.; Qiao, S. Lactobacillus reuteri I5007 Modulates Intestinal Host Defense Peptide Expression in the Model of IPEC-J2 Cells and Neonatal Piglets. Nutrients 2017, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emami, N.K.; Daneshmand, A.; Naeini, S.Z.; Graystone, E.N.; Broom, L.J. Effects of commercial organic acid blends on male broilers challenged with E. coli K88: Performance, microbiology, intestinal morphology, and immune response. Poult. Sci. 2017, 96, 3254–3263. [Google Scholar] [CrossRef] [PubMed]
- Jazi, V.; Foroozandeh, A.D.; Toghyani, M.; Dastar, B.; Rezaie Koochaksaraie, R.; Toghyani, M. Effects of Pediococcus acidilactici, mannan-oligosaccharide, butyric acid and their combination on growth performance and intestinal health in young broiler chickens challenged with Salmonella Typhimurium. Poult. Sci. 2018, 97, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Vandeplas, S.; Dauphin, R.D.; Thiry, C.; Beckers, Y.; Welling, G.W.; Thonart, P.; Thewis, A. Efficiency of a Lactobacillus plantarum-xylanase combination on growth performances, microflora populations, and nutrient digestibilities of broilers infected with Salmonella Typhimurium. Poult. Sci. 2009, 88, 1643–1654. [Google Scholar] [CrossRef]
- Wang, Y. Effect of lactoferrin on the growth performance, intestinal morphology, and expression of PR-39 and protegrin-1 genes in weaned piglets. J. Anim. Sci. 2006, 84, 2636–2641. [Google Scholar] [CrossRef]
- Kierończyk, B.; Rawski, M.; Mikołajczak, Z.; Świątkiewicz, S.; Józefiak, D. Nisin as a Novel Feed Additive: The Effects on Gut Microbial Modulation and Activity, Histological Parameters, and Growth Performance of Broiler Chickens. Animals 2020, 10, 101. [Google Scholar] [CrossRef] [Green Version]
- Ohh, S.H.; Shinde, P.L.; Jin, Z.; Choi, J.Y.; Hahn, T.-W.; Lim, H.T.; Kim, G.Y.; Park, Y.; Hahm, K.-S.; Chae, B.J. Potato (Solanum tuberosum L. cv. Gogu valley) protein as an antimicrobial agent in the diets of broilers. Poult Sci. 2009, 88, 1227–1234. [Google Scholar] [CrossRef]
- Yoon, J.H.; Ingale, S.L.; Kim, J.S.; Kim, K.H.; Lee, S.H.; Park, Y.K.; Kwon, I.K.; Chae, B.J. Effects of dietary supplementation of antimicrobial peptide-A3 on growth performance, nutrient digestibility, intestinal and fecal microflora and intestinal morphology in weanling pigs. Anim. Feed. Sci. Technol. 2012, 177, 98–107. [Google Scholar] [CrossRef]
- Jin, Z.; Yang, Y.X.; Choi, J.Y.; Shinde, P.L.; Yoon, S.Y.; Hahn, T.-W.; Lim, H.T.; Park, Y.K.; Hahm, K.S.; Joo, J.W. Effects of potato (Solanum tuberosum L. cv. Golden valley) protein having antimicrobial activity on the growth performance, and intestinal microflora and morphology in weanling pigs. Anim. Feed Sci. Technol. 2008, 140, 139–154. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Shan, T.-Z.; Xu, Z.-R.; Feng, J.; Wang, Z.-Q. Effects of the lactoferrin (LF) on the growth performance, intestinal microflora and morphology of weanling pigs. Anim. Feed Sci. Technol. 2007, 135, 263–272. [Google Scholar] [CrossRef]
- Fuller, M.F.; Benevenga, N.; Fisher, C.; Lall, S.; Phillips, C.J.C. The Encyclopedia of Farm Animal Nutrition; Rowett Research Institute: Aberdeen, UK, 2004. [Google Scholar]
- Xu, Z.R.; Hu, C.H.; Xia, M.S.; Zhan, X.A.; Wang, M.Q. Effects of dietary fructooligosaccharide on digestive enzyme activities, intestinal microflora and morphology of male broilers. Poult. Sci. 2003, 82, 1030–1036. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, F.; Huang, Z.; Liu, H.; Xie, C.; Zhang, J.; Thacker, P.A.; Qiao, S. Effects of the antimicrobial peptide cecropin AD on performance and intestinal health in weaned piglets challenged with Escherichia coli. Peptides 2012, 35, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Yin, Y.; Zhang, Y.; Huang, R.; Sun, Z.; Li, T.; Chu, W.; Kong, X.; Li, L.; Geng, M.; et al. Effects of dietary supplementation with an expressed fusion peptide bovine lactoferricin-lactoferrampin on performance, immune function and intestinal mucosal morphology in piglets weaned at age 21 d. Br. J. Nutr. 2009, 101, 998–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.K.; Croom, J.; Christensen, V.L.; Black, B.L.; Eisen, E.J. Jejunal Glucose Uptake and Oxygen Consumption in Turkey Poults Selected for Rapid Growth. Poult. Sci. 1997, 76, 1738–1745. [Google Scholar] [CrossRef]
- Samanya, M.; Yamauchi, K. Histological alterations of intestinal villi in chickens fed dried Bacillus subtilis var. natto. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 2002, 133, 95–104. [Google Scholar] [CrossRef]
- Tayal, V.; Kalra, B.S. Cytokines and anti-cytokines as therapeutics—An update. Eur. J. Pharmacol. 2008, 579, 1–12. [Google Scholar] [CrossRef]
- Hing, T.C.; Ho, S.; Shih, D.Q.; Ichikawa, R.; Cheng, M.; Chen, J.; Chen, X.; Law, I.; Najarian, R.; Kelly, C.P.; et al. The antimicrobial peptide cathelicidin modulates Clostridium difficile-associated colitis and toxin A-mediated enteritis in mice. Gut 2013, 62, 1295–1305. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Item | (1–21 d) | (21–42 d) |
|---|---|---|
| Ingredients (%) | ||
| Corn | 55.97 | 61.37 |
| Soybean meal | 37.63 | 31.88 |
| Soybean oil | 2.37 | 3.13 |
| Limestone | 1.27 | 1.21 |
| NaCl | 0.35 | 0.35 |
| CaHPO4 | 1.75 | 1.49 |
| DL-Methionine | 0.2 | 0.12 |
| HCl L-Lys·HCl | 0.02 | 0.01 |
| Antioxygen | 0.02 | 0.02 |
| Vitamin premix 1 | 0.02 | 0.02 |
| Choline chloride 1 | 0.2 | 0.2 |
| Trace mineral premix 2 | 0.2 | 0.2 |
| Total | 100 | 100 |
| Analyzed nutritional value (%) 3 | ||
| Metabolic energy/(MJ/kg) | 11.76 | 12.22 |
| Crude protein | 21.36 | 19.87 |
| Crude fat | 4.83 | 5.02 |
| Crude fiber | 3.46 | 3.09 |
| Ca | 1.01 | 0.91 |
| Total phosphorus | 0.60 | 0.54 |
| Lys | 1.14 | 1.02 |
| Met | 0.50 | 0.41 |
| MccJ25 μg/mL | 0 | 0.0625 | 0.125 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| E. coli AZ1 | + | + | + | + | − | − | − | − | − | − | − |
| Salmonella CVCC519 | + | + | + | + | − | − | − | − | − | − | − |
| E. coli ATCC25922 | + | + | + | + | + | − | − | − | − | − | − |
| Item | Treatment | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Control Group | Challenge Group | 0.5 mg/kg MccJ25 | 1.0 mg/kg MccJ25 | Antibiotic Group | |||
| BW (g) | |||||||
| days 1–21 | 872 a | 823.25 d | 857.67 ab | 840.24 c | 846.18 bc | 5.32 | <0.01 |
| days 22–42 | 1619.09 | 1576.98 | 1604.12 | 1623.53 | 1603.51 | 14.37 | 0.19 |
| days 1–42 | 2491.1 a | 2400.2 c | 2461.8 ab | 2463.8 ab | 2449.7 bc | 14.98 | <0.01 |
| Feed-to-Gain ratio (G:F) | |||||||
| days 1–21 | 1.36 b | 1.42 a | 1.39 ab | 1.4 ab | 1.37 b | 0.014 | 0.04 |
| days 1–42 | 1.76 c | 1.9 a | 1.82 b | 1.83 b | 1.84 b | 0.01 | <0.01 |
| Feed Intake (kg) | |||||||
| days 1–21 | 1.26 | 1.24 | 1.26 | 1.25 | 1.23 | 0.02 | 0.63 |
| days 1–42 | 4.47 | 4.66 | 4.57 | 4.60 | 4.60 | 1.49 | 0.45 |
| Mortality (%) | |||||||
| days 1–14 | 0.32 b | 2.72 a | 1.12 b | 1.28 b | 1.16 b | 0.32 | <0.01 |
| days 1–21 | 0.48 c | 4.01 a | 2.08 b | 2.08 b | 1.71 b | 0.42 | <0.01 |
| days 1–42 | 0.52 c | 5.03 a | 2.78 b | 2.43 b | 2.43 b | 0.41 | <0.01 |
| Item | Experimental Treatments | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|
| Control Group | Challenge Group | 0.5 mg/kg MccJ25 | 1.0 mg/kg MccJ25 | Antibiotic Group | |||
| Duodenum | |||||||
| Villus height (μm) | 754.82 a | 636.27 e | 686.04 c | 695.05 b | 672.18 d | 2.68 | <0.01 |
| Crypt depth (μm) | 101.21 a | 90.56 b | 90.2 b | 91.42 b | 90.12 b | 0.64 | <0.01 |
| Villus height/crypt depth | 7.46 a | 7.03 b | 7.61 a | 7.61 a | 7.46 a | 0.06 | <0.01 |
| Jejunum | |||||||
| Villus height (μm) | 322.27 bc | 303.57 d | 325.15 b | 331.47 a | 318.78 c | 1.92 | <0.01 |
| Crypt depth (μm) | 94.24 | 97.94 | 96.66 | 96.37 | 97.29 | 1.30 | 0.33 |
| Villus height/crypt depth | 3.42 ab | 3.11 c | 3.37 ab | 3.44 a | 3.28 b | 0.05 | <0.01 |
| Ileum | |||||||
| Villus height (μm) | 252.94 a | 242 b | 236.19 b | 242.56 b | 245.63 ab | 3.10 | <0.01 |
| Crypt depth (μm) | 63.07 b | 75.11 a | 60.72 b | 60.53 b | 60.85 b | 0.90 | <0.01 |
| Villus height/crypt depth | 4.02 a | 3.23 b | 3.9 a | 4.02 a | 4.04 a | 0.07 | <0.01 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Song, Q.; Huang, S.; Wang, Y.; Cai, S.; Yu, H.; Ding, X.; Zeng, X.; Zhang, J. Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella. Animals 2020, 10, 345. https://doi.org/10.3390/ani10020345
Wang G, Song Q, Huang S, Wang Y, Cai S, Yu H, Ding X, Zeng X, Zhang J. Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella. Animals. 2020; 10(2):345. https://doi.org/10.3390/ani10020345
Chicago/Turabian StyleWang, Gang, Qinglong Song, Shuo Huang, Yuming Wang, Shuang Cai, Haitao Yu, Xiuliang Ding, Xiangfang Zeng, and Jiang Zhang. 2020. "Effect of Antimicrobial Peptide Microcin J25 on Growth Performance, Immune Regulation, and Intestinal Microbiota in Broiler Chickens Challenged with Escherichia coli and Salmonella" Animals 10, no. 2: 345. https://doi.org/10.3390/ani10020345