
Assessment of the Degradation Potential and Genomic Insights towards Phenanthrene by Dietzia psychralcaliphila JI1D

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sediment Collection
2.2. Media and Chemicals
2.3. Isolation of Phenanthrene Degrading Bacteria
2.4. Molecular and Phylogenetic Analysis
2.5. E24 Assay
2.6. Biodegradation Analysis of Phenanthrene
2.6.1. Quantification Analysis by High-Performance Liquid Chromatography (HPLC)
2.6.2. Pathway Analysis by Gas Chromatography–Mass Spectrometry (GC-MS)
2.7. Whole-Genome Sequencing
2.7.1. Whole Genome Sequencing and De Novo Assembly
2.7.2. Genome Annotation
2.7.3. Phylogenetic Relationship
3. Results
3.1. Isolation, Molecular Identification, and Phylogenetic Relationship of Phenanthrene-Degrading Bacteria
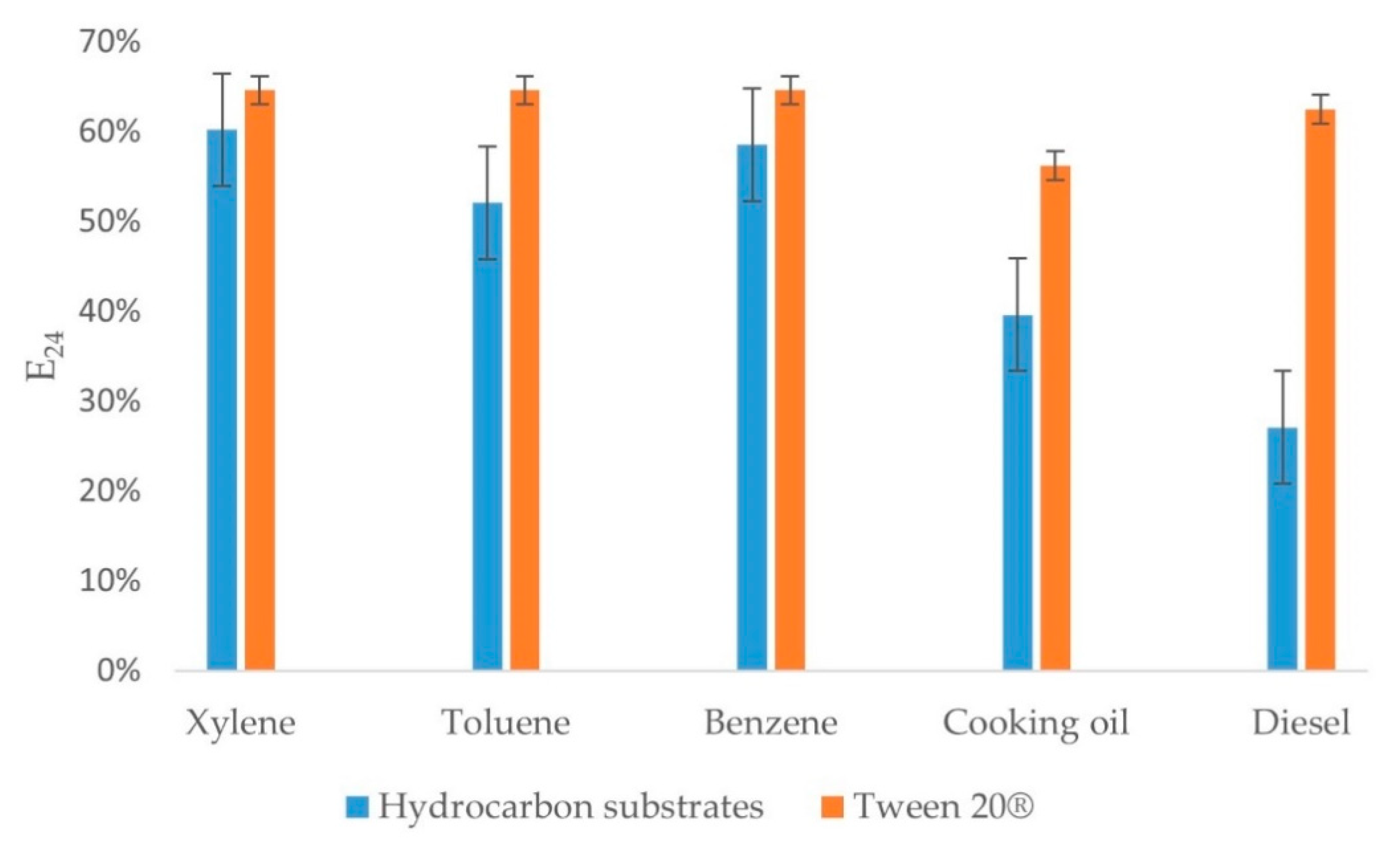
3.2. E24 Assay
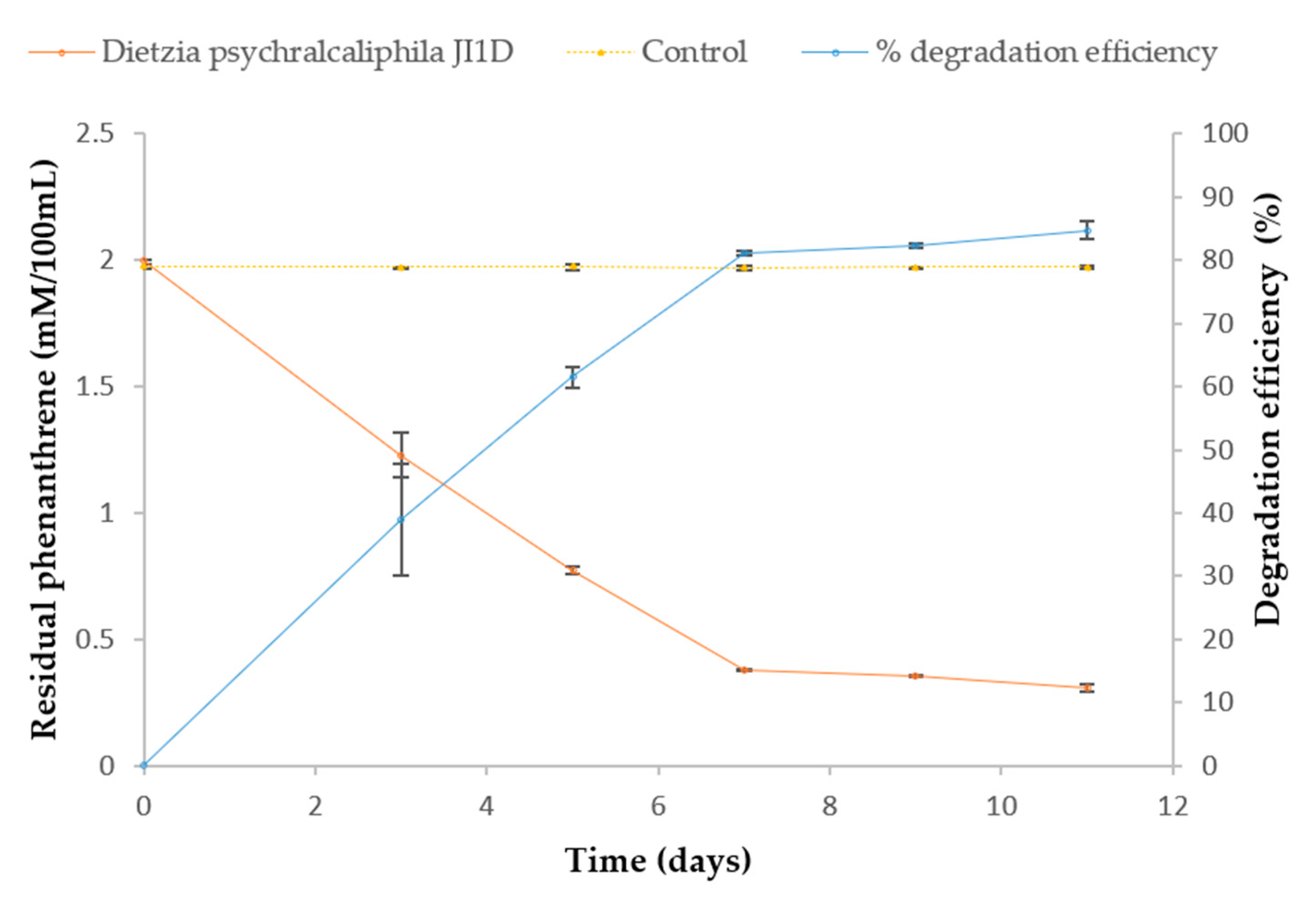
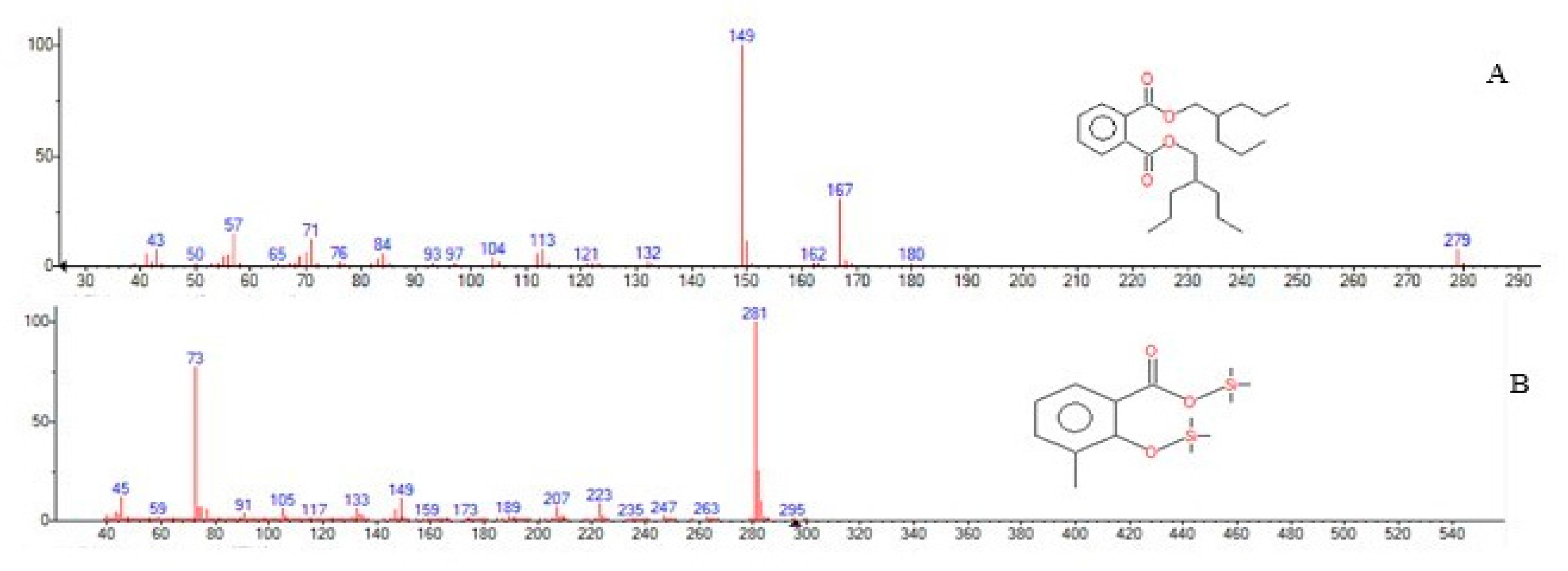
3.3. Degradation and Identification of Phenanthrene Degradation Products
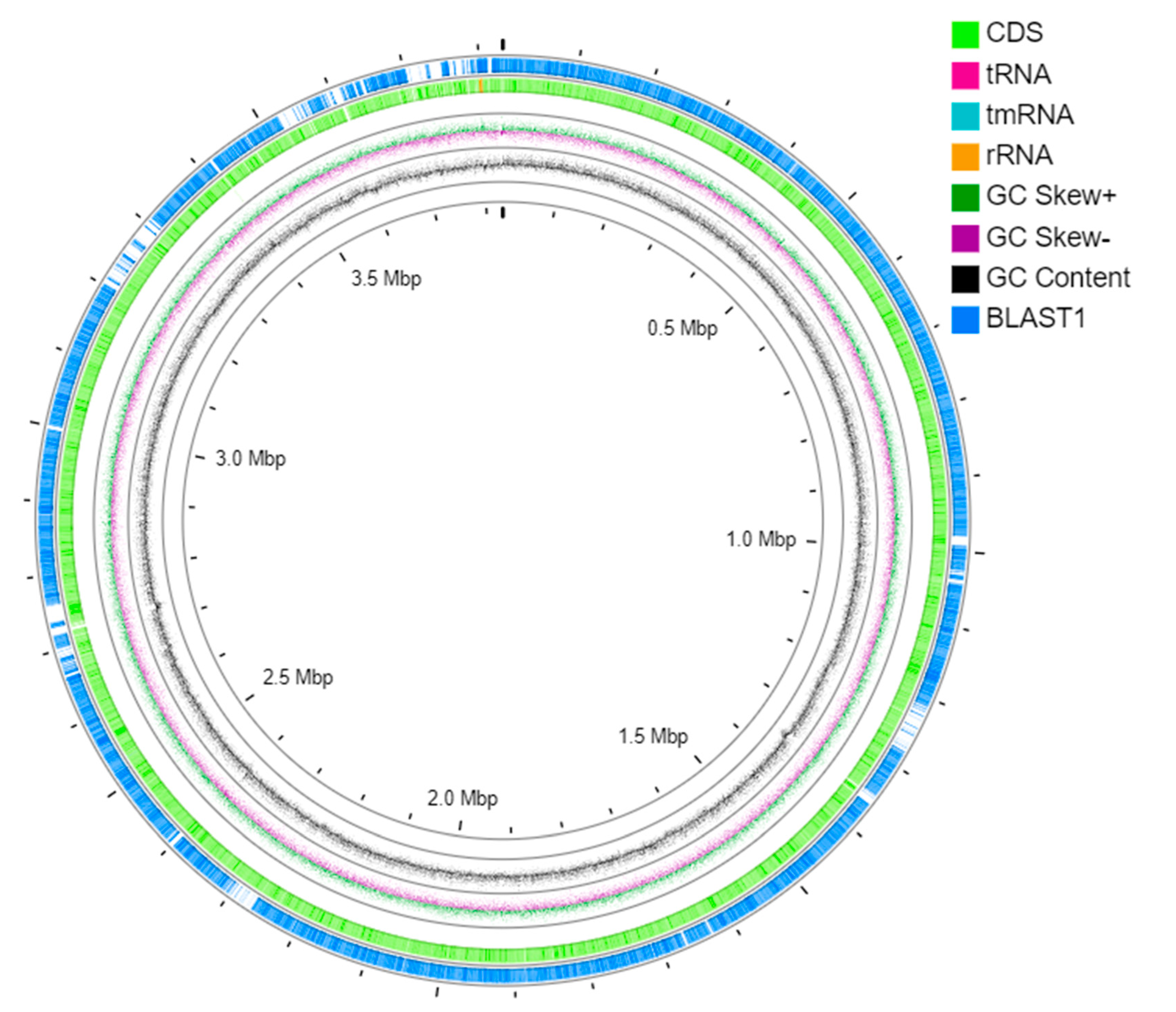
3.4. Whole Genome Sequencing and Genome Assembly
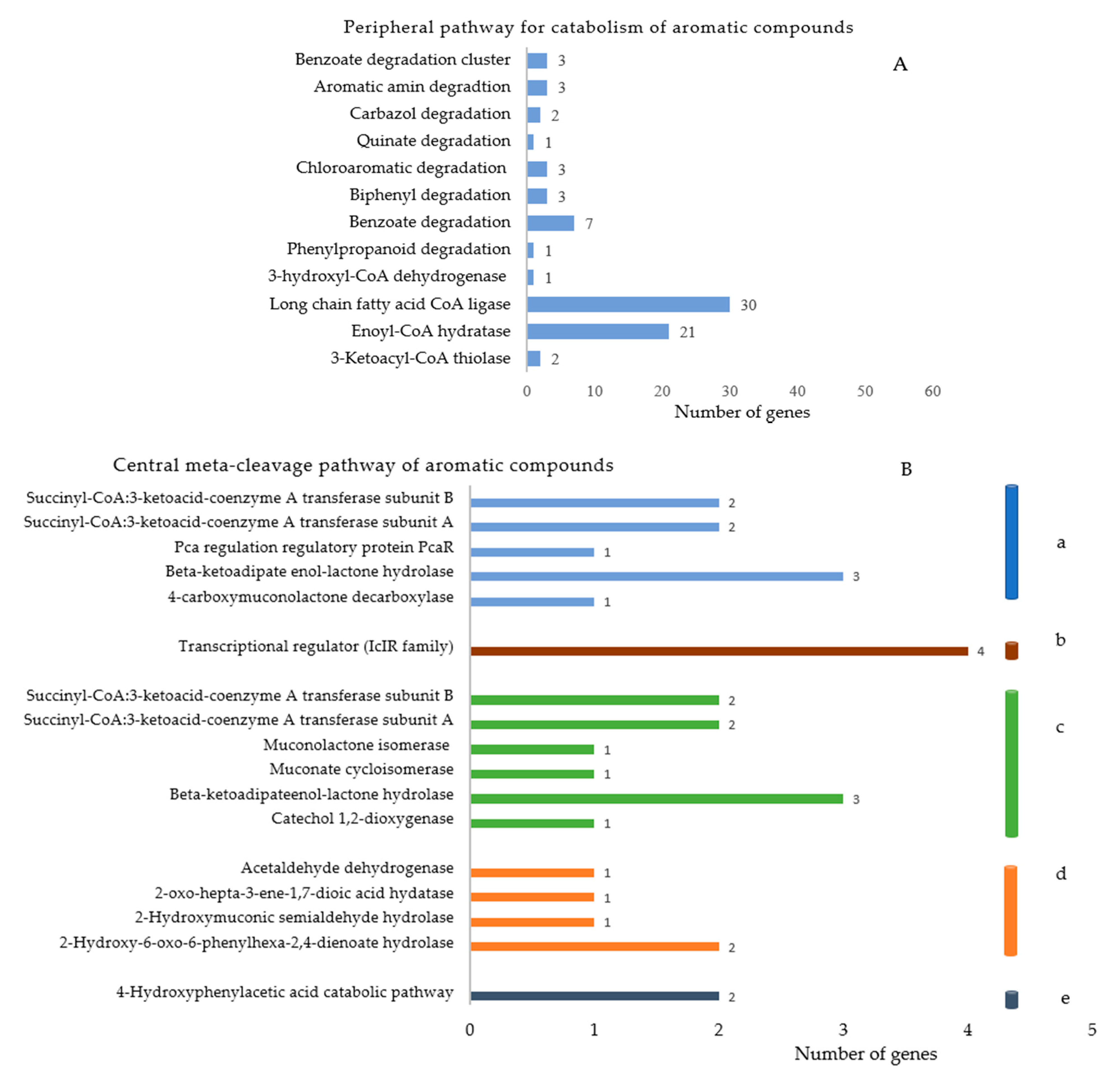
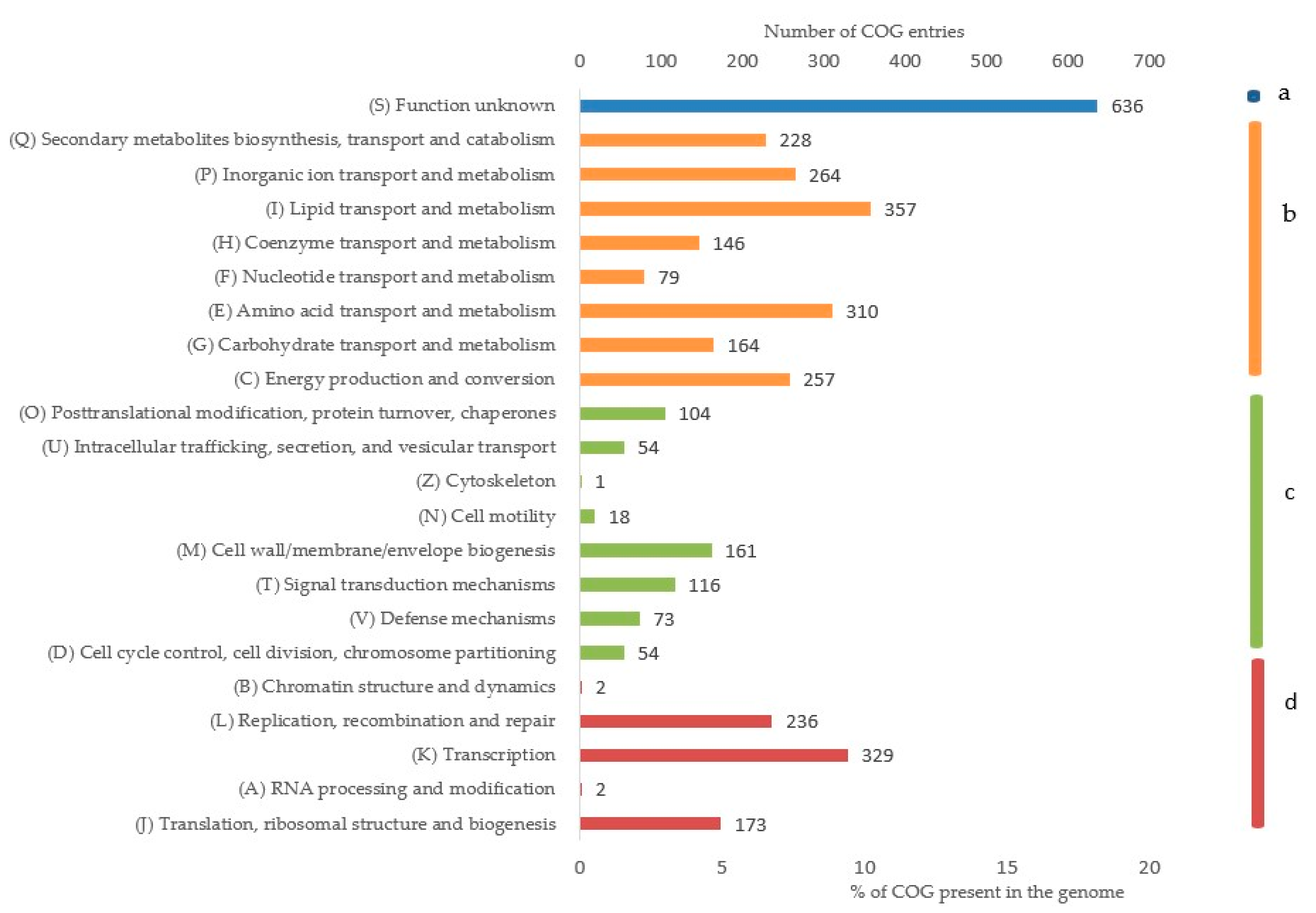
3.5. Genome Annotation
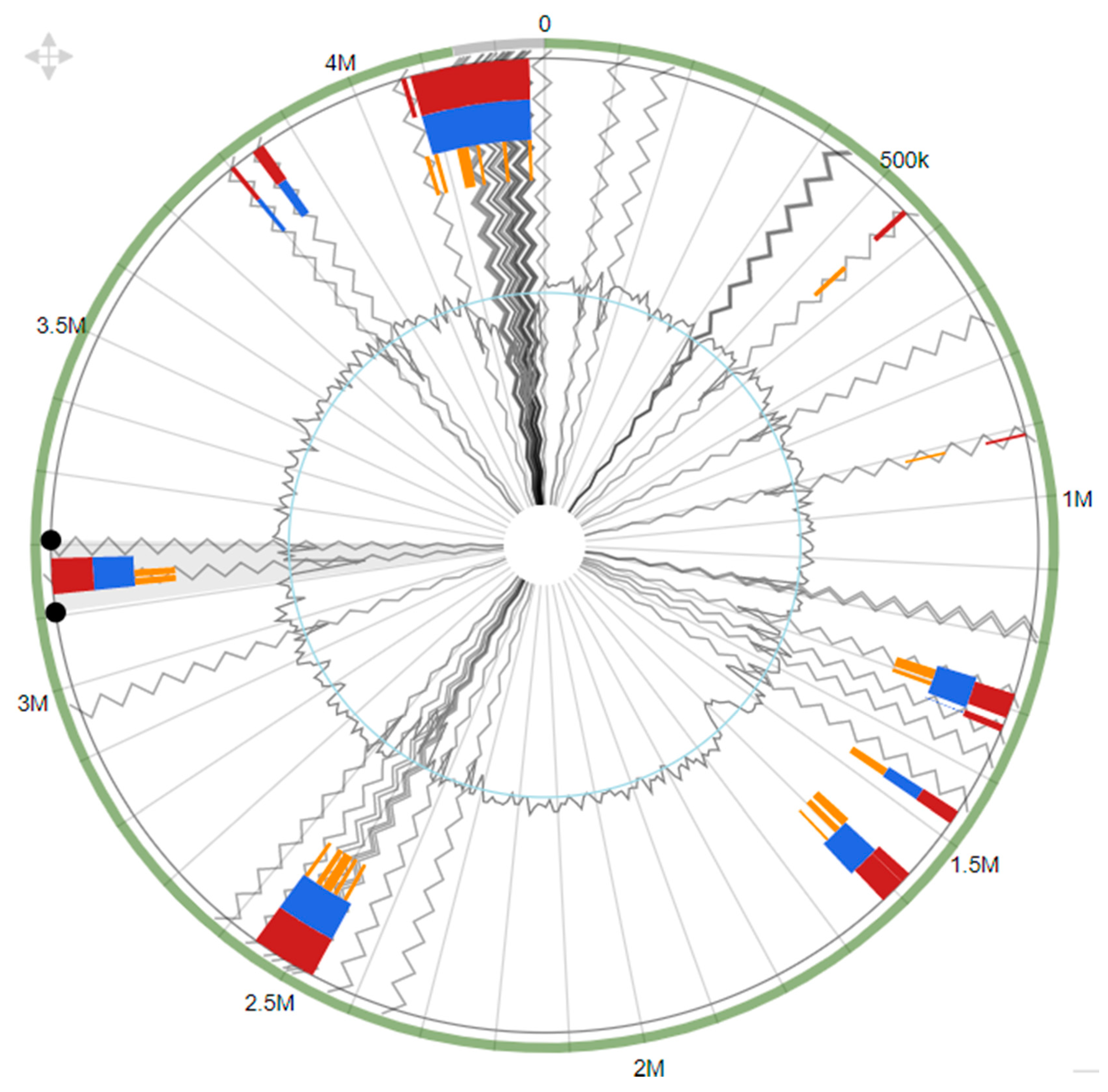
3.6. Genomic Islands
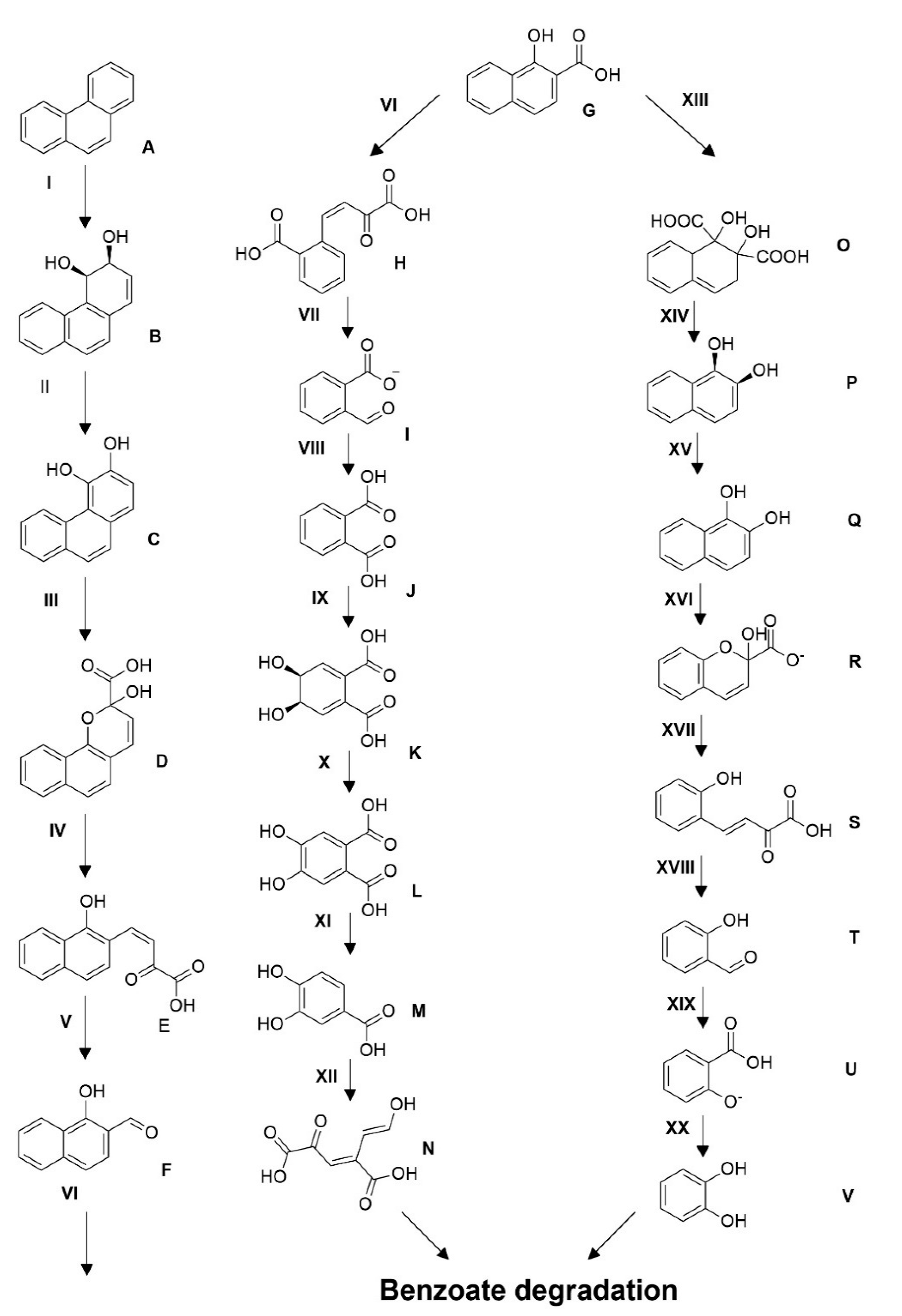
3.7. Proposed Pathway of Phenanthrene Degradation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lerda, D. Polycyclic Aromatic Hydrocarbons (PAHs) Factsheet; European Commission: Brussels, Belgium, 2011; p. 27. [Google Scholar]
- Mackay, D.; Callcott, D. Partitioning and Physical Chemical Properties of PAHs. In PAHs and Related Compounds; Springer: Berlin/Heidelberg, Germany, 1998; Volume 3, pp. 325–345. [Google Scholar] [CrossRef]
- Rathour, R.; Gupta, J.; Tyagi, B.; Kumari, T.; Thakur, I.S. Biodegradation of pyrene in soil microcosm by Shewanella sp. ISTPL2, a psychrophilic, alkalophilic and halophilic bacterium. Bioresour. Technol. Rep. 2018, 4, 129–136. [Google Scholar] [CrossRef]
- Gan, S.; Von Lau, E.; Ng, H.K. Remediation of soils contaminated with polycyclic aromatic hydrocarbons (PAHs). J. Hazard. Mater. 2009, 172, 532–549. [Google Scholar] [CrossRef] [PubMed]
- A Heitkamp, M.; Franklin, W.; E Cerniglia, C. Microbial metabolism of polycyclic aromatic hydrocarbons: Isolation and characterization of a pyrene-degrading bacterium. Appl. Environ. Microbiol. 1988, 54, 2549–2555. [Google Scholar] [CrossRef] [Green Version]
- Mallick, S.; Chakraborty, J.; Dutta, T.K. Role of oxygenases in guiding diverse metabolic pathways in the bacterial degradation of low-molecular-weight polycyclic aromatic hydrocarbons: A review. Crit. Rev. Microbiol. 2010, 37, 64–90. [Google Scholar] [CrossRef] [PubMed]
- Nogales, J.; García, J.L.; Díaz, E. Degradation of Aromatic Compounds in Pseudomonas: A Systems Biology View. In Aerobic Utilization of Hydrocarbons, Oils and Lipids; Springer: New York, NY, USA, 2017; pp. 1–49. [Google Scholar]
- Ghosal, D.; Ghosh, S.; Dutta, T.K.; Ahn, Y. Current State of Knowledge in Microbial Degradation of Polycyclic Aromatic Hydrocarbons (PAHs): A Review. Front. Microbiol. 2016, 7, 1369. [Google Scholar] [CrossRef] [Green Version]
- Evans, W.C.; Fernley, H.N.; Griffiths, E.; Davies, J.; Ribbons, D.W.; Hopper, D.J.; Chapman, P.J.; Dagley, S. Oxidative metabolism of phenanthrene and anthracene by soil pseudomonads. The ring-fission mechanism. Biochem. J. 1965, 95, 819–831. [Google Scholar] [CrossRef] [Green Version]
- Seo, J.-S.; Keum, Y.-S.; Hu, Y.; Lee, S.-E.; Li, Q.X. Degradation of phenanthrene by Burkholderia sp. C3: Initial 1,2- and 3,4-dioxygenation and meta- and ortho-cleavage of naphthalene-1,2-diol. Biodegradation 2007, 18, 123–131. [Google Scholar] [CrossRef]
- Seo, J.-S.; Keum, Y.-S.; Li, Q.X. Bacterial Degradation of Aromatic Compounds. Int. J. Environ. Res. Public Health 2009, 6, 278–309. [Google Scholar] [CrossRef]
- Elufisan, T.O.; Rodríguez-Luna, I.C.; Oyedara, O.O.; Sánchez-Varela, A.; Hernández-Mendoza, A.; Gonzalez, E.D.; Paz-González, A.D.; Muhammad, K.; Rivera, G.; Villalobos-Lopez, M.A.; et al. The Polycyclic Aromatic Hydrocarbon (PAH) degradation activities and genome analysis of a novel strain Stenotrophomonas sp. Pemsol isolated from Mexico. PeerJ 2020, 8, e8102. [Google Scholar] [CrossRef] [Green Version]
- Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Biodegradation of high-molecular weight PAHs by Rhodococcus wratislaviensis strain 9: Overexpression of amidohydrolase induced by pyrene and BaP. Sci. Total Environ. 2019, 651, 813–821. [Google Scholar] [CrossRef]
- Peng, T.; Luo, A.; Kan, J.; Liang, L.; Huang, T.; Hu, Z. Identification of A Ring-Hydroxylating Dioxygenases Capable of Anthracene and Benz[a]anthracene Oxidization from Rhodococcus sp. P14. J. Mol. Microbiol. Biotechnol. 2018, 28, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Kundu, A.; Das Banerjee, T.; Mohapatra, B.; Roy, A.; Manna, R.; Sar, P.; Kazy, S.K. Genome analysis of crude oil degrading Franconibacter pulveris strain DJ34 revealed its genetic basis for hydrocarbon degradation and survival in oil contaminated environment. Genomics 2017, 109, 374–382. [Google Scholar] [CrossRef]
- Kotoky, R.; Pandey, P. The genomic attributes of Cd-resistant, hydrocarbonoclastic Bacillus subtilis SR1 for rhizodegradation of benzo(a)pyrene under co-contaminated conditions. Genomics 2021, 113, 613–623. [Google Scholar] [CrossRef]
- Bødtker, G.; Hvidsten, I.V.; Barth, T.; Torsvik, T. Hydrocarbon degradation by Dietzia sp. A14101 isolated from an oil reservoir model column. Antonie Leeuwenhoek 2009, 96, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Bihari, Z.; Szabó, Z.; Szvetnik, A.; Balázs, M.; Bartos, P.; Tolmacsov, P.; Zombori, Z.; Kiss, I. Characterization of a Novel Long-Chain n-Alkane-Degrading Strain, Dietzia sp. E1. Z. Nat. C 2010, 65, 693–700. [Google Scholar] [CrossRef]
- Chen, W.; Li, J.; Sun, X.; Min, J.; Hu, X. High efficiency degradation of alkanes and crude oil by a salt-tolerant bacterium Dietzia species CN-3. Int. Biodeterior. Biodegrad. 2017, 118, 110–118. [Google Scholar] [CrossRef]
- Al-Awadhi, H.; Sulaiman, R.H.D.; Mahmoud, H.M.; Radwan, S.S. Alkaliphilic and halophilic hydrocarbon-utilizing bacteria from Kuwaiti coasts of the Arabian Gulf. Appl. Microbiol. Biotechnol. 2007, 77, 183–186. [Google Scholar] [CrossRef]
- Gurav, R.; Lyu, H.; Ma, J.; Tang, J.; Liu, Q.; Zhang, H. Degradation of n-alkanes and PAHs from the heavy crude oil using salt-tolerant bacterial consortia and analysis of their catabolic genes. Environ. Sci. Pollut. Res. 2017, 24, 11392–11403. [Google Scholar] [CrossRef] [PubMed]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Mishra, A.; Rathour, R.; Singh, R.; Kumari, T.; Thakur, I.S. Degradation and detoxification of phenanthrene by actinobacterium Zhihengliuella sp. ISTPL4. Environ. Sci. Pollut. Res. 2020, 27, 27256–27267. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Meslier, V.; Loux, V.; Renault, P. Genome Sequence of Leuconostoc pseudomesenteroides Strain 4882, Isolated from a Dairy Starter Culture. J. Bacteriol. 2012, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; von Mering, C.; Bork, P. Fast Genome-Wide Functional Annotation through Orthology Assignment by eggNOG-Mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. eggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelli, C.; Laird, M.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. Simon Fraser University Research Computing Group IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef] [PubMed]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35, W182–W185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, C.; Rodriguez-R, L.M.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Gran-Scheuch, A.; Fuentes, E.; Bravo, D.M.; Jiménez, J.C.; Pérez-Donoso, J.M. Isolation and Characterization of Phenanthrene Degrading Bacteria from Diesel Fuel-Contaminated Antarctic Soils. Front. Microbiol. 2017, 8, 1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haleyur, N.; Shahsavari, E.; Taha, M.; Khudur, L.S.; Koshlaf, E.; Osborn, A.M.; Ball, A.S. Assessing the degradation efficacy of native PAH-degrading bacteria from aged, weathered soils in an Australian former gasworks site. Geoderma 2018, 321, 110–117. [Google Scholar] [CrossRef]
- Yumoto, I.; Nakamura, A.; Iwata, H.; Kojima, K.; Kusumoto, K.; Nodasaka, Y.; Matsuyama, H. Dietzia psychralcaliphila sp. nov., a novel, facultatively psychrophilic alkaliphile that grows on hydrocarbons. Int. J. Syst. Evol. Microbiol. 2002, 52, 85–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Yu, M.; Chen, J. Draft genome analysis of Dietzia sp. 111N12-1, isolated from the South China Sea with bioremediation activity. Braz. J. Microbiol. 2017, 48, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Duckworth, A.W.; Grant, S.; Grant, W.D.; Jones, B.E.; Meijer, D. Dietzia natronolimnaios sp. nov., a new member of the genus Dietzia isolated from an East African soda lake. Extremophiles 1998, 2, 359–366. [Google Scholar] [CrossRef]
- Rainey, F.A.; Klatte, S.; Kroppenstedt, R.M.; Stackebrandt, E. Dietzia, New Genus Including Dietzia maris comb. nov., Formerly Rhodococcus maris. Int. J. Syst. Bacteriol. 1995, 45, 32–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezza, F.A.; Beukes, M.; Chirwa, E.M.N. Application of biosurfactant produced by Ochrobactrum intermedium CN3 for enhancing petroleum sludge bioremediation. Process. Biochem. 2015, 50, 1911–1922. [Google Scholar] [CrossRef]
- Kavyanifard, A.; Ebrahimipour, G.; Ghasempour, A. Structure characterization of a methylated ester biosurfactant produced by a newly isolated Dietzia cinnamea KA1. Microbiology 2016, 85, 430–435. [Google Scholar] [CrossRef]
- Ewang, W.; Ecai, B.; Eshao, Z. Oil degradation and biosurfactant production by the deep sea bacterium Dietzia maris As-13-3. Front. Microbiol. 2014, 5, 711. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Luan, T.; Lin, L.; Liu, H.; Tam, N.F. Production of metabolites in the biodegradation of phenanthrene, fluoranthene and pyrene by the mixed culture of Mycobacterium sp. and Sphingomonas sp. Bioresour. Technol. 2011, 102, 2965–2972. [Google Scholar] [CrossRef]
- Fathi, Z.; Ebrahimipour, G.; Kavyanifard, A.; Najmi, Z. Optimization of Phenanthrene-degradation by Dietzia cinnamea AP for Bioremediation Applications. Res. Mol. Med. 2016, 4, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum Hydrocarbon-Degrading Bacteria for the Remediation of Oil Pollution Under Aerobic Conditions: A Perspective Analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef]
- Mahto, K.U.; Das, S. Whole genome characterization and phenanthrene catabolic pathway of a biofilm forming marine bacterium Pseudomonas aeruginosa PFL-P1. Ecotoxicol. Environ. Saf. 2020, 206, 111087. [Google Scholar] [CrossRef]
- Shukla, S.K.; Mangwani, N.; Rao, T.; Das, S. Biofilm-Mediated Bioremediation of Polycyclic Aromatic Hydrocarbons. In Microbial Biodegradation and Bioremediation; Elsevier BV: Amsterdam, The Netherlands, 2014; pp. 203–232. [Google Scholar]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; I Hurwitz, D.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser-Liggett, C.M. Insights on biology and evolution from microbial genome sequencing. Genome Res. 2005, 15, 1603–1610. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, L.; Shao, Z. Pseudomonas, the dominant polycyclic aromatic hydrocarbon-degrading bacteria isolated from Antarctic soils and the role of large plasmids in horizontal gene transfer. Environ. Microbiol. 2006, 8, 455–465. [Google Scholar] [CrossRef]
- Tropel, D.; van der Meer, J.R. Bacterial Transcriptional Regulators for Degradation Pathways of Aromatic Compounds. Microbiol. Mol. Biol. Rev. 2004, 68, 474–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binnewies, T.T.; Motro, Y.; Hallin, P.F.; Lund, O.; Dunn, D.; La, T.; Hampson, D.; Bellgard, M.; Wassenaar, T.; Ussery, D.W. Ten years of bacterial genome sequencing: Comparative-genomics-based discoveries. Funct. Integr. Genom. 2006, 6, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Eulberg, D.; Schlömann, M. The putative regulator of catechol catabolism in Rhodococcus opacus 1CP—An IclR-type, not a LysR-type transcriptional regulator. Antonie Leeuwenhoek 1998, 74, 71–82. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Statistics |
|---|---|
| Total genome size including gaps (bp) | 4,216,480 |
| Number of contigs | 52 |
| Ratio of bases that have phred quality score of over 20 (%) | 96.08 |
| Ratio of bases that have phred quality score of over 30 (%) | 88.96 |
| Contig N50 (bp) | 270,109 |
| Average contig length (bp) | 81,086 |
| Longest contig length (bp) | 601,190 |
| Shortest contig length (bp) | 1026 |
| GC (%) | 69.09 |
| Content | Statistics |
|---|---|
| Total number of genes predicted | 3961 |
| Number of genes assigned with CDSs | 3902 |
| Number of genes assigned to UniprotKB | 1899 |
| Number of genomic islands assigned | 29 |
| tRNA | 55 |
| rRNA | 4 (2 × 5S rRNA,1 × 23S rRNA,1 × 16S rRNA) |
| ncRNAs | 21 |
| Number of genes with non-hypothetical function | 2514 |
| Number of genes with EC-number | 1611 |
| Number of genes with Seed Subsystem Ontology | 1245 |
| Average protein length (aa) | 325 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ausuri, J.; Vitale, G.A.; Coppola, D.; Palma Esposito, F.; Buonocore, C.; de Pascale, D. Assessment of the Degradation Potential and Genomic Insights towards Phenanthrene by Dietzia psychralcaliphila JI1D. Microorganisms 2021, 9, 1327. https://doi.org/10.3390/microorganisms9061327
Ausuri J, Vitale GA, Coppola D, Palma Esposito F, Buonocore C, de Pascale D. Assessment of the Degradation Potential and Genomic Insights towards Phenanthrene by Dietzia psychralcaliphila JI1D. Microorganisms. 2021; 9(6):1327. https://doi.org/10.3390/microorganisms9061327
Chicago/Turabian StyleAusuri, Janardhan, Giovanni Andrea Vitale, Daniela Coppola, Fortunato Palma Esposito, Carmine Buonocore, and Donatella de Pascale. 2021. "Assessment of the Degradation Potential and Genomic Insights towards Phenanthrene by Dietzia psychralcaliphila JI1D" Microorganisms 9, no. 6: 1327. https://doi.org/10.3390/microorganisms9061327